
A Study of the Metabolic Pathways Affected by Gestational Diabetes Mellitus: Comparison with Type 2 Diabetes
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Samples
2.3. Determination of Biochemical Parameters
2.4. 1H NMR Spectroscopy of Macromolecules-Free Serum
2.5. Targeted Metabolite Profiling
2.6. Statistical Analysis
2.7. Untargeted Metabolite Profiling
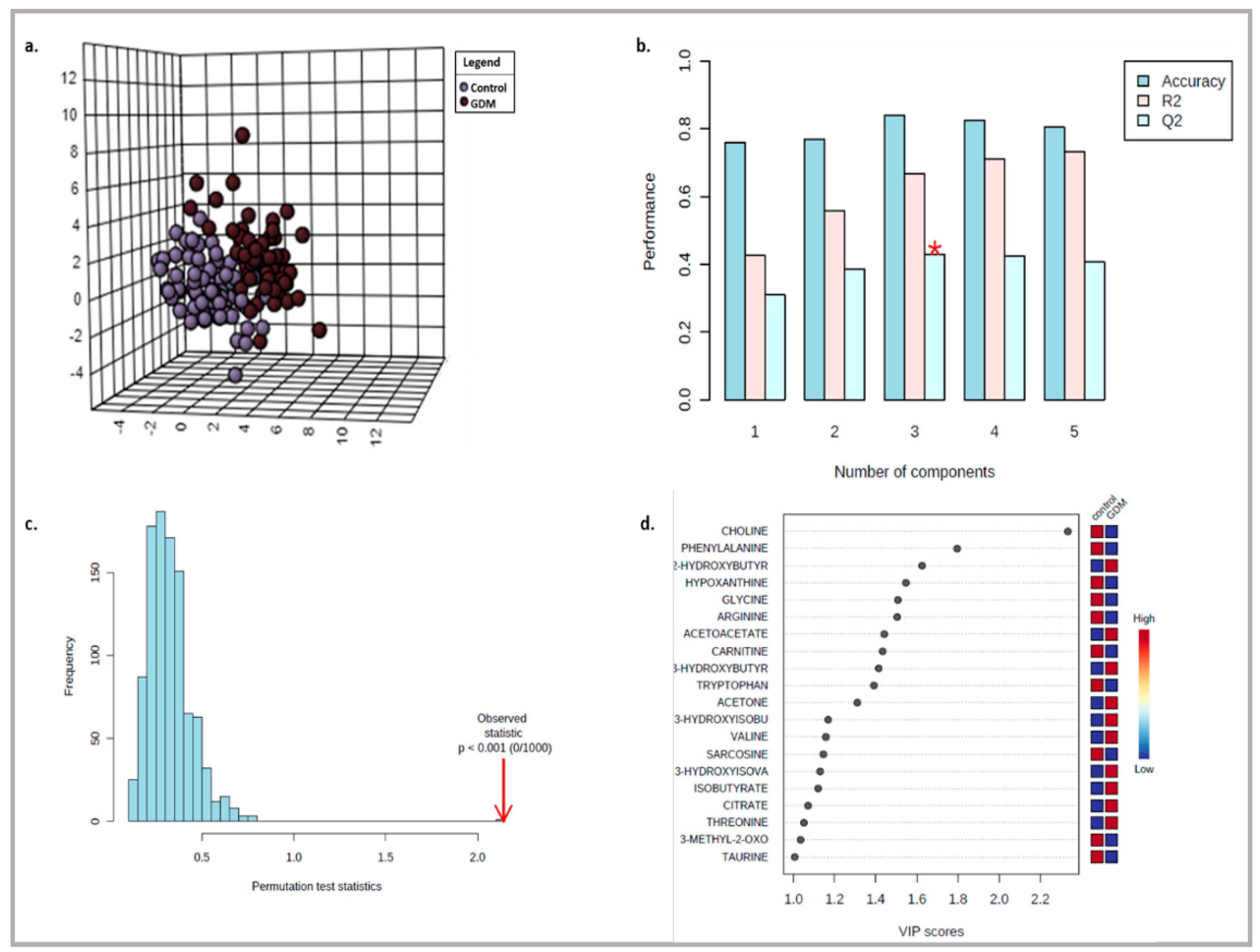
3. Results
3.1. Targeted Metabolite Profiling
3.2. Untargeted Metabolite Profiling
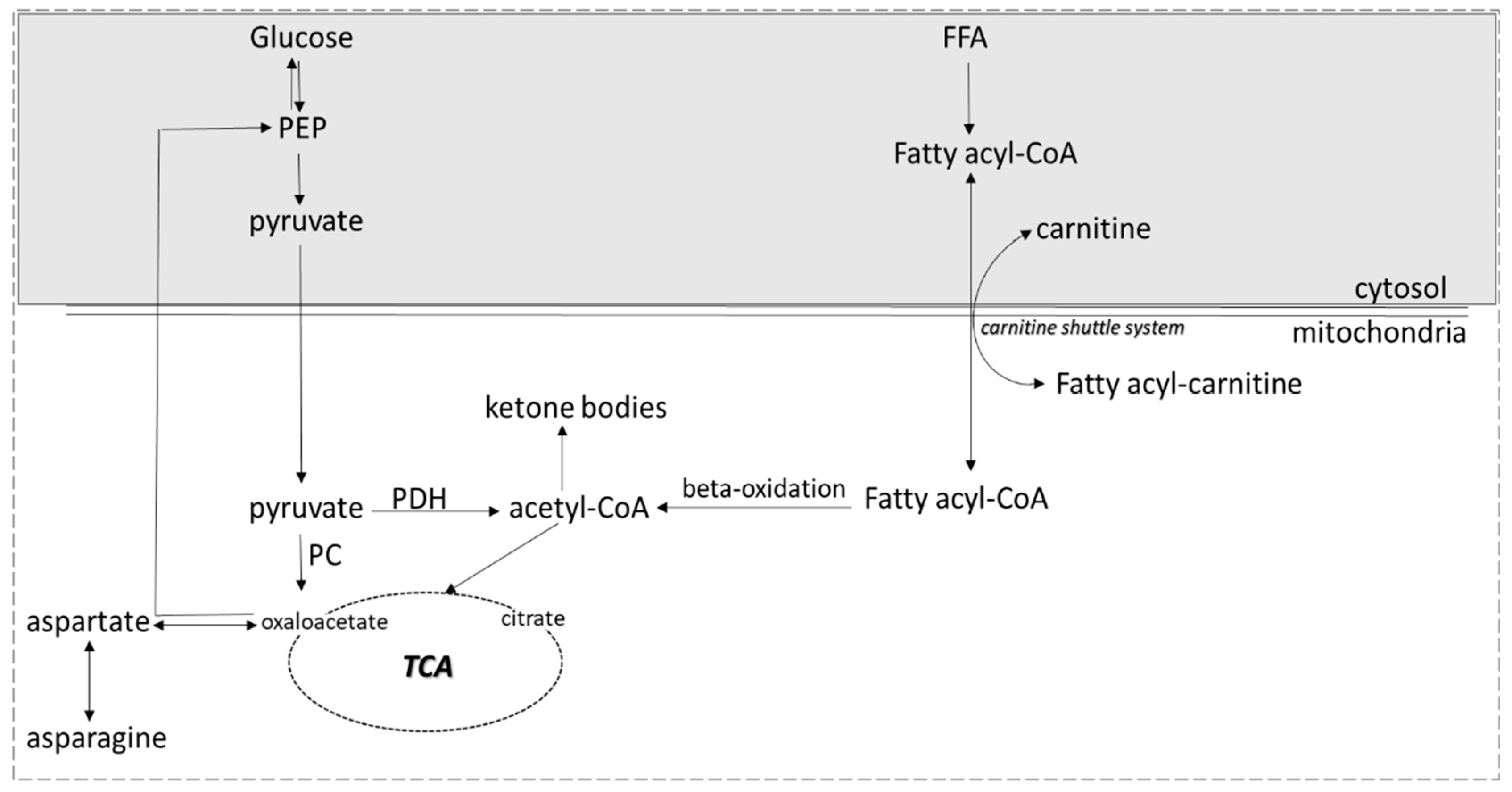
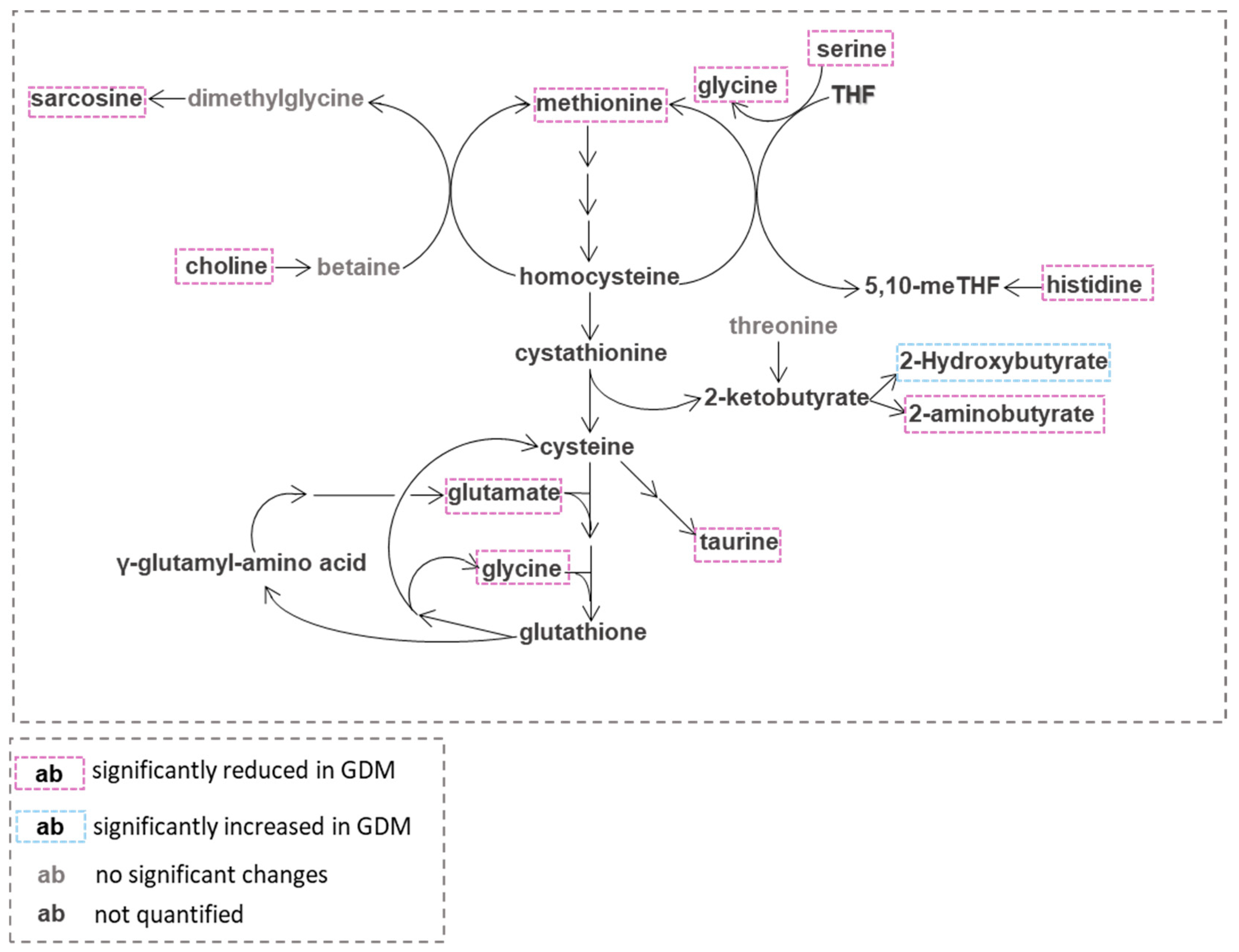
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American Diabetes Association. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes—2019. Diabetes Care 2018, 42, S13–S28. [Google Scholar] [CrossRef] [Green Version]
- International Association of Diabetes and Pregnancy Study Groups Consensus Panel; Metzger, B.E.; Gabbe, S.G.; Persson, B.; Buchanan, T.A.; Catalano, P.A.; Damm, P.; Dyer, A.R.; Leiva, A.; Hod, M.; et al. International association of diabetes and pregnancy study groups recommendations on the diagnosis and classification of hyperglycemia in pregnancy. Diabetes Care 2010, 33, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, L.P.; Metzger, B.E.; Dyer, A.R.; Lowe, J.; McCance, D.R.; Lappin, T.R.; Trimble, E.R.; Coustan, D.R.; Hadden, D.R.; Hod, M.; et al. Hyperglycemia and Adverse Pregnancy Outcome (HAPO) Study: Associations of maternal A1C and glucose with pregnancy outcomes. Diabetes Care 2012, 35, 574–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchanan, T.A.; Xiang, A.H. Gestational diabetes mellitus. J. Clin. Investig. 2005, 115, 485–491. [Google Scholar] [CrossRef] [Green Version]
- Dalfra, M.G.; Burlina, S.; Del Vescovo, G.G.; Lapolla, A. Genetics and Epigenetics: New Insight on Gestational Diabetes Mellitus. Front. Endocrinol. 2020, 11, 602477. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Holmes, E.; Kinross, J.M.; Darzi, A.W.; Takats, Z.; Lindon, J.C. Metabolic phenotyping in clinical and surgical environments. Nature 2012, 491, 384–392. [Google Scholar] [CrossRef]
- Azad, R.K.; Shulaev, V. Metabolomics technology and bioinformatics for precision medicine. Brief. Bioinform. 2019, 20, 1957–1971. [Google Scholar] [CrossRef]
- Nagana Gowda, G.A.; Raftery, D. Quantitating metabolites in protein precipitated serum using NMR spectroscopy. Anal. Chem. 2014, 86, 5433–5440. [Google Scholar] [CrossRef]
- Dudzik, D.; Zorawski, M.; Skotnicki, M.; Zarzycki, W.; Kozlowska, G.; Bibik-Malinowska, K.; Vallejo, M.; Garcia, A.; Barbas, C.; Ramos, M.P. Metabolic fingerprint of Gestational Diabetes Mellitus. J. Proteom. 2014, 103, 57–71. [Google Scholar] [CrossRef]
- Huo, X.; Li, J.; Cao, Y.F.; Li, S.N.; Shao, P.; Leng, J.; Li, W.; Liu, J.; Yang, K.; Ma, R.C.W.; et al. Trimethylamine N-Oxide Metabolites in Early Pregnancy and Risk of Gestational Diabetes: A Nested Case-Control Study. J. Clin. Endocrinol. Metab. 2019, 104, 5529–5539. [Google Scholar] [CrossRef]
- Flanagan, J.L.; Simmons, P.A.; Vehige, J.; Willcox, M.D.; Garrett, Q. Role of carnitine in disease. Nutr. Metab. 2010, 7, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoppel, C. The role of carnitine in normal and altered fatty acid metabolism. Am. J. Kidney Dis. 2003, 41, S4–S12. [Google Scholar] [CrossRef]
- Lopaschuk, G. Regulation of carbohydrate metabolism in ischemia and reperfusion. Am. Heart J. 2000, 139, S115–S119. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Liu, Y.; Zhang, Y.; Long, M.; Guo, Y.; Wang, Z.; Li, X.; Zhang, C.; Li, X.; He, J.; et al. Effect of non-esterified fatty acids on fatty acid metabolism-related genes in calf hepatocytes cultured in vitro. Cell Physiol. Biochem. 2013, 32, 1509–1516. [Google Scholar] [CrossRef] [PubMed]
