
Homeobox Gene Expression Dysregulation as Potential Diagnostic and Prognostic Biomarkers in Bladder Cancer

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Method of Search
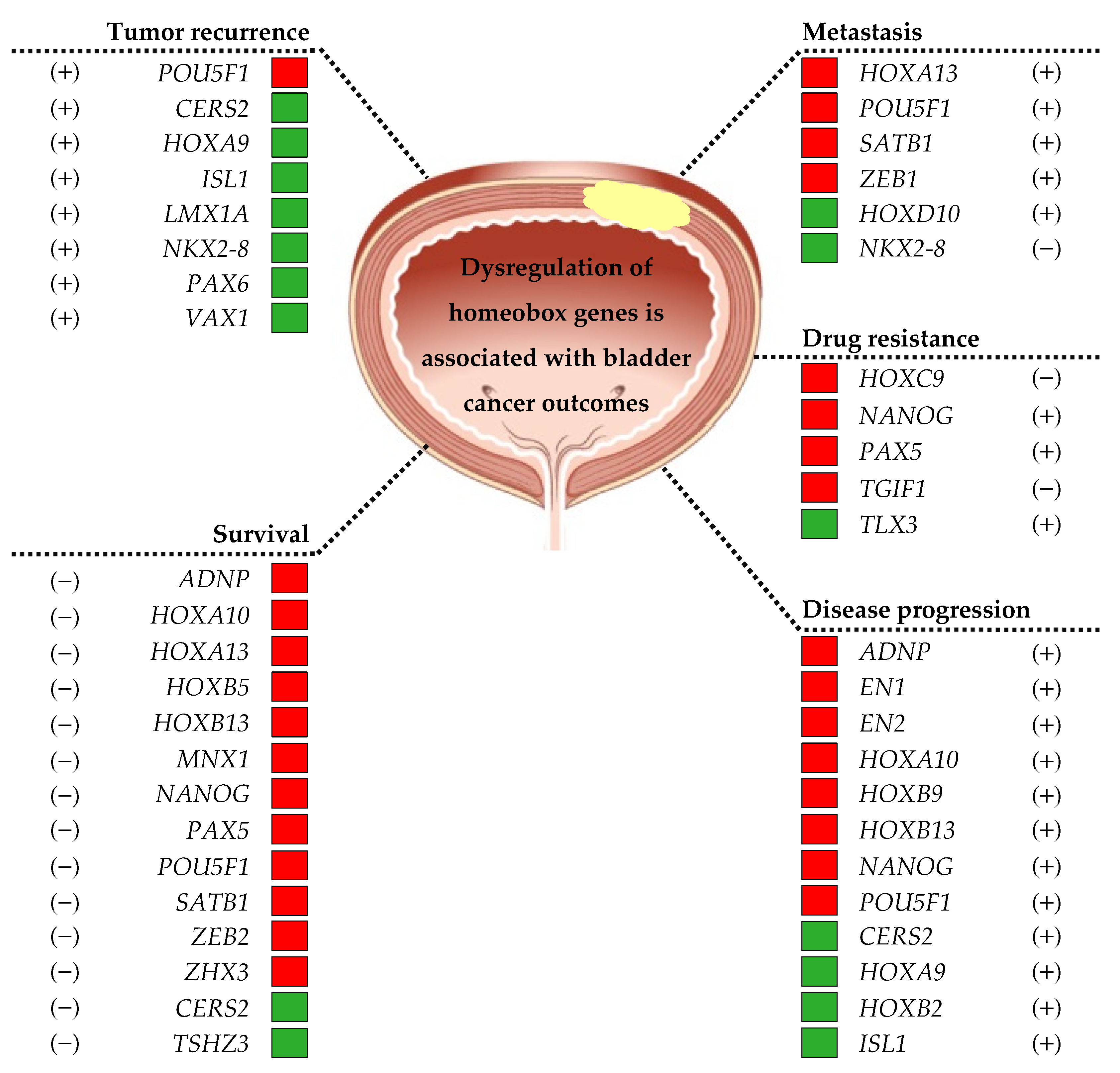
3. Homeobox Genes in Bladder Cancer
3.1. The ANTP Homeobox Class
3.1.1. CDX2
3.1.2. EN1 and HOXB9
3.1.3. EN2
3.1.4. HOXA1
3.1.5. HOXB2
3.1.6. HOXB5
3.1.7. HOXC4, HOXC5, HOXC6, HOXC11, and HOXD11
3.1.8. HOXC8
3.1.9. HOXA9
3.1.10. HOXC9
3.1.11. HOXA10
3.1.12. HOXA11
3.1.13. HOXD10
3.1.14. HOXA13
3.1.15. HOXB13
3.1.16. MNX1
3.1.17. BARX2
3.1.18. NANOG
3.1.19. NANOGP8
3.1.20. NKX2-8
3.1.21. NKX6-2
3.1.22. TLX1
3.1.23. TLX3
3.1.24. VAX1
3.1.25. VAX2
3.2. The CERS Homeobox Class
CERS2
3.3. The CUT Homeobox Class
3.3.1. CUX1
3.3.2. ONECUT2
3.3.3. SATB1
3.4. The LIM Homeobox Classes
ISL1 and LHX5
3.5. The POU Homeobox Class
POU5F1
3.6. The PRD Homeobox Class
3.6.1. ALX4
3.6.2. DUXAP10
3.6.3. OTX1
3.6.4. PAX2
3.6.5. PAX5
3.6.6. PAX6
3.6.7. PAX8
3.6.8. PRRX1
3.7. The PROS Homeobox Class
PROX1
3.8. The SINE Homeobox Class
SIX4
3.9. The TALE Homeobox Class
TGIF1
3.10. The ZF Homeobox Class
3.10.1. ADNP
3.10.2. ZEB1
3.10.3. ZEB2
3.10.4. TSHZ3
3.10.5. ZHX3
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gorski, D.H.; Walsh, K. The Role of Homeobox Genes in Vascular Remodeling and Angiogenesis. Circ. Res. 2000, 87, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Mark, M.; Rijli, F.M.; Chambon, P. Homeobox Genes in Embryogenesis and Pathogenesis. Pediatr. Res. 1997, 42, 421–429. [Google Scholar] [CrossRef] [PubMed]
- McGinnis, W.; Krumlauf, R. Homeobox genes and axial patterning. Cell 1992, 68, 283–302. [Google Scholar] [CrossRef]
- Quiring, R.; Walldorf, U.; Kloter, U.; Gehring, W.J. Homology of the eyeless Gene of Drosophila to the Small eye Gene in Mice and Aniridia in Humans. Science 1994, 265, 785–789. [Google Scholar] [CrossRef] [PubMed]
- James, J.A.; DavidG, D.G.; Thompson, A.; Lappin, T.R.J. Development and cancer: The HOX gene connection. In The Cancer Handbook, 2nd ed.; Alison, M., Ed.; John Wiley & Sons Ltd: Chichester, UK, 2007; pp. 298–310. [Google Scholar]
- Gao, A.C.; Lou, W.; Isaacs, J.T. Enhanced GBX2 expression stimulates growth of human prostate cancer cells via transcriptional up-regulation of the interleukin 6 gene. Clin. Cancer Res. 2000, 6, 493–497. [Google Scholar]
- Lu, S.; Liu, R.; Su, M.; Wei, Y.; Yang, S.; He, S.; Wang, X.; Qiang, F.; Chen, C.; Zhao, S.; et al. Overexpression of HOXC8 is Associated With Poor Prognosis in Epithelial Ovarian Cancer. Reprod. Sci. 2016, 23, 944–954. [Google Scholar] [CrossRef]
- Chen, Y.; Knösel, T.; Ye, F.; Pacyna-Gengelbach, M.; Deutschmann, N.; Petersen, I. Decreased PITX1 homeobox gene expression in human lung cancer. Lung Cancer 2007, 55, 287–294. [Google Scholar] [CrossRef]
- Li, Y.; Yang, X.H.; Fang, S.J.; Qin, C.F.; Sun, R.L.; Liu, Z.Y.; Jiang, B.Y.; Wu, X.; Li, G. HOXA7 stimulates human hepatocellular carcinoma proliferation through cyclin E1/CDK2. Oncol. Rep. 2014, 33, 990–996. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-J.; Zhu, Y.; Yuan, H.-X.; Zhang, J.-P.; Guo, J.-M.; Lin, Z.-M. Overexpression of HOXC11 homeobox gene in clear cell renal cell carcinoma induces cellular proliferation and is associated with poor prognosis. Tumor Biol. 2014, 36, 2821–2829. [Google Scholar] [CrossRef]
