
The Expression of the Senescence-Associated Biomarker Lamin B1 in Human Breast Cancer

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Patients Clinicopathological Characteristics
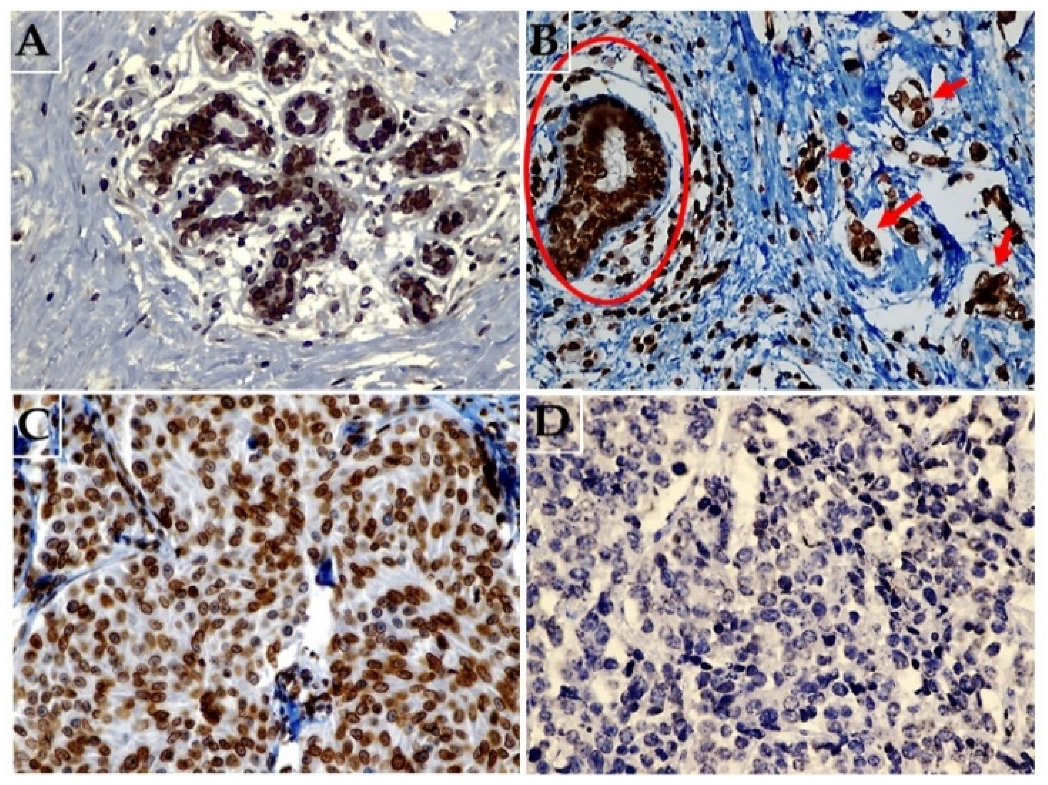
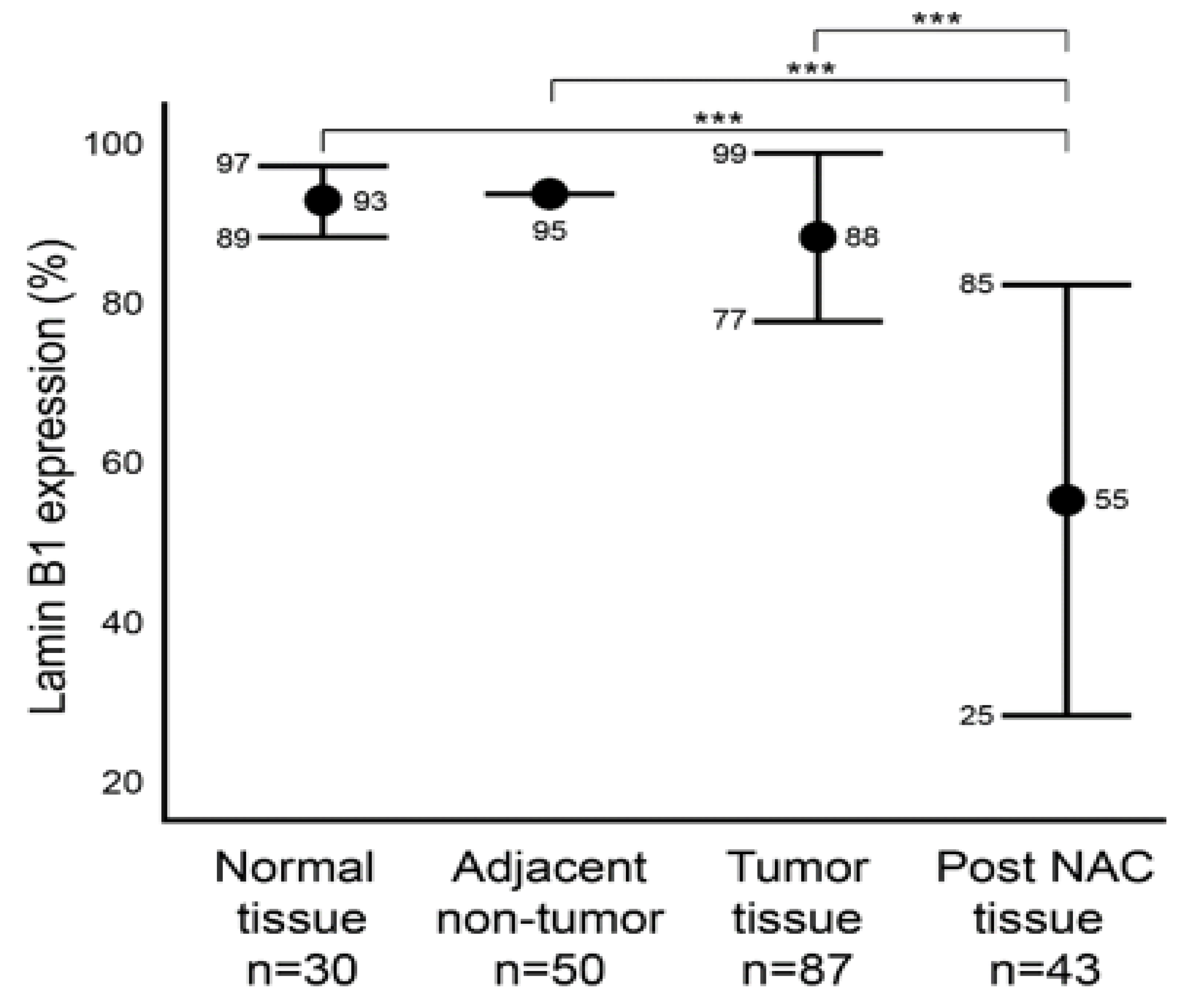
2.2. Lamin B1 Immunoexpression Levels in Normal Breast Epithelium, Primary Invasive Breast Carcinoma Exposed or Not Exposed to NAC
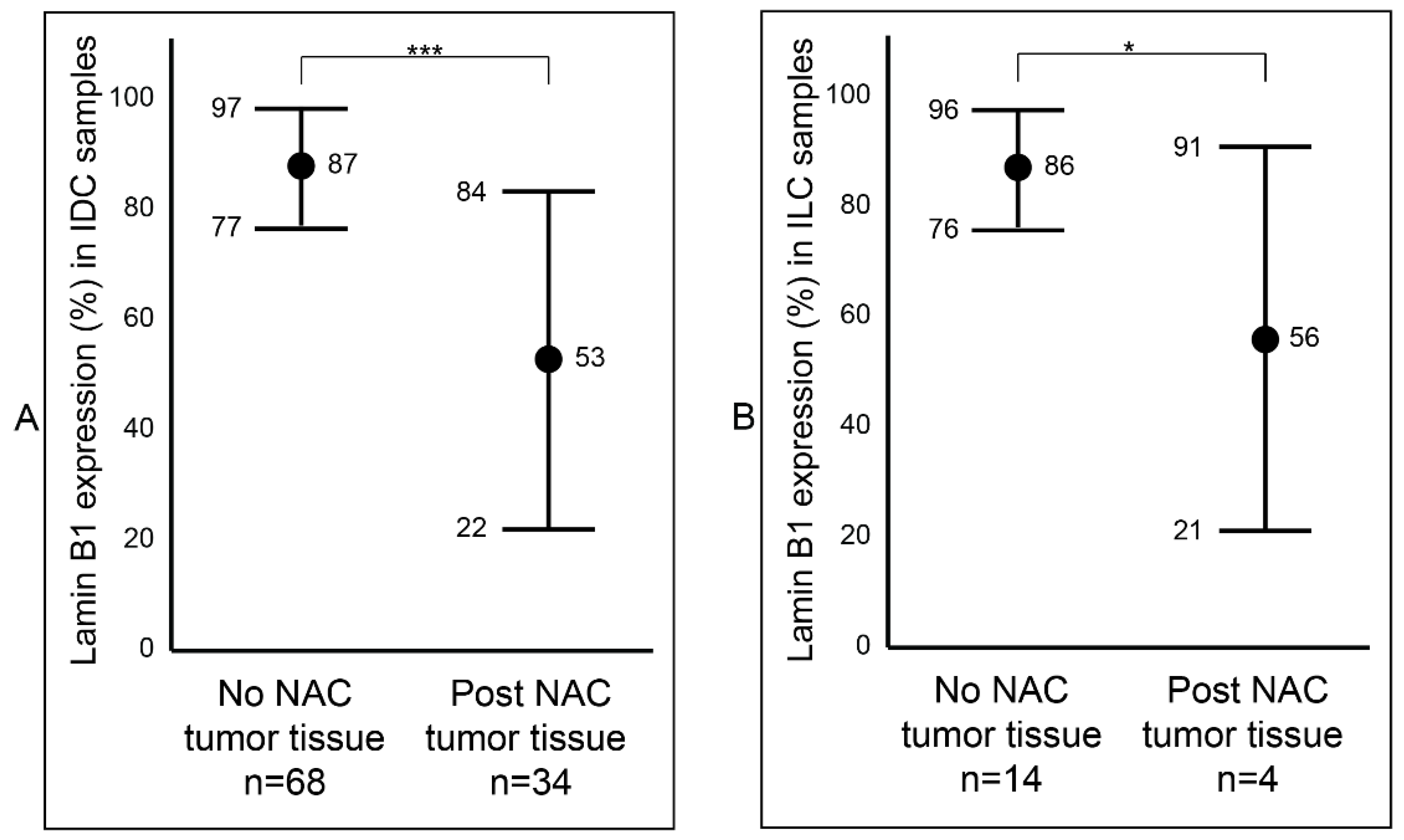
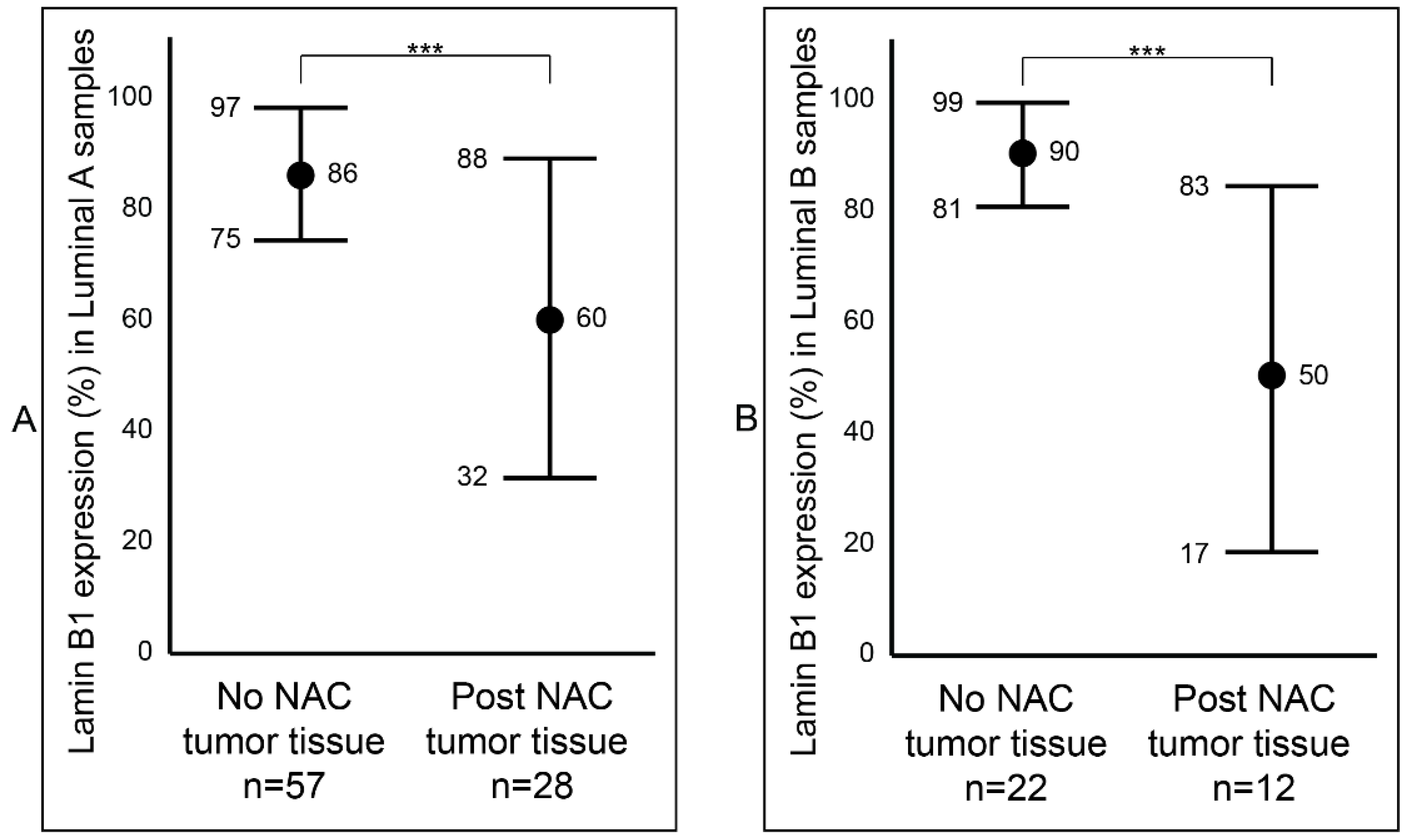
2.3. Association between Lamin B1 and Clinicopathological Characteristics of Patients with Invasive Breast Carcinoma
2.4. Analysis of Lamin B1 Protein Expression in Association with the Clinicopathological Characteristics of Patients with Invasive Breast Carcinoma
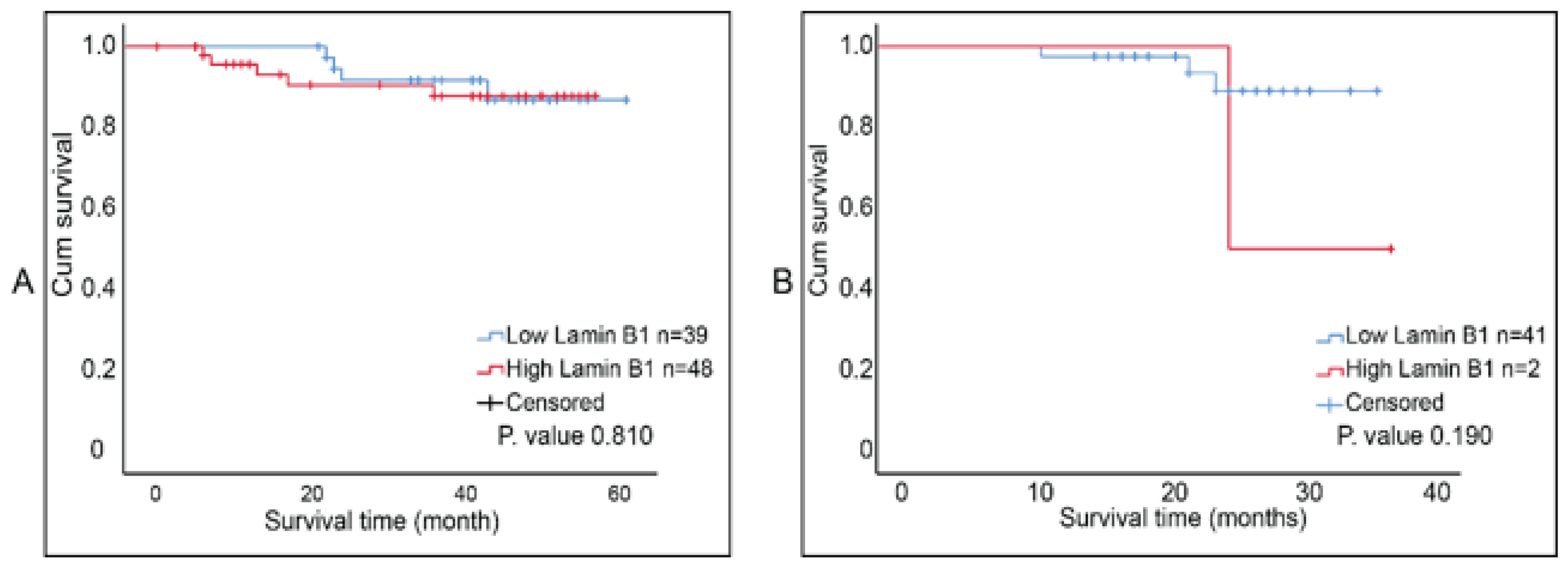
2.5. Survival Analysis of Lamin B1 Expression in Invasive Breast Carcinoma
3. Discussion
4. Materials and Methods
4.1. Sample
4.2. Immunohistochemical Procedures
4.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Wang, B.; Kohli, J.; Demaria, M. Senescent Cells in Cancer Therapy: Friends or Foes? Trends Cancer 2020, 6, 838–857. [Google Scholar] [CrossRef] [PubMed]
- Saleh, T.; Bloukh, S.; Carpenter, V.J.; Alwohoush, E.; Bakeer, J.; Darwish, S.; Azab, B.; Gewirtz, D.A. Therapy-Induced Senescence: An “Old” Friend Becomes the Enemy. Cancers 2020, 12, 822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasanna, P.G.; Citrin, D.E.; Hildesheim, J.; Ahmed, M.M.; Venkatachalam, S.; Riscuta, G.; Xi, D.; Zheng, G.; van Deursen, J.; Goronzy, J.; et al. Therapy-Induced Senescence: Opportunities to Improve Anticancer Therapy. J. Natl. Cancer Inst. 2021, 113, 1285–1298. [Google Scholar] [CrossRef]
- Sharpless, N.E.; Sherr, C.J. Forging a signature of in vivo senescence. Nat. Rev. Cancer 2015, 15, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Short, S.; Fielder, E.; Miwa, S.; von Zglinicki, T. Senolytics and senostatics as adjuvant tumour therapy. EBioMedicine 2019, 41, 683–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.T.; Lee, Y.S.; Cho, K.A.; Park, S.C. Adjustment of the lysosomal-mitochondrial axis for control of cellular senescence. Ageing Res. Rev. 2018, 47, 176–182. [Google Scholar] [CrossRef]