- Ringseis, R.; Hanisch, N.; Seliger, G.; Eder, K. Low availability of carnitine precursors as a possible reason for the diminished plasma carnitine concentrations in pregnant women. BMC Pregnancy Childbirth 2010, 10, 17. [Google Scholar] [CrossRef] [Green Version]
- Talian, G.C.; Komlosi, K.; Decsi, T.; Koletzko, B.; Melegh, B. Determination of carnitine ester patterns during the second half of pregnancy, at delivery, and in neonatal cord blood by tandem mass spectrometry: Complex and dynamic involvement of carnitine in the intermediary metabolism. Pediatr. Res. 2007, 62, 88–92. [Google Scholar] [CrossRef] [Green Version]
- Arenas, J.; Rubio, J.C.; Martin, M.A.; Campos, Y. Biological roles of L-carnitine in perinatal metabolism. Early Hum. Dev. 1998, 53 (Suppl. 1), S43–S50. [Google Scholar] [CrossRef]
- Grube, M.; Meyer Zu Schwabedissen, H.; Draber, K.; Prager, D.; Moritz, K.U.; Linnemann, K.; Fusch, C.; Jedlitschky, G.; Kroemer, H.K. Expression, localization, and function of the carnitine transporter octn2 (slc22a5) in human placenta. Drug Metab. Dispos. 2005, 33, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Herrera, E.; Ortega-Senovilla, H. Maternal lipid metabolism during normal pregnancy and its implications to fetal development. Clin. Lipidol. 2010, 5, 899–911. [Google Scholar] [CrossRef] [Green Version]
- Catalano, P.M.; Nizielski, S.E.; Shao, J.; Preston, L.; Qiao, L.; Friedman, J.E. Downregulated IRS-1 and PPARgamma in obese women with gestational diabetes: Relationship to FFA during pregnancy. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E522–E533. [Google Scholar] [CrossRef]
- Villafan-Bernal, J.R.; Acevedo-Alba, M.; Reyes-Pavon, R.; Diaz-Parra, G.A.; Lip-Sosa, D.L.; Vazquez-Delfin, H.I.; Hernandez-Munoz, M.; Bravo-Aguirre, D.E.; Figueras, F.; Martinez-Portilla, R.J. Plasma Levels of Free Fatty Acids in Women with Gestational Diabetes and Its Intrinsic and Extrinsic Determinants: Systematic Review and Meta-Analysis. J. Diabetes Res. 2019, 2019, 7098470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mynatt, R.L. Carnitine and type 2 diabetes. Diabetes Metab. Res. Rev. 2009, 25 (Suppl. S1), S45–S49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ringseis, R.; Keller, J.; Eder, K. Role of carnitine in the regulation of glucose homeostasis and insulin sensitivity: Evidence from in vivo and in vitro studies with carnitine supplementation and carnitine deficiency. Eur. J. Nutr. 2012, 51, 1–18. [Google Scholar] [CrossRef]
- Butte, N.F. Carbohydrate and lipid metabolism in pregnancy: Normal compared with gestational diabetes mellitus. Am. J. Clin. Nutr. 2000, 71, 1256S–1261S. [Google Scholar] [CrossRef] [Green Version]
- Dudzik, D.; Zorawski, M.; Skotnicki, M.; Zarzycki, W.; Garcia, A.; Angulo, S.; Lorenzo, M.P.; Barbas, C.; Ramos, M.P. GC-MS based Gestational Diabetes Mellitus longitudinal study: Identification of 2-and 3-hydroxybutyrate as potential prognostic biomarkers. J. Pharm. Biomed. Anal. 2017, 144, 90–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montelongo, A.; Lasuncion, M.A.; Pallardo, L.F.; Herrera, E. Longitudinal study of plasma lipoproteins and hormones during pregnancy in normal and diabetic women. Diabetes 1992, 41, 1651–1659. [Google Scholar] [CrossRef]
- Pappa, K.I.; Vlachos, G.; Theodora, M.; Roubelaki, M.; Angelidou, K.; Antsaklis, A. Intermediate metabolism in association with the amino acid profile during the third trimester of normal pregnancy and diet-controlled gestational diabetes. Am. J. Obstet. Gynecol. 2007, 196, 65.e1–65.e5. [Google Scholar] [CrossRef]