- Ji, M.; Feng, Q.; He, G.; Yang, L.; Tang, W.; Lao, X.; Zhu, D.; Lin, Q.; Xu, P.; Wei, Y.; et al. Silencing homeobox C6 inhibits colorectal cancer cell proliferation. Oncotarget 2016, 7, 29216–29227. [Google Scholar] [CrossRef] [Green Version]
- Makiyama, K.; Hamada, J.-I.; Takada, M.; Murakawa, K.; Takahashi, Y.; Tada, M.; Tamoto, E.; Shindo, G.; Matsunaga, A.; Teramoto, K.-I.; et al. Aberrant expression of HOX genes in human invasive breast carcinoma. Oncol. Rep. 2005, 13, 673–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.; Chen, Y.; Cheng, S.; Li, G.; Zhang, Z. Dysregulated expression of homebox gene HOXA13 is correlated with the poor prognosis in bladder cancer. Wien. Klin. Wochenschr. 2016, 129, 391–397. [Google Scholar] [CrossRef]
- Marra, L.; Cantile, M.; Scognamiglio, G.; Perdona, S.; La Mantia, E.; Cerrone, M.; Gigantino, V.; Cillo, C.; Caraglia, M.; Pignata, S.; et al. Deregulation of HOX B13 Expression in Urinary Bladder Cancer Progression. Curr. Med. Chem. 2013, 20, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Chen, H.; Parker, B.; Rubin, E.; Zhu, T.; Lee, J.S.; Argani, P.; Sukumar, S. HOXB7, a Homeodomain Protein, Is Overexpressed in Breast Cancer and Confers Epithelial-Mesenchymal Transition. Cancer Res. 2006, 66, 9527–9534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuel, S.; Naora, H. Homeobox gene expression in cancer: Insights from developmental regulation and deregulation. Eur. J. Cancer 2005, 41, 2428–2437. [Google Scholar] [CrossRef]
- Shah, N.; Sukumar, S. The Hox genes and their roles in oncogenesis. Nat. Rev. Cancer 2010, 10, 361–371. [Google Scholar] [CrossRef]
- Abate-Shen, C. Deregulated homeobox gene expression in cancer: Cause or consequence? Nat. Rev. Cancer 2002, 2, 777–785. [Google Scholar] [CrossRef]
- Joo, M.K.; Park, J.-J.; Chun, H.J. Impact of homeobox genes in gastrointestinal cancer. World J. Gastroenterol. 2016, 22, 8247–8256. [Google Scholar] [CrossRef]
- Nio, C.Y. Urothelial cell carcinoma in lower urinary tract: Conventional imaging techniques. In Imaging in Oncological Urology; Rosette, J.J.M.C.H., Manyak, M.J., Harisinghani, M.G., Wijkstra, H., Eds.; Springer: London, UK, 2009; pp. 177–184. [Google Scholar]
- Hurst, C.; Rosenberg, J.; Knowles, M. SnapShot: Bladder Cancer. Cancer Cell 2018, 34, 350–350.e1. [Google Scholar] [CrossRef]
- Tung, M.-C.; Wen, Y.-C.; Wang, S.-S.; Lin, Y.-W.; Chow, J.-M.; Yang, S.-F.; Chien, M.-H. Impact of Long Non-Coding RNA HOTAIR Genetic Variants on the Susceptibility and Clinicopathologic Characteristics of Patients with Urothelial Cell Carcinoma. J. Clin. Med. 2019, 8, 282. [Google Scholar] [CrossRef] [Green Version]
- Pstein, J.I.; Amin, M.B.; Reuter, V.R.; Mostofi, F.K. The World Health Organization/International Society of Urological Pathology consensus classification of urothelial (transitional cell) neoplasms of the urinary bladder. Bladder Consensus Conference Committee. Am. J. Surg. Pathol. 1998, 22, 1435–1448. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Gomez, J.; Madero, R.; Solsona, E.; Unda, M.; Martinez-Piñeiro, L.; Gonzalez, M.; Portillo, J.; Ojea, A.; Pertusa, C.; Rodriguez-Molina, J.; et al. Predicting Nonmuscle Invasive Bladder Cancer Recurrence and Progression in Patients Treated With Bacillus Calmette-Guerin: The CUETO Scoring Model. J. Urol. 2009, 182, 2195–2203. [Google Scholar] [CrossRef] [PubMed]