- You, R.; Dai, J.; Zhang, P.; Barding, G.A.; Raftery, D. Dynamic metabolic response to adriamycin-induced senescence in breast cancer cells. Metabolites 2018, 8, 95. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.Y.; Han, J.A.; Im, J.S.; Morrone, A.; Johung, K.; Goodwin, E.C.; Kleijer, W.J.; DiMaio, D.; Hwang, E.S. Senescence-associated β-galactosidase is lysosomal β-galactosidase. Aging Cell 2006, 5, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Kurz, D.J.D.J.; Decary, S.; Hong, Y.; Erusalimsky, J.D.J.D. Senescence-associated (beta)-galactosidase reflects an increase in lysosomal mass during replicative ageing of human endothelial cells. J. Cell Sci. 2000, 113, 3613–3622. [Google Scholar] [CrossRef]
- Di Micco, R.; Sulli, G.; Dobreva, M.; Liontos, M.; Botrugno, O.A.; Gargiulo, G.; dal Zuffo, R.; Matti, V.; D’Ario, G.; Montani, E.; et al. Interplay between oncogene-induced DNA damage response and heterochromatin in senescence and cancer. Nat. Cell Biol. 2011, 13, 292–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coppé, J.P.; Patil, C.K.; Rodier, F.; Sun, Y.Y.; Muñoz, D.P.; Goldstein, J.; Nelson, P.S.; Desprez, P.-Y.Y.; Campisi, J.; Coppe, J.-P.; et al. Senescence-Associated Secretory Phenotypes Reveal Cell-Nonautonomous Functions of Oncogenic RAS and the p53 Tumor Suppressor. Aging Cell 2008, 6, 2853–2868. [Google Scholar] [CrossRef]
- Chandra, T.; Ewels, P.A.; Schoenfelder, S.; Furlan-Magaril, M.; Wingett, S.W.; Kirschner, K.; Thuret, J.Y.; Andrews, S.; Fraser, P.; Reik, W. Global reorganization of the nuclear landscape in senescent cells. Cell Rep. 2015, 10, 471–483. [Google Scholar] [CrossRef]
- Wydner, K.L.; McNeil, J.A.; Lin, F.; Worman, H.J.; Lawrence, J.B. Chromosomal assignment of human nuclear envelope protein genes LMNA, LMNB1, and LBR by fluorescence in situ hybridization. Genomics 1996, 32, 474–478. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.-J.; Lee, J.M.; Yang, S.H.; Young, S.G.; Fong, L.G. Nuclear lamins in the brain—New insights into function and regulation. Mol. Neurobiol. 2013, 47, 290–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Wang, J.; Chan, K.M.; Tjia, W.M.; Deng, W.; Guan, X.; Huang, J.; Li, K.M.; Chau, P.Y.; Chen, D.J.; et al. Genomic instability in laminopathy-based premature aging. Nat. Med. 2005, 11, 780–785. [Google Scholar] [CrossRef]