- Scholtens, D.M.; Muehlbauer, M.J.; Daya, N.R.; Stevens, R.D.; Dyer, A.R.; Lowe, L.P.; Metzger, B.E.; Newgard, C.B.; Bain, J.R.; Lowe, W.L., Jr.; et al. Metabolomics reveals broad-scale metabolic perturbations in hyperglycemic mothers during pregnancy. Diabetes Care 2014, 37, 158–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahendran, Y.; Vangipurapu, J.; Cederberg, H.; Stancakova, A.; Pihlajamaki, J.; Soininen, P.; Kangas, A.J.; Paananen, J.; Civelek, M.; Saleem, N.K.; et al. Association of ketone body levels with hyperglycemia and type 2 diabetes in 9398 Finnish men. Diabetes 2013, 62, 3618–3626. [Google Scholar] [CrossRef] [Green Version]
- Ottosson, F.; Smith, E.; Melander, O.; Fernandez, C. Altered Asparagine and Glutamate Homeostasis Precede Coronary Artery Disease and Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2018, 103, 3060–3069. [Google Scholar] [CrossRef]
- Vangipurapu, J.; Stancakova, A.; Smith, U.; Kuusisto, J.; Laakso, M. Nine Amino Acids Are Associated with Decreased Insulin Secretion and Elevated Glucose Levels in a 7.4-Year Follow-up Study of 5,181 Finnish Men. Diabetes 2019, 68, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Sinclair, K.D. One-carbon metabolism and epigenetic regulation of embryo development. Reprod. Fertil. Dev. 2015, 27, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Ducker, G.S.; Rabinowitz, J.D. One-Carbon Metabolism in Health and Disease. Cell Metab. 2017, 25, 27–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solans, A.; Estivill, X.; De la Luna, S. Cloning and characterization of human FTCD on 21q22.3, a candidate gene for glutamate formiminotransferase deficiency. Cytogenet. Cell Genet. 2000, 88, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, G.; Motokawa, Y.; Yoshida, T.; Hiraga, K. Glycine cleavage system: Reaction mechanism, physiological significance, and hyperglycinemia. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2008, 84, 246–263. [Google Scholar] [CrossRef] [Green Version]
- Finkelstein, J.D. Methionine metabolism in mammals. J. Nutr. Biochem. 1990, 1, 228–237. [Google Scholar] [CrossRef]
- Su, X.; Wellen, K.E.; Rabinowitz, J.D. Metabolic control of methylation and acetylation. Curr. Opin. Chem. Biol. 2016, 30, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Mentch, S.J.; Mehrmohamadi, M.; Huang, L.; Liu, X.; Gupta, D.; Mattocks, D.; Gomez Padilla, P.; Ables, G.; Bamman, M.M.; Thalacker-Mercer, A.E.; et al. Histone Methylation Dynamics and Gene Regulation Occur through the Sensing of One-Carbon Metabolism. Cell. Metab. 2015, 22, 861–873. [Google Scholar] [CrossRef] [Green Version]
- Kalhan, S.C. One-carbon metabolism, fetal growth and long-term consequences. Nestle Nutr. Inst. Workshop Ser. 2013, 74, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Kalhan, S.C. One carbon metabolism in pregnancy: Impact on maternal, fetal and neonatal health. Mol. Cell Endocrinol. 2016, 435, 48–60. [Google Scholar] [CrossRef]
- Knight, A.K.; Park, H.J.; Hausman, D.B.; Fleming, J.M.; Bland, V.L.; Rosa, G.; Kennedy, E.M.; Caudill, M.A.; Malysheva, O.; Kauwell, G.P.A.; et al. Association between one-carbon metabolism indices and DNA methylation status in maternal and cord blood. Sci. Rep. 2018, 8, 16873. [Google Scholar] [CrossRef] [PubMed]