- Sylvester, R.J.; van der Meijden, A.P.; Oosterlinck, W.; Witjes, J.A.; Bouffioux, C.; Denis, L.; Newling, D.W.; Kurth, K. Predicting Recurrence and Progression in Individual Patients with Stage Ta T1 Bladder Cancer Using EORTC Risk Tables: A Combined Analysis of 2596 Patients from Seven EORTC Trials. Eur. Urol. 2006, 49, 466–477. [Google Scholar] [CrossRef]
- Cheng, T.; Roth, B.; Choi, W.; Black, P.C.; Dinney, C.; McConkey, D.J. Fibroblast Growth Factor Receptors-1 and -3 Play Distinct Roles in the Regulation of Bladder Cancer Growth and Metastasis: Implications for Therapeutic Targeting. PLoS ONE 2013, 8, e57284. [Google Scholar] [CrossRef] [Green Version]
- Avritscher, E.B.; Cooksley, C.D.; Grossman, H.B.; Sabichi, A.L.; Hamblin, L.; Dinney, C.P.; Elting, L.S. Clinical model of lifetime cost of treating bladder cancer and associated complications. Urology 2006, 68, 549–553. [Google Scholar] [CrossRef]
- Chamie, K.; Saigal, C.S.; Lai, J.; Hanley, J.M.; Setodji, C.M.; Konety, B.R.; Litwin, M.S.; Project, T.U.D.I.A. Compliance with guidelines for patients with bladder cancer: Variation in the delivery of care. Cancer 2011, 117, 5392–5401. [Google Scholar] [CrossRef]
- Winn, B.; Tavares, R.; Matoso, A.; Noble, L.; Fanion, J.; Waldman, S.A.; Resnick, M.B. Expression of the intestinal biomarkers Guanylyl cyclase C and CDX2 in poorly differentiated colorectal carcinomas. Hum. Pathol. 2010, 41, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Gao, N.; White, P.; Kaestner, K.H. Establishment of Intestinal Identity and Epithelial-Mesenchymal Signaling by Cdx2. Dev. Cell 2009, 16, 588–599. [Google Scholar] [CrossRef] [Green Version]
- De Lott, B.L.B.; Morrison, C.; Suster, S.; Cohn, D.E.; Frankel, W.L. CDX2 Is a Useful Marker of Intestinal-Type Differentiation: A Tissue Microarray–Based Study of 629 Tumors From Various Sites. Arch. Pathol. Lab. Med. 2005, 129, 1100–1105. [Google Scholar] [CrossRef] [PubMed]
- Kaimaktchiev, V.; Terracciano, L.; Tornillo, L.; Spichtin, H.; Stoios, D.; Bundi, M.; Korcheva, V.; Mirlacher, M.; Loda, M.; Sauter, G.; et al. The homeobox intestinal differentiation factor CDX2 is selectively expressed in gastrointestinal adenocarcinomas. Mod. Pathol. 2004, 17, 1392–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werling, R.W.; Yaziji, H.; Bacchi, C.E.; Gown, A.M. CDX2, a Highly Sensitive and Specific Marker of Adenocarcinomas of Intestinal Origin: An immunohistochemical survey of 476 primary and metastatic carcinomas. Am. J. Surg. Pathol. 2003, 27, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Sung, M.-T.; Zhang, S.; Lopez-Beltran, A.; Montironi, R.; Wang, M.; Davidson, D.D.; O Koch, M.; Cain, M.P.; Rink, R.C.; Cheng, L. Urothelial carcinoma following augmentation cystoplasty: An aggressive variant with distinct clinicopathological characteristics and molecular genetic alterations. Histopathology 2009, 55, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Suh, N.; Yang, X.J.; Tretiakova, M.S.; A Humphrey, P.; Wang, H.L. Value of CDX2, villin, and α-methylacyl coenzyme A racemase immunostains in the distinction between primary adenocarcinoma of the bladder and secondary colorectal adenocarcinoma. Mod. Pathol. 