- Shimi, T.; Butin-Israeli, V.; Adam, S.A.; Hamanaka, R.B.; Goldman, A.E.; Lucas, C.A.; Shumaker, D.K.; Kosak, S.T.; Chandel, N.S.; Goldman, R.D. The role of nuclear lamin B1 in cell proliferation and senescence. Genes Dev. 2011. [Google Scholar] [CrossRef] [Green Version]
- Camps, J.; Erdos, M.R.; Ried, T. The role of lamin B1 for the maintenance of nuclear structure and function. Nucleus 2015, 6, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Fracchia, A.; Asraf, T.; Salmon-Divon, M.; Gerlitz, G. Increased Lamin B1 Levels Promote Cell Migration by Altering Perinuclear Actin Organization. Cells 2020, 9, 2161. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Han, Y.C.; Han, Q.; Liang, Y.; Luo, Y.; Wei, L.; Yan, T.; Yang, Y.; Liu, S.L.; Wang, E.H. Lamin B2 promotes the malignant phenotype of non-small cell lung cancer cells by upregulating dimethylation of histone 3 lysine 9. Exp. Cell Res. 2020, 393, 112090. [Google Scholar] [CrossRef]
- Freund, A.; Laberge, R.-M.R.M.; Demaria, M.; Campisi, J. Lamin B1 loss is a senescence-associated biomarker. Mol. Biol. Cell 2012, 23, 2066–2075. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.P.; Donahue, G.; Otte, G.L.; Capell, B.C.; Nelson, D.M.; Cao, K.; Aggarwala, V.; Cruickshanks, H.A.; Rai, T.S.; McBryan, T.; et al. Lamin B1 depletion in senescent cells triggers large-scale changes in gene expression and the chromatin landscape. Genes Dev. 2013, 27, 1787–1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wazir, U.; Ahmed, M.H.; Bridger, J.M.; Harvey, A.; Jiang, W.G.; Sharma, A.K.; Mokbel, K. The clinicopathological significance of lamin A/C, lamin B1 and lamin B receptor mRNA expression in human breast cancer. Cell. Mol. Biol. Lett. 2013, 18, 595–611. [Google Scholar] [CrossRef] [PubMed]
- Coradeghini, R.; Barboro, P.; Rubagotti, A.; Boccardo, F.; Parodi, S.; Carmignani, G.; D’Arrigo, C.; Patrone, E.; Balbi, C. Differential expression of nuclear lamins in normal and cancerous prostate tissues. Oncol. Rep. 2006, 15, 609–613. [Google Scholar] [CrossRef]