- Shere, M.; Nguyen, P.; Tam, C.; Stern, S.; Kapur, B.; O’Connor, D.L.; Koren, G. Pregnancy-induced changes in the long-term pharmacokinetics of 1.1 mg vs. 5 mg folic acid: A randomized clinical trial. J. Clin. Pharmacol. 2015, 55, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Finer, S.; Saravanan, P.; Hitman, G.; Yajnik, C. The role of the one-carbon cycle in the developmental origins of Type 2 diabetes and obesity. Diabet. Med. 2014, 31, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.R.; Tyagi, S.C. Impaired Folate-Mediated One-Carbon Metabolism in Type 2 Diabetes, Late-Onset Alzheimer’s Disease and Long COVID. Medicina 2021, 58, 16. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Emiliano, J.; Fajardo-Araujo, M.E.; Zuniga-Trujillo, I.; Perez-Vazquez, V.; Sandoval-Salazar, C.; Ornelas-Vazquez, J.K. Mitochondrial content, oxidative, and nitrosative stress in human full-term placentas with gestational diabetes mellitus. Reprod. Biol. Endocrinol. 2017, 15, 26. [Google Scholar] [CrossRef] [Green Version]
- Alves, A.; Bassot, A.; Bulteau, A.L.; Pirola, L.; Morio, B. Glycine Metabolism and Its Alterations in Obesity and Metabolic Diseases. Nutrients 2019, 11, 1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, X.R.; Ong, S.E.; Goldberger, O.; Peng, J.; Sharma, R.; Thompson, D.A.; Vafai, S.B.; Cox, A.G.; Marutani, E.; Ichinose, F.; et al. Mitochondrial dysfunction remodels one-carbon metabolism in human cells. eLife 2016, 5, e35977. [Google Scholar] [CrossRef]
- Gall, W.E.; Beebe, K.; Lawton, K.A.; Adam, K.P.; Mitchell, M.W.; Nakhle, P.J.; Ryals, J.A.; Milburn, M.V.; Nannipieri, M.; Camastra, S.; et al. alpha-hydroxybutyrate is an early biomarker of insulin resistance and glucose intolerance in a nondiabetic population. PLoS ONE 2010, 5, e10883. [Google Scholar] [CrossRef] [Green Version]
- Irino, Y.; Toh, R.; Nagao, M.; Mori, T.; Honjo, T.; Shinohara, M.; Tsuda, S.; Nakajima, H.; Satomi-Kobayashi, S.; Shinke, T.; et al. 2-Aminobutyric acid modulates glutathione homeostasis in the myocardium. Sci. Rep. 2016, 6, 36749. [Google Scholar] [CrossRef] [Green Version]
- Bletsa, E.; Filippas-Dekouan, S.; Kostara, C.; Dafopoulos, P.; Dimou, A.; Pappa, E.; Chasapi, S.; Spyroulias, G.; Koutsovasilis, A.; Bairaktari, E.; et al. Effect of Dapagliflozin on Urine Metabolome in Patients with Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2021, 106, 1269–1283. [Google Scholar] [CrossRef]
- Ferrannini, E.; Natali, A.; Camastra, S.; Nannipieri, M.; Mari, A.; Adam, K.P.; Milburn, M.V.; Kastenmuller, G.; Adamski, J.; Tuomi, T.; et al. Early metabolic markers of the development of dysglycemia and type 2 diabetes and their physiological significance. Diabetes 2013, 62, 1730–1737. [Google Scholar] [CrossRef] [Green Version]
- Alesi, S.; Ghelani, D.; Rassie, K.; Mousa, A. Metabolomic Biomarkers in Gestational Diabetes Mellitus: A Review of the Evidence. Int. J. Mol. Sci. 2021, 22, 5512. [Google Scholar] [CrossRef] [PubMed]
- Huxtable, R.J. Physiological actions of taurine. Physiol. Rev. 1992, 72, 101–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nisoli, E.; Aquilani, R.; D’Antona, G. Amino Acid Supplements and Diabetes. In Bioactive Food as Dietary Interventions for Diabetes; Elsevier: Amsterdam, The Netherlands, 2013; pp. 83–95. [Google Scholar] [CrossRef]
- Bustamante, J.; Alonso, F.J.; Lobo, M.V.; Gine, E.; Tamarit-Rodriguez, J.; Solis, J.M.; Martin del Rio, R. Taurine levels and localization in pancreatic islets. Adv. Exp. Med. Biol. 1998, 442, 65–69. [Google Scholar] [CrossRef]
- Song, Y.; Manson, J.E.; Tinker, L.; Howard, B.V.; Kuller, L.H.; Nathan, L.; Rifai, N.; Liu, S. Insulin sensitivity and insulin secretion determined by homeostasis model assessment and risk of diabetes in a multiethnic cohort of women: The Women’s Health Initiative Observational Study. Diabetes Care 2007, 30, 1747–1752. [Google Scholar] [CrossRef] [Green Version]
- McManus, M.L.; Churchwell, K.B.; Strange, K. Regulation of cell volume in health and disease. N. Engl. J. Med. 1995, 333, 1260–1266. [Google Scholar] [CrossRef] [PubMed]
- Handler, J.S.; Kwon, H.M. Kidney cell survival in high tonicity. Comp. Biochem. Physiol. A Physiol. 1997, 117, 301–306. [Google Scholar] [CrossRef]