2005, 18, 1217–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, M.-T.; Lopez-Beltran, A.; Eble, J.N.; MacLennan, G.T.; Tan, P.-H.; Montironi, R.; Jones, T.D.; Ulbright, T.M.; E Blair, J.; Cheng, L. Divergent pathway of intestinal metaplasia and cystitis glandularis of the urinary bladder. Mod. Pathol. 2006, 19, 1395–1401. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Beltran, A.; Jimenez, R.E.; Montironi, R.; Patriarca, C.; Blanca, A.; Menendez, C.L.; Algaba, F.; Cheng, L. Flat urothelial carcinoma in situ of the bladder with glandular differentiation. Hum. Pathol. 2011, 42, 1653–1659. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, M.; Li, H.; Zou, D. Ruguo key genes and tumor driving factors identification of bladder cancer based on the RNA-seq profile. OncoTargets Ther. 2016, 9, 2717–2723. [Google Scholar] [CrossRef] [Green Version]
- Morgan, R.; Bryan, R.T.; Javed, S.; Launchbury, F.; Zeegers, M.P.; Cheng, K.; James, N.D.; Wallace, D.M.A.; Hurst, C.D.; Ward, D.G.; et al. Expression of Engrailed-2 (EN2) protein in bladder cancer and its potential utility as a urinary diagnostic biomarker. Eur. J. Cancer 2013, 49, 2214–2222. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, H.; Lai, C.; Su, Z.; Heng, B.; Gao, S. Repression of engrailed 2 inhibits the proliferation and invasion of human bladder cancer in vitro and in vivo. Oncol. Rep. 2015, 33, 2319–2330. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Duan, Q.; Gan, L.; Li, W.; Yang, J.; Huang, G. microRNA-27b inhibits cell proliferation and invasion in bladder cancer by targeting engrailed-2. Biosci. Rep. 2021, 41, BSR20201000. [Google Scholar] [CrossRef]
- Elamin, A.A.; Klunkelfuß, S.; Kämpfer, S.; Oehlmann, W.; Stehr, M.; Smith, C.; Simpson, G.R.; Morgan, R.; Pandha, H.; Singh, M. A Specific Blood Signature Reveals Higher Levels of S100A12: A Potential Bladder Cancer Diagnostic Biomarker Along With Urinary Engrailed-2 Protein Detection. Front. Oncol. 2020, 9, 1484. [Google Scholar] [CrossRef] [Green Version]
- De Paoli, M.; Gogalic, S.; Sauer, U.; Preininger, C.; Pandha, H.; Simpson, G.; Horvath, A.; Marquette, C. Multiplatform Biomarker Discovery for Bladder Cancer Recurrence Diagnosis. Dis. Markers 2016, 2016, 4591910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gogalic, S.; Sauer, U.; Doppler, S.; Heinzel, A.; Perco, P.; Lukas, A.; Simpson, G.; Pandha, H.; Horvath, A.; Preininger, C. Validation of a protein panel for the noninvasive detection of recurrent non-muscle invasive bladder cancer. Biomarkers 2017, 22, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.-S.; Toyokawa, G.; Daigo, Y.; Hayami, S.; Masuda, K.; Ikawa, N.; Yamane, Y.; Maejima, K.; Tsunoda, T.; Field, H.I.; et al. The JmjC domain-containing histone demethylase KDM3A is a positive regulator of the G1/S transition in cancer cells via transcriptional regulation of the HOXA1 gene. Int. J. Cancer 2011, 131, E179–E189. [Google Scholar] [CrossRef] [PubMed]