- Li, L.; Du, Y.; Kong, X.; Li, Z.; Jia, Z.; Cui, J.; Gao, J.; Wang, G.; Xie, K. Lamin B1 is a novel therapeutic target of betulinic acid in pancreatic cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2013, 19, 4651–4661. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Xu, M.Z.; Poon, R.T.; Day, P.J.; Luk, J.M. Circulating Lamin B1 (LMNB1) biomarker detects early stages of liver cancer in patients. J. Proteome Res. 2010, 9, 70–78. [Google Scholar] [CrossRef]
- Radspieler, M.M.; Schindeldecker, M.; Stenzel, P.; Försch, S.; Tagscherer, K.E.; Herpel, E.; Hohenfellner, M.; Hatiboglu, G.; Roth, W.; Macher-Goeppinger, S. Lamin-B1 is a senescence-associated biomarker in clear-cell renal cell carcinoma. Oncol. Lett. 2019. [Google Scholar] [CrossRef] [Green Version]
- Garvalov, B.K.; Muhammad, S.; Dobreva, G. Lamin B1 in cancer and aging. Aging 2019, 11, 7336–7338. [Google Scholar] [CrossRef] [PubMed]
- Evangelisti, C.; Rusciano, I.; Mongiorgi, S.; Ramazzotti, G.; Lattanzi, G.; Manzoli, L.; Cocco, L.; Ratti, S. The wide and growing range of lamin B-related diseases: From laminopathies to cancer. Cell. Mol. Life Sci. 2022, 79, 126. [Google Scholar] [CrossRef]
- Amin, M.B.; Greene, F.L.; Edge, S.B.; Compton, C.C.; Gershenwald, J.E.; Brookland, R.K.; Meyer, L.; Gress, D.M.; Byrd, D.R.; Winchester, D.P. The Eighth Edition AJCC Cancer Staging Manual: Continuing to build a bridge from a population-based to a more “personalized” approach to cancer staging. CA Cancer J. Clin. 2017, 67, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Gorgoulis, V.; Adams, P.D.; Alimonti, A.; Bennett, D.C.; Bischof, O.; Bishop, C.; Campisi, J.; Collado, M.; Evangelou, K.; Ferbeyre, G.; et al. Cellular Senescence: Defining a Path Forward. Cell 2019, 179, 813–827. [Google Scholar] [CrossRef] [PubMed]
- Poele, R.H.; Okorokov, A.L.; Jardine, L.; Cummings, J.; Joel, S.P.; te Poele, R.H.; Okorokov, A.L.; Jardine, L.; Cummings, J.; Joel, S.P. DNA Damage Is Able to Induce Senescence in Tumor Cells in Vitro and in Vivo. Cancer Res. 2002, 62, 1876–1883. [Google Scholar]
- González-Gualda, E.; Baker, A.G.; Fruk, L.; Muñoz-Espín, D. A guide to assessing cellular senescence in vitro and in vivo. FEBS J. 2021, 288, 56–80. [Google Scholar] [CrossRef] [PubMed]