- Hansen, S.H. The role of taurine in diabetes and the development of diabetic complications. Diabetes Metab. Res. Rev. 2001, 17, 330–346. [Google Scholar] [CrossRef]
- Devamanoharan, P.S.; Ali, A.H.; Varma, S.D. Prevention of lens protein glycation by taurine. Mol. Cell. Biochem. 1997, 177, 245–250. [Google Scholar] [CrossRef]
- Park, S.; Kim, M.Y.; Baik, S.H.; Woo, J.T.; Kwon, Y.J.; Daily, J.W.; Park, Y.M.; Yang, J.H.; Kim, S.H. Gestational diabetes is associated with high energy and saturated fat intakes and with low plasma visfatin and adiponectin levels independent of prepregnancy BMI. Eur. J. Clin. Nutr. 2013, 67, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Seghieri, G.; Tesi, F.; Bianchi, L.; Loizzo, A.; Saccomanni, G.; Ghirlanda, G.; Anichini, R.; Franconi, F. Taurine in women with a history of gestational diabetes. Diabetes Res. Clin. Pract. 2007, 76, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Guasch-Ferre, M.; Hruby, A.; Toledo, E.; Clish, C.B.; Martinez-Gonzalez, M.A.; Salas-Salvado, J.; Hu, F.B. Metabolomics in Prediabetes and Diabetes: A Systematic Review and Meta-analysis. Diabetes Care 2016, 39, 833–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A branched-chain amino acid-related metabolic signature that differentiates obese and lean humans and contributes to insulin resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef] [Green Version]
- Butte, N.F.; Hsu, H.W.; Thotathuchery, M.; Wong, W.W.; Khoury, J.; Reeds, P. Protein metabolism in insulin-treated gestational diabetes. Diabetes Care 1999, 22, 806–811. [Google Scholar] [CrossRef]
- Metzger, B.E.; Phelps, R.L.; Freinkel, N.; Navickas, I.A. Effects of gestational diabetes on diurnal profiles of plasma glucose, lipids, and individual amino acids. Diabetes Care 1980, 3, 402–409. [Google Scholar] [CrossRef]
- Cetin, I.; de Santis, M.S.; Taricco, E.; Radaelli, T.; Teng, C.; Ronzoni, S.; Spada, E.; Milani, S.; Pardi, G. Maternal and fetal amino acid concentrations in normal pregnancies and in pregnancies with gestational diabetes mellitus. Am. J. Obstet. Gynecol. 2005, 192, 610–617. [Google Scholar] [CrossRef]
- Mock, D.M.; Stratton, S.L.; Horvath, T.D.; Bogusiewicz, A.; Matthews, N.I.; Henrich, C.L.; Dawson, A.M.; Spencer, H.J.; Owen, S.N.; Boysen, G.; et al. Urinary excretion of 3-hydroxyisovaleric acid and 3-hydroxyisovaleryl carnitine increases in response to a leucine challenge in marginally biotin-deficient humans. J. Nutr. 2011, 141, 1925–1930. [Google Scholar] [CrossRef] [Green Version]
- van Hove, J.L.; Rutledge, S.L.; Nada, M.A.; Kahler, S.G.; Millington, D.S. 3-Hydroxyisovalerylcarnitine in 3-methylcrotonyl-CoA carboxylase deficiency. J. Inherit. Metab. Dis. 1995, 18, 592–601. [Google Scholar] [CrossRef]
- Nilsen, M.S.; Jersin, R.A.; Ulvik, A.; Madsen, A.; McCann, A.; Svensson, P.A.; Svensson, M.K.; Nedrebo, B.G.; Gudbrandsen, O.A.; Tell, G.S.; et al. 3-Hydroxyisobutyrate, A Strong Marker of Insulin Resistance in Type 2 Diabetes and Obesity That Modulates White and Brown Adipocyte Metabolism. Diabetes 2020, 69, 1903–1916. [Google Scholar] [CrossRef]
- Andersson-Hall, U.; Gustavsson, C.; Pedersen, A.; Malmodin, D.; Joelsson, L.; Holmang, A. Higher Concentrations of BCAAs and 3-HIB Are Associated with Insulin Resistance in the Transition from Gestational Diabetes to Type 2 Diabetes. J. Diabetes Res. 2018, 2018, 4207067. [Google Scholar] [CrossRef] [Green Version]