- Marsit, C.J.; Houseman, E.A.; Christensen, B.C.; Gagne, L.; Wrensch, M.R.; Nelson, H.H.; Wiemels, J.; Zheng, S.; Wiencke, J.K.; Andrew, A.S.; et al. Identification of Methylated Genes Associated with Aggressive Bladder Cancer. PLoS ONE 2010, 5, e12334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Li, S.; Cheng, X.; Du, P.; Yang, Y.; Jiang, W.G. HOXB2 is a Putative Tumour Promotor in Human Bladder Cancer. Anticancer Res. 2019, 39, 6915–6921. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Cai, Q.; Wang, W.; Huang, H.; Zeng, H.; He, W.; Deng, W.; Yu, H.; Chan, E.; Ng, C.-F.; et al. A MicroRNA-7 Binding Site Polymorphism in HOXB5 Leads to Differential Gene Expression in Bladder Cancer. PLoS ONE 2012, 7, e40127. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Wang, C.; Xiang, X.; Li, J.; Huang, J. Characterization of mRNA Expression and Endogenous RNA Profiles in Bladder Cancer Based on The Cancer Genome Atlas (TCGA) Database. Experiment 2019, 25, 3041–3060. [Google Scholar] [CrossRef]
- Cantile, M.; Cindolo, L.; Napodano, G.; Altieri, V.; Cillo, C. Hyperexpression of locus C genes in the HOX network is strongly associated in vivo with human bladder transitional cell carcinomas. Oncogene 2003, 22, 6462–6468. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Liu, X.; Dai, G.; Qu, L.; Tan, B.; Zhu, B.; Qi, F.; Gai, X.; Cheng, B. CircNT5E promotes the proliferation and migration of bladder cancer via sponging miR-502-5p. J. Cancer 2021, 12, 2430–2439. [Google Scholar] [CrossRef]
- Reinert, T.; Modin, C.; Castano, F.M.; Lamy, P.; Wojdacz, T.K.; Hansen, L.L.; Wiuf, C.; Borre, M.; Dyrskjøt, L.; Ørntoft, T.F. Comprehensive Genome Methylation Analysis in Bladder Cancer: Identification and Validation of Novel Methylated Genes and Application of These as Urinary Tumor Markers. Clin. Cancer Res. 2011, 17, 5582–5592. [Google Scholar] [CrossRef] [Green Version]
- Reinert, T.; Borre, M.; Christiansen, A.; Hermann, G.G.; Ørntoft, T.F.; Dyrskjøt, L. Diagnosis of Bladder Cancer Recurrence Based on Urinary Levels of EOMES, HOXA9, POU4F2, TWIST1, VIM, and ZNF154 Hypermethylation. PLoS ONE 2012, 7, e46297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Chen, S.; Wu, L.; Wu, Y.; Jiang, G.; Shao, J.; Chen, L.; Sun, J.; Na, R.; Wang, X.; et al. Identification of Cancer-Specific Methylation of Gene Combination for the Diagnosis of Bladder Cancer. J. Cancer 2019, 10, 6761–6766. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Jiang, G.; Zhang, N.; Liu, S.; Lin, X.; Perschon, C.; Zheng, S.; Ding, Q.; Wang, X.; Na, R.; et al. HOXA9, PCDH17, POU4F2, and ONECUT2 as a Urinary Biomarker Combination for the Detection of Bladder Cancer in Chinese Patients with Hematuria. Eur. Urol. Focus 2018, 6, 284–291. [Google Scholar] [CrossRef]
- Xylinas, E.; Hassler, M.R.; Zhuang, D.; Krzywinski, M.; Erdem, Z.; Robinson, B.D.; Elemento, O.; Clozel, T.; Shariat, S.F. An Epigenomic Approach to Improving Response to Neoadjuvant Cisplatin Chemotherapy in Bladder Cancer. Biomolecules 2016, 6, 37. [Google Scholar] [CrossRef]