- Roberson, R.S.; Kussick, S.J.; Vallieres, E.; Chen, S.Y.J.; Wu, D.Y. Escape from therapy-induced accelerated cellular senescence in p53-null lung cancer cells and in human lung cancers. Cancer Res. 2005, 65, 2795–2803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidi, R.; Pasello, G.; Opitz, I.; Soltermann, A.; Tutic, M.; Rehrauer, H.; Weder, W.; Stahel, R.A.; Felley-Bosco, E. Induction of senescence markers after neo-adjuvant chemotherapy of malignant pleural mesothelioma and association with clinical outcome: An exploratory analysis. Eur. J. Cancer 2011, 47, 326–332. [Google Scholar] [CrossRef]
- Sanoff, H.K.; Deal, A.M.; Krishnamurthy, J.; Torrice, C.; Dillon, P.; Sorrentino, J.; Ibrahim, J.G.; Jolly, T.A.; Williams, G.; Carey, L.A.; et al. Effect of Cytotoxic Chemotherapy on Markers of Molecular Age in Patients With Breast Cancer. J. Natl. Cancer Inst. 2014, 106, dju057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotarelo, C.L.; Schad, A.; Kirkpatrick, C.J.; Sleeman, J.P.; Springer, E.; Schmidt, M.; Thaler, S. Detection of cellular senescence within human invasive breast carcinomas distinguishes different breast tumor subtypes. Oncotarget 2016, 7, 74846–748597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, T.; Tyutyunyk-Massey, L.; Gewirtz, D.A.D.A. Tumor Cell Escape from Therapy-Induced Senescence as a Model of Disease Recurrence after Dormancy. Cancer Res. 2019, 79, 1044–1046. [Google Scholar] [CrossRef] [Green Version]
- Saleh, T.; Tyutyunyk-Massey, L.; Murray, G.F.G.F.; Alotaibi, M.R.M.R.M.R.; Kawale, A.S.A.S.A.S.; Elsayed, Z.; Henderson, S.C.S.C.; Yakovlev, V.; Elmore, L.W.L.W.L.W.; Toor, A.; et al. Tumor cell escape from therapy-induced senescence. Biochem. Pharmacol. 2019, 162, 202–212. [Google Scholar] [CrossRef]
- Milanovic, M.; Fan, D.N.Y.; Belenki, D.; Däbritz, J.H.M.; Zhao, Z.; Yu, Y.; Dörr, J.R.; Dimitrova, L.; Lenze, D.; Monteiro Barbosa, I.A.; et al. Senescence-associated reprogramming promotes cancer stemness. Nature 2018, 553, 96–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Fang, J.; Chen, J. Tumor cell senescence response produces aggressive variants. Cell Death Discov. 2017, 3, 17049. [Google Scholar] [CrossRef] [PubMed]