- Menni, C.; Fauman, E.; Erte, I.; Perry, J.R.; Kastenmuller, G.; Shin, S.Y.; Petersen, A.K.; Hyde, C.; Psatha, M.; Ward, K.J.; et al. Biomarkers for type 2 diabetes and impaired fasting glucose using a nontargeted metabolomics approach. Diabetes 2013, 62, 4270–4276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsay, K.L.; Hellmuth, C.; Uhl, O.; Buss, C.; Wadhwa, P.D.; Koletzko, B.; Entringer, S. Longitudinal Metabolomic Profiling of Amino Acids and Lipids across Healthy Pregnancy. PLoS ONE 2015, 10, e0145794. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Alvarez, M.I.; Diaz-Ramos, A.; Berdasco, M.; Cobb, J.; Planet, E.; Cooper, D.; Pazderska, A.; Wanic, K.; O’Hanlon, D.; Gomez, A.; et al. Early-onset and classical forms of type 2 diabetes show impaired expression of genes involved in muscle branched-chain amino acids metabolism. Sci. Rep. 2017, 7, 13850. [Google Scholar] [CrossRef] [Green Version]
- Floegel, A.; Stefan, N.; Yu, Z.; Muhlenbruch, K.; Drogan, D.; Joost, H.G.; Fritsche, A.; Haring, H.U.; Hrabe de Angelis, M.; Peters, A.; et al. Identification of serum metabolites associated with risk of type 2 diabetes using a targeted metabolomic approach. Diabetes 2013, 62, 639–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite profiles and the risk of developing diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef] [Green Version]
- Huynh, J.; Xiong, G.; Bentley-Lewis, R. A systematic review of metabolite profiling in gestational diabetes mellitus. Diabetologia 2014, 57, 2453–2464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Wurtz, P.; Auro, K.; Makinen, V.P.; Kangas, A.J.; Soininen, P.; Tiainen, M.; Tynkkynen, T.; Jokelainen, J.; Santalahti, K.; et al. Metabolic profiling of pregnancy: Cross-sectional and longitudinal evidence. BMC Med. 2016, 14, 205. [Google Scholar] [CrossRef] [Green Version]
- Leitner, M.; Fragner, L.; Danner, S.; Holeschofsky, N.; Leitner, K.; Tischler, S.; Doerfler, H.; Bachmann, G.; Sun, X.; Jaeger, W.; et al. Combined Metabolomic Analysis of Plasma and Urine Reveals AHBA, Tryptophan and Serotonin Metabolism as Potential Risk Factors in Gestational Diabetes Mellitus (GDM). Front. Mol. Biosci. 2017, 4, 84. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Morris, S.M., Jr. Arginine metabolism: Nitric oxide and beyond. Biochem. J. 1998, 336, 1–17. [Google Scholar] [CrossRef]
- Burzynska-Pedziwiatr, I.; Jankowski, A.; Kowalski, K.; Sendys, P.; Zieleniak, A.; Cypryk, K.; Zurawska-Klis, M.; Wozniak, L.A.; Bukowiecka-Matusiak, M. Associations of Arginine with Gestational Diabetes Mellitus in a Follow-Up Study. Int. J. Mol. Sci. 2020, 21, 7811. [Google Scholar] [CrossRef]
- Marti, I.L.A.A.; Reith, W. Arginine-dependent immune responses. Cell Mol. Life Sci. 2021, 78, 5303–5324. [Google Scholar] [CrossRef] [PubMed]
- Ignarro, L.J.; Buga, G.M.; Wood, K.S.; Byrns, R.E.; Chaudhuri, G. Endothelium-derived relaxing factor produced and released from artery and vein is nitric oxide. Proc. Natl. Acad. Sci. USA 1987, 84, 9265–9269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo-Castrejon, M.; Migoya-Graue, A.; Meraz-Cruz, N.; Vadillo-Ortega, F. The role of L-arginine in human pregnancy. Agro Food Ind. Hi-Tech 2011, 22, 26–28. [Google Scholar]
- Guzman-Gutierrez, E.; Arroyo Zúñiga, P.; Pardo, F.; Leiva, A.; Sobrevia, L. The Adenosine–Insulin Signaling Axis in the Fetoplacental Endothelial Dysfunction in Gestational Diabetes. In Gestational Diabetes-Causes, Diagnosis and Treatment; InTech Open: Rijeka, Croatia, 2013. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Mahdi, A.; Tratsiakovich, Y.; Zahoran, S.; Kovamees, O.; Nordin, F.; Uribe Gonzalez, A.E.; Alvarsson, M.; Ostenson, C.G.; Andersson, D.C.; et al. Erythrocytes from Patients with Type 2 Diabetes Induce Endothelial Dysfunction Via Arginase I. J. Am. Coll. Cardiol. 2018, 72, 769–780. [Google Scholar] [CrossRef]