- López, J.I.; Angulo, J.C.; Martín, A.; Sánchez-Chapado, M.; González-Corpas, A.; Colás, B.; Ropero, S. A DNA hypermethylation profile reveals new potential biomarkers for the evaluation of prognosis in urothelial bladder cancer. APMIS 2017, 125, 787–796. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Yoon, H.-Y.; Kim, J.S.; Kang, H.W.; Min, B.-D.; Kim, S.-K.; Ha, Y.-S.; Kim, I.Y.; Ryu, K.H.; Lee, S.-C.; et al. HOXA9, ISL1 and ALDH1A3 methylation patterns as prognostic markers for nonmuscle invasive bladder cancer: Array-based DNA methylation and expression profiling. Int. J. Cancer 2013, 133, 1135–1142. [Google Scholar] [CrossRef]
- Kitchen, M.O.; Bryan, R.T.; Haworth, K.E.; Emes, R.D.; Luscombe, C.; Gommersall, L.; Cheng, K.K.; Zeegers, M.P.; James, N.D.; Devall, A.J.; et al. Methylation of HOXA9 and ISL1 Predicts Patient Outcome in High-Grade Non-Invasive Bladder Cancer. PLoS ONE 2015, 10, e0137003. [Google Scholar] [CrossRef] [Green Version]
- Lv, L.; Li, Y.; Deng, H.; Zhang, C.; Pu, Y.; Qian, L.; Xiao, J.; Zhao, W.; Liu, Q.; Zhang, D.; et al. MiR-193a-3p promotes the multi-chemoresistance of bladder cancer by targeting the HOXC9 gene. Cancer Lett. 2015, 357, 105–113. [Google Scholar] [CrossRef]
- Liu, C.; Ge, M.; Ma, J.; Zhang, Y.; Zhao, Y.; Cui, T. Homeobox A10 promotes the proliferation and invasion of bladder cancer cells via regulation of matrix metalloproteinase-3. Oncol. Lett. 2019, 18, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.-P.; Xie, M.; Pan, W.-X.; Zhang, Z.-Y.; Li, W.-F. HOXA10 promotes the development of bladder cancer through regulating FOSL1. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2945–2954. [Google Scholar] [PubMed]
- Baffa, R.; Fassan, M.; Volinia, S.; O’Hara, B.; Liu, C.-G.; Palazzo, J.P.; Gardiman, M.; Rugge, M.; Gomella, L.G.; Croce, C.M.; et al. MicroRNA expression profiling of human metastatic cancers identifies cancer gene targets. J. Pathol. 2009, 219, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Luo, A.; Cai, Y.; Su, Q.; Ding, F.; Chen, H.; Liu, Z. MicroRNA-10b Promotes Migration and Invasion through KLF4 in Human Esophageal Cancer Cell Lines. J. Biol. Chem. 2010, 285, 7986–7994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Z.; Yuan, L.; Sun, N. Propofol exhibits a tumor-suppressive effect and regulates cell viability, migration and invasion in bladder carcinoma by targeting the microRNA-10b/HOXD10 signaling pathway. Oncol. Lett. 2019, 18, 6228–6236. [Google Scholar] [CrossRef] [PubMed]
- Scott, V.; Morgan, E.A.; Stadler, H.S. Genitourinary Functions of Hoxa13 and Hoxd13. J. Biochem. 2005, 137, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Holyoake, A.; O’Sullivan, P.; Pollock, R.; Best, T.; Watanabe, J.; Kajita, Y.; Matsui, Y.; Ito, M.; Nishiyama, H.; Kerr, N.; et al. Development of a Multiplex RNA Urine Test for the Detection and Stratification of Transitional Cell Carcinoma of the Bladder. Clin. Cancer Res. 2008, 14, 742–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, B.; Che, T.; Shi, B.; Guo, L.; Yin, Y.; Li, L.; Wang, J.; Yan, D.; Chen, Y. Screening and identification of specific markers for bladder transitional cell carcinoma from urine urothelial cells with suppressive subtractive hybridization and cDNA microarray. Can. Urol. Assoc. J. 2011, 5, e129–e137. [Google Scholar] [CrossRef]