- Duy, C.; Li, M.; Teater, M.; Meydan, C.; Garrett-bakelman, F.E.; Lee, T.C.; Chin, C.R.; Durmaz, C.; Kawabata, K.C.; Mitsiades, C.S.; et al. Chemotherapy induces senescence-like resilient cells capable of initiating AML recurrence. Cancer Discov. 2021, 11, 1542–1561. [Google Scholar] [CrossRef] [PubMed]
- Demaria, M.; Leary, M.N.O.; Chang, J.; Shao, L.; Liu, S.; Alimirah, F.; Koenig, K.; Le, C.; Mitin, N.; Deal, A.M.; et al. Cellular Senescence Promotes Adverse Effects of Chemotherapy and Cancer Relapse. Cancer Discov. 2017, 7, 165–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, T.; Carpenter, V.; Tyutyunyk-Massey, L.; Murray, G.; Leverson, J.; Souers, A.; Alotaibi, M.; Faber, A.; Reed, J.; Harada, H.; et al. Clearance of therapy-induced senescent tumor cells by the senolytic ABT-263 via interference with BCL-X L -BAX Interaction. Mol. Oncol. 2020, 14, 1–16. [Google Scholar] [CrossRef]
- Muñoz-Espín, D.; Rovira, M.; Galiana, I.; Giménez, C.; Lozano-Torres, B.; Paez-Ribes, M.; Llanos, S.; Chaib, S.; Muñoz-Martín, M.; Ucero, A.C.; et al. A versatile drug delivery system targeting senescent cells. EMBO Mol. Med. 2018, 10, e9355. [Google Scholar] [CrossRef]
- Shahbandi, A.; Rao, S.G.; Anderson, A.Y.; Frey, W.D.; Olayiwola, J.O.; Ungerleider, N.A.; Jackson, J.G. BH3 mimetics selectively eliminate chemotherapy-induced senescent cells and improve response in TP53 wild-type breast cancer. Cell Death Differ. 2020, 27, 3097–3116. [Google Scholar] [CrossRef]
- Wang, C.; Vegna, S.; Jin, H.; Benedict, B.; Lieftink, C.; Ramirez, C.; de Oliveira, R.L.; Morris, B.; Gadiot, J.; Wang, W.; et al. Inducing and exploiting vulnerabilities for the treatment of liver cancer. Nature 2019, 574, 268–272. [Google Scholar] [CrossRef]
- Carpenter, V.J.; Saleh, T.; Gewirtz, D.A. Senolytics for cancer therapy: Is all that glitters really gold? Cancers 2021, 13, 723. [Google Scholar] [CrossRef]
- Yu, Z.Y.; Jiang, X.Y.; Zhao, R.R.; Luo, C.J.; Ren, Y.X.; Ma, Z.J.; Ye, H.L.; Shi, W.G.; Wang, C.; Jiao, Z.Y. Lamin B1 deficiency promotes malignancy and predicts poor prognosis in gastric cancer. Neoplasma 2020, 67, 1303–1313. [Google Scholar] [CrossRef]
- Li, W.; Li, X.; Li, X.; Li, M.; Yang, P.; Wang, X.; Li, L.; Yang, B. Lamin B1 Overexpresses in Lung Adenocarcinoma and Promotes Proliferation in Lung Cancer Cells via AKT Pathway. Oncotargets Ther. 2020, 13, 3129–3139. [Google Scholar] [CrossRef] [Green Version]