- Ueland, P.M. Choline and betaine in health and disease. J. Inherit. Metab. Dis. 2011, 34, 3–15. [Google Scholar] [CrossRef]
- Yan, J.; Jiang, X.; West, A.A.; Perry, C.A.; Malysheva, O.V.; Devapatla, S.; Pressman, E.; Vermeylen, F.; Stabler, S.P.; Allen, R.H.; et al. Maternal choline intake modulates maternal and fetal biomarkers of choline metabolism in humans. Am. J. Clin. Nutr. 2012, 95, 1060–1071. [Google Scholar] [CrossRef] [Green Version]
- Velasquez, M.T.; Ramezani, A.; Manal, A.; Raj, D.S. Trimethylamine N-Oxide: The Good, the Bad and the Unknown. Toxins 2016, 8, 326. [Google Scholar] [CrossRef] [Green Version]
- Nanobashvili, K.; Jack-Roberts, C.; Bretter, R.; Jones, N.; Axen, K.; Saxena, A.; Blain, K.; Jiang, X. Maternal Choline and Betaine Supplementation Modifies the Placental Response to Hyperglycemia in Mice and Human Trophoblasts. Nutrients 2018, 10, 1507. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Zhong, C.; Li, S.; Sun, T.; Huang, H.; Chen, X.; Zhu, Y.; Hu, X.; Peng, X.; Zhang, X.; et al. Plasma concentration of trimethylamine-N-oxide and risk of gestational diabetes mellitus. Am. J. Clin. Nutr. 2018, 108, 603–610. [Google Scholar] [CrossRef] [Green Version]
- Raubenheimer, P.J.; Nyirenda, M.J.; Walker, B.R. A choline-deficient diet exacerbates fatty liver but attenuates insulin resistance and glucose intolerance in mice fed a high-fat diet. Diabetes 2006, 55, 2015–2020. [Google Scholar] [CrossRef] [Green Version]
- Kalagi, N.A.; Thota, R.N.; Stojanovski, E.; Alburikan, K.A.; Garg, M.L. Association between Plasma Trimethylamine N-Oxide Levels and Type 2 Diabetes: A Case Control Study. Nutrients 2022, 14, 93. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Zhao, M.; Wang, F.; Li, Y.; Li, H.; Zheng, J.; Chen, X.; Zhao, X.; Ji, L.; Gao, X.; et al. Integrating Choline and Specific Intestinal Microbiota to Classify Type 2 Diabetes in Adults: A Machine Learning Based Metagenomics Study. Front. Endocrinol. 2022, 13, 906310. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Control Group | GDM Group | p Value |
|---|---|---|---|
| Ν | 77 | 62 | |
| Age (years) | 32.69 ± 4.72 | 34.27 ± 5.07 | NS |
| Gestational Week | 28.06 ± 1.29 | 27.91 ± 1.22 | NS |
| BMI (kg/m2) | 25.61 ± 5.01 | 26 ± 4.98 | NS |
| Systolic Blood pressure (mm Hg) | 109.21 ± 13.15 | 111.05 ± 11.74 | NS |
| Diastolic Blood pressure (mm Hg) | 59.74 ± 7.72 | 59.74 ± 12.07 | NS |
| Metabolic Parameters | |||
| Total cholesterol (mg/dL) | 251.08 ± 34.78 | 254 ± 48.45 | NS |
| Triglycerides (mg/dL) | 190.96 ± 66.85 | 191.15 ± 76.13 | NS |
| HDL- cholesterol (mg/dL) | 63.54 ± 12.02 | 64.40 ± 12.22 | NS |
| LDL- cholesterol (mg/dL) | 149 ± 32 | 154 ± 37.32 | NS |
| Fasting glucose (mg/dL) | 83.04 ± 5.60 | 93.19 ± 9.08 | <0.001 |
| Glucose 60′ (mg/dL) | 130.27 ± 26.39 | 177.48 ± 29.90 | <0.001 |
| Glucose 120′ (mg/dL) | 103.77 ± 24.31 | 129.26 ± 34.74 | <0.001 |
| HbA1c (%) | 4.98 ± 0.29 | 5.13 ± 0.34 | <0.01 |
| Metabolites (µM) | Control | GDM | p Value | |
|---|---|---|---|---|
| 1 | 2-Aminobutyrate | 14.54 ± 2.55 | 13.28 ± 3.51 | <0.05 |
| 2 | 2-Hydroxybutyrate | 39.05 ± 14.07 | 49.29 ± 17.78 | <0.001 |
| 3 | 2-Hydroxyisovalerate | 8.08 ± 2.31 | 8.80 ± 2.94 | NS |
| 4 | 2-Oxocaproate | 8.68 ± 2.69 | 8.59 ± 3.33 | NS |
| 5 | 2-Oxoglutarate | 13.44 ± 4.28 | 12.74 ± 3.85 | NS |
| 6 | 2-Oxoisocaproate | 20.62 ± 5.12 | 20.87 ± 6.56 | NS |
| 7 | 3-Hydroxybutyrate | 96.93 ± 83.45 | 145.15 ± 124.75 | <0.05 |
| 8 | 3-Hydroxyisobutyrate | 5.54 ± 1.85 | 6.40 ± 2.33 | <0.05 |
| 9 | 3-Hydroxyisovalerate | 4.04 ± 1.08 | 4.66 ± 1.60 | <0.05 |
| 10 | 3-Methyl-2-oxovalerate | 11.79 ± 2.73 | 10.43 ± 2.96 | <0.01 |
| 11 | Acetate | 46.86 ± 16.58 | 48.53 ± 18.55 | NS |