- Guo, B.; Che, T.; Shi, B.; Guo, L.; Zhang, Z.; Li, L.; Cai, C.; Chen, Y. Interaction network analysis of differentially expressed genes and screening of cancer marker in the urine of patients with invasive bladder cancer. Int. J. Clin. Exp. Med. 2015, 8, 3619–3628. [Google Scholar]
- Chen, M.; Wu, R.; Li, G.; Liu, C.; Tan, L.; Xiao, K.; Ye, Y.; Qin, Z. Motor neuron and pancreas homeobox 1/HLXB9 promotes sustained proliferation in bladder cancer by upregulating CCNE1/2. J. Exp. Clin. Cancer Res. 2018, 37, 154. [Google Scholar] [CrossRef] [Green Version]
- Zuo, L.; Zhu, Y.; Han, J.; Liu, H. Circular RNA circSHPRH inhibits the malignant behaviors of bladder cancer by regulating the miR-942/BARX2 pathway. Aging 2022, 14, 1891–1909. [Google Scholar] [CrossRef]
- Amini, S.; Fathi, F.; Mobalegi, J.; Sofimajidpour, H.; Ghadimi, T. The expressions of stem cell markers: Oct4, Nanog, Sox2, nucleostemin, Bmi, Zfx, Tcl1, Tbx3, Dppa4, and Esrrb in bladder, colon, and prostate cancer, and certain cancer cell lines. Anat. Cell Biol. 2014, 47, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Migita, T.; Ueda, A.; Ohishi, T.; Hatano, M.; Seimiya, H.; Horiguchi, S.-I.; Koga, F.; Shibasaki, F. Epithelial–mesenchymal transition promotes SOX2 and NANOG expression in bladder cancer. Lab. Investig. 2017, 97, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, Z.; Yu, J.; Shi, J.Z.; Wang, C.; Fu, W.H.; Chen, Z.W.; Yang, J. Cancer stem-like cells contribute to cisplatin resistance and progression in bladder cancer. Cancer Lett. 2012, 322, 70–77. [Google Scholar] [CrossRef] [PubMed]
- A Akhir, M.K.; Hussin, H.; Veerakumarasivam, A.; Choy, C.S.; Abdullah, M.A.; Ghani, F.A. Immunohistochemical expression of NANOG in urothelial carcinoma of the bladder. Malays. J. Pathol. 2017, 39, 227–234. [Google Scholar]
- Gawlik-Rzemieniewska, N.; Galilejczyk, A.; Krawczyk, M.; Bednarek, I. Silencing expression of the NANOG gene and changes in migration and metastasis of urinary bladder cancer cells. Arch. Med. Sci. 2016, 12, 889–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Xie, W.; Gu, P.; Cai, Q.; Wang, B.; Xie, Y.; Dong, W.; He, W.; Zhong, G.; Lin, T.; et al. Upregulated WDR5 promotes proliferation, self-renewal and chemoresistance in bladder cancer via mediating H3K4 trimethylation. Sci. Rep. 2015, 5, srep08293. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, Z.; Srivastava, A.; Sankhwar, S.; Dalela, D.; Singh, V.; Zaidi, N.; Fatima, N.; Bano, I.; Anjum, S. Synergic effects of cancer stem cells markers, CD44 and embryonic stem cell transcription factor Nanog, on bladder cancer prognosis. Br. J. Biomed. Sci. 2019, 77, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, B.; Shao, J.; Wang, X. Chemotherapy sorting can be used to identify cancer stem cell populations. Mol. Biol. Rep. 2012, 39, 9955–9963. [Google Scholar] [CrossRef]
- Hepburn, A.C.; Veeratterapillay, R.; Williamson, S.C.; El-Sherif, A.; Sahay, N.; Thomas, H.D.; Mantilla, A.; Pickard, R.S.; Robson, C.N.; Heer, R. Side Population in Human Non-Muscle Invasive Bladder Cancer Enriches for Cancer Stem Cells That Are Maintained by MAPK Signalling. PLoS ONE 2012, 7, e50690. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Li, M.; Han, J.; Chen, B.; Wang, B.; Dai, J. NANOGP8 is a retrogene expressed in cancers. FEBS J. 2006, 273, 1723–1730. [Google Scholar] [CrossRef]