- Saleh, T.; Alhesa, A.; Al-Balas, M.; Abuelaish, O.; Mansour, A.; Awad, H.; El-Sadoni, M.; Carpenter, V.J.; Azab, B. Expression of therapy-induced senescence markers in breast cancer samples upon incomplete response to neoadjuvant chemotherapy. Biosci. Rep. 2021, 41, BSR20210079. [Google Scholar] [CrossRef] [PubMed]
- Saleh, T.; El-sadoni, M.; Alhesa, A.; Awad, H.; Jaradat, M.; Al-hazaimeh, M.; Dawoud, R.; Mryyian, A.; Azab, B. Expression of Senescence and Apoptosis Biomarkers in Synchronous Bilateral Breast Cancer: A Case Report. Curr. Oncol. 2021, 28, 3836. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group B (n = 87) n (%) | Group C (n = 43) n (%) | ||
|---|---|---|---|
| Breast Cancer Subtype | IDC | 68 (78%) | 34 (79%) |
| ILC | 14 (16%) | 4 (9%) | |
| Other | 5 (6%) | 5 (12%) | |
| ER | Positive | 67 (77%) | 38 (88%) |
| Negative | 20 (23%) | 5 (12%) | |
| PR | Positive | 66 (76%) | 35 (81%) |
| Negative | 21 (24%) | 8 (19%) | |
| HER2 | Positive | 23 (26%) | 14 (33%) |
| Negative | 64 (74%) | 29 (67%) | |
| Luminal A | ER+/PR+ & HER2− | 57 (66%) | 28 (65%) |
| Luminal B | ER+/PR+ & HER2+ | 22 (25%) | 12 (28%) |
| HER2+ | ER−/PR− & HER2+ | 1 (1%) | 2 (5%) |
| TNBC | ER−/PR− & HER2− | 7 (8%) | 1 (2%) |
| Stage | Stage I | 36 (41%) | 22 (51%) |
| Stage II | 24 (28%) | 12 (28%) | |
| Stage III | 27 (31%) | 9 (21%) | |
| Grade | Grade 1 | 7 (8%) | 1 (2%) |
| Grade 2 | 42 (48%) | 23 (%53) | |
| Grade 3 | 38 (44%) | 19 (45%) | |
| Lymphovascular invasion | Present | 52 (60%) | 25 (58%) |
| Not identified | 35 (40%) | 18 (42%) | |
| Lymph node involvement | Positive | 59 (70%) | 30 (70%) |
| Negative | 28 (30%) | 13 (30%) |
| Lamin B1 Low | Lamin B1 High | p Value | ||
|---|---|---|---|---|
| Luminal A (n = 57) | ER+/PR+ & HER2− | 29 (51%) | 28 (49%) | 0.366 |
| Luminal B (n = 22) | ER+/PR+ & HER2+ | 7 (32%) | 15 (68%) | |
| HER2+ (n = 1) | ER−/PR− & HER2+ | 0 (0%) | 1 (100%) | |
| TNBC (n = 7) | ER−/PR− & HER2− | 3 (43%) | 4 (57%) | |
| Stage | Stage I (n = 36) | 14 (39%) | 22 (61%) | 0.506 |
| Stage II (n = 24) | 13 (54%) | 11 (46%) | ||
| Stage III (n = 27) | 12 (44%) | 15 (56%) | ||