| 12 | Acetoacetate | 36.00 ± 29.43 | 56.79 ± 44.91 | <0.01 |
| 13 | Acetone | 16.91 ± 9.13 | 18.39 ± 11.77 | NS |
| 14 | Alanine | 462.08 ± 84.31 | 442.12 ± 108.42 | NS |
| 15 | Arginine | 125.55 ± 13.57 | 109.37 ± 6.13 | <0.001 |
| 16 | Asparagine | 58.36 ± 7.51 | 53.56 ± 6.70 | <0.01 |
| 17 | Aspartate | 36.04 ± 6.27 | 31.40 ± 7.38 | <0.001 |
| 18 | Betaine | 38.85 ± 9.45 | 37.76 ± 12.72 | NS |
| 19 | Carnitine | 32.14 ± 6.39 | 26.60 ± 6.92 | <0.001 |
| 20 | Choline | 18.40 ± 4.29 | 13.07 ± 3.16 | <0.001 |
| 21 | Citrate | 162.35 ± 35.27 | 172.10 ± 37.03 | NS |
| 22 | Creatine | 42.47 ± 12.49 | 40.83 ± 12.88 | NS |
| 23 | Dimethylamine | 4.29 ± 1.91 | 3.67 ± 1.96 | NS |
| 24 | Formate | 19.19 ± 7.58 | 17.13 ± 6.86 | NS |
| 25 | Glutamate | 98.30 ± 21.07 | 88.56 ± 24.95 | <0.05 |
| 26 | Glutamine | 440.13 ± 56.78 | 437.82 ± 69.07 | NS |
| 27 | Glycine | 292.86 ± 53.47 | 232.30 ± 54.93 | <0.001 |
| 28 | Histidine | 100.90 ± 24.27 | 86.83 ± 19.93 | <0.001 |
| 29 | Hypoxanthine | 19.15 ± 5.67 | 13.87 ± 6.43 | <0.001 |
| 30 | Inosine | 6.02 ± 0.82 | 5.79 ± 0.95 | NS |
| 31 | Isobutyrate | 9.19 ± 3.13 | 10.12 ± 3.28 | NS |
| 32 | Isoleucine | 57.26 ± 8.90 | 55.45 ± 10.87 | NS |
| 33 | Lactate | 2388.66 ± 727.39 | 2326.54 ± 819.48 | NS |
| 34 | Leucine | 120.97 ± 16.21 | 115.68 ± 22.86 | NS |
| 35 | Lysine | 170.45 ± 28.42 | 163.33 ± 32.25 | NS |
| 36 | Malonate | 14.31 ± 3.39 | 13.80 ± 4.43 | NS |
| 37 | Mannose | 120.14 ± 41.99 | 114.69 ± 40.03 | NS |
| 38 | Methanol | 52.10 ± 18.46 | 46.92 ± 19.88 | NS |
| 39 | Methionine | 28.38 ± 5.51 | 25.36 ± 5.61 | <0.01 |
| 40 | myo-Inositol | 30.01 ± 8.66 | 30.36 ± 9.14 | NS |
| 41 | N,N-Dimethylglycine | 2.29 ± 0.94 | 2.06 ± 0.98 | NS |
| 42 | O-Acetylcarnitine | 4.52 ± 1.41 | 4.70 ± 1.57 | NS |
| 43 | Ornithine | 31.79 ± 10.39 | 26.60 ± 9.46 | <0.01 |
| 44 | Phenylalanine | 81.21 ± 16.06 | 64.13 ± 14.29 | <0.001 |
| 45 | Proline | 173.11 ± 41.93 | 170.37 ± 48.63 | NS |
| 46 | Pyruvate | 119.01 ± 39.60 | 105.13 ± 32.50 | <0.05 |
| 47 | Sarcosine | 4.89 ± 1.52 | 3.79 ± 1.82 | <0.001 |
| 48 | Serine | 208.51 ± 33.88 | 189.89 ± 34.96 | <0.01 |
| 49 | Succinate | 7.04 ± 1.80 | 7.48 ± 2.36 | NS |
| 50 | Taurine | 141.41 ± 39.58 | 123.81 ± 43.30 | <0.05 |
| 51 | Threonine | 291.61 ± 59.23 | 310.07 ± 79.27 | NS |
| 52 | Trimethylamine N-oxide | 35.45 ± 9.49 | 31.61 ± 9.90 | <0.05 |
| 53 | Tryptophan | 50.31 ± 16.18 | 38.72 ± 16.55 | <0.001 |
| 54 | Tyrosine | 50.85 ± 8.26 | 47.39 ± 12.06 | NS |
| 55 | Valine | 193.94 ± 27.28 | 198.05 ± 37.34 | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spanou, L.; Dimou, A.; Kostara, C.E.; Bairaktari, E.; Anastasiou, E.; Tsimihodimos, V. A Study of the Metabolic Pathways Affected by Gestational Diabetes Mellitus: Comparison with Type 2 Diabetes. Diagnostics 2022, 12, 2881. https://doi.org/10.3390/diagnostics12112881
Spanou L, Dimou A, Kostara CE, Bairaktari E, Anastasiou E, Tsimihodimos V. A Study of the Metabolic Pathways Affected by Gestational Diabetes Mellitus: Comparison with Type 2 Diabetes. Diagnostics. 2022; 12(11):2881. https://doi.org/10.3390/diagnostics12112881
Chicago/Turabian StyleSpanou, Loukia, Aikaterini Dimou, Christina E. Kostara, Eleni Bairaktari, Eleni Anastasiou, and Vasilis Tsimihodimos. 2022. "A Study of the Metabolic Pathways Affected by Gestational Diabetes Mellitus: Comparison with Type 2 Diabetes" Diagnostics 12, no. 11: 2881. https://doi.org/10.3390/diagnostics12112881
APA StyleSpanou, L., Dimou, A., Kostara, C. E., Bairaktari, E., Anastasiou, E., & Tsimihodimos, V. (2022). A Study of the Metabolic Pathways Affected by Gestational Diabetes Mellitus: Comparison with Type 2 Diabetes. Diagnostics, 12(11), 2881. https://doi.org/10.3390/diagnostics12112881