| Lymphovascular Invasion | Positive (n = 52) | 22 (42%) | 30 (58%) | 0.565 |
| Negative (n = 35) | 17 (49%) | 18 (51%) | ||
| Lymph node involvement | Positive (n = 28) | 16 (57%) | 12 (43%) | 0.112 |
| Negative (n = 59) | 23 (39%) | 36 (61%) |
| Lamin B1 Low | Lamin B1 High | p Value | ||
|---|---|---|---|---|
| Luminal A (n = 28) | ER+/PR+ & HER2− | 26 (93%) | 2 (7%) | 0.771 |
| Luminal B (n = 12) | ER+/PR+ & HER2+ | 12 (100%) | 0 (0%) | |
| HER2+ (n = 1) | ER−/PR− & HER2+ | 1 (100%) | 0 (0%) | |
| TNBC (n = 2) | ER−/PR− & HER2− | 2 (100%) | 0 (0%) | |
| Stage | Stage I (n = 22) | 22 (100%) | 0 (0%) | 0.319 |
| Stage II (n = 12) | 11 (92%) | 1 (8%) | ||
| Stage III (n = 9) | 8 (89%) | 1 (11%) | ||
| Lymphovascular Invasion | Positive (n = 25) | 24 (96%) | 1 (4%) | 0.881 |
| Negative (n = 18) | 17 (94%) | 1 (6%) | ||
| Lymph node involvement | Positive (n = 30) | 28 (93%) | 2 (7%) | 0.340 |
| Negative (n = 13) | 13 (100%) | 0 (0%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saleh, T.; Alhesa, A.; El-Sadoni, M.; Abu Shahin, N.; Alsharaiah, E.; Al Shboul, S.; Awad, H.; Bloukh, S.; Al-Balas, M.; Alsalem, M.; et al. The Expression of the Senescence-Associated Biomarker Lamin B1 in Human Breast Cancer. Diagnostics 2022, 12, 609. https://doi.org/10.3390/diagnostics12030609
Saleh T, Alhesa A, El-Sadoni M, Abu Shahin N, Alsharaiah E, Al Shboul S, Awad H, Bloukh S, Al-Balas M, Alsalem M, et al. The Expression of the Senescence-Associated Biomarker Lamin B1 in Human Breast Cancer. Diagnostics. 2022; 12(3):609. https://doi.org/10.3390/diagnostics12030609
Chicago/Turabian StyleSaleh, Tareq, Ahmad Alhesa, Mohammed El-Sadoni, Nisreen Abu Shahin, Elham Alsharaiah, Sofian Al Shboul, Heyam Awad, Sarah Bloukh, Mahmoud Al-Balas, Mohammad Alsalem, and et al. 2022. "The Expression of the Senescence-Associated Biomarker Lamin B1 in Human Breast Cancer" Diagnostics 12, no. 3: 609. https://doi.org/10.3390/diagnostics12030609
APA StyleSaleh, T., Alhesa, A., El-Sadoni, M., Abu Shahin, N., Alsharaiah, E., Al Shboul, S., Awad, H., Bloukh, S., Al-Balas, M., Alsalem, M., Azab, B., & Aladily, T. N. (2022). The Expression of the Senescence-Associated Biomarker Lamin B1 in Human Breast Cancer. Diagnostics, 12(3), 609. https://doi.org/10.3390/diagnostics12030609