- Yu, C.; Zhang, Z.; Liao, W.; Zhao, X.; Liu, L.; Wu, Y.; Liu, Z.; Li, Y.; Zhong, Y.; Chen, K.; et al. The tumor-suppressor gene Nkx2.8 suppresses bladder cancer proliferation through upregulation of FOXO3a and inhibition of the MEK/ERK signaling pathway. Carcinogenesis 2012, 33, 678–686. [Google Scholar] [CrossRef]
- Yu, C.; Liu, Z.; Chen, Q.; Li, Y.; Jiang, L.; Zhang, Z.; Zhou, F. Nkx2.8 Inhibits Epithelial–Mesenchymal Transition in Bladder Urothelial Carcinoma via Transcriptional Repression of Twist1. Cancer Res. 2018, 78, 1241–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, W.; Bondaruk, J.; Jelinek, J.; Lotan, Y.; Liang, S.; Czerniak, B.; Issa, J.-P.J. Detection of Bladder Cancer Using Novel DNA Methylation Biomarkers in Urine Sediments. Cancer Epidemiol. Biomark. Prev. 2011, 20, 1483–1491. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; He, H.; Rui, W.; Xie, X.; Wang, D.; Zhu, Y. Long Non-Coding RNA BCAR4 Binds to miR-644a and Targets TLX1 to Promote the Progression of Bladder Cancer. OncoTargets Ther. 2020, 13, 2483–2490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tada, Y.; Yokomizo, A.; Shiota, M.; Tsunoda, T.; Plass, C.; Naito, S. Aberrant DNA methylation of T-cell leukemia, homeobox 3 modulates cisplatin sensitivity in bladder cancer. Int. J. Oncol. 2011, 39, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Guo, S.; Sun, J.; Huang, Z.; Zhu, T.; Zhang, H.; Gu, J.; He, Y.; Wang, W.; Ma, K.; et al. Methylcap-Seq Reveals Novel DNA Methylation Markers for the Diagnosis and Recurrence Prediction of Bladder Cancer in a Chinese Population. PLoS ONE 2012, 7, e35175. [Google Scholar] [CrossRef]
- Kitchen, M.O.; Bryan, R.T.; Emes, R.D.; Glossop, J.R.; Luscombe, C.; Cheng, K.K.; Zeegers, M.P.; James, N.D.; Devall, A.J.; Mein, C.A.; et al. Quantitative genome-wide methylation analysis of high-grade non-muscle invasive bladder cancer. Epigenetics 2016, 11, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Wang, H.; Yang, M.; Yang, D.; Zuo, Y.; Wang, J. Expression of a tumor-associated gene, LASS2, in the human bladder carcinoma cell lines BIU-87, T24, EJ and EJ-M3. Exp. Ther. Med. 2013, 5, 942–946. [Google Scholar] [CrossRef]
- Aldoghachi, A.; Baharudin, A.; Ahmad, U.; Chan, S.C.; Ong, T.A.; Yunus, R.; Razack, A.H.; Yusoff, K.; Veerakumarasivam, A. Evaluation of CERS2 Gene as a Potential Biomarker for Bladder Cancer. Dis. Markers 2019, 2019, 3875147–3875148. [Google Scholar] [CrossRef]
- Wang, H.; Wang, J.; Zuo, Y.; Ding, M.; Yan, R.; Yang, D.; Ke, C. Expression and prognostic significance of a new tumor metastasis suppressor gene LASS2 in human bladder carcinoma. Med. Oncol. 2011, 29, 1921–1927. [Google Scholar] [CrossRef]
- Xiao, S.; Chen, Y.; Luan, T.; Huang, Y.; Fu, S.; Zuo, Y.; Wang, H.; Wang, J. MicroRNA-20a Targeting LASS2 Promotes the Proliferation, Invasiveness and Migration of Bladder Cancer. Clin. Lab. 2021, 67, 1789–1798. [Google Scholar] [CrossRef]
- Fu, S.; Luan, T.; Jiang, C.; Huang, Y.; Li, N.; Wang, H.; Wang, J. miR-3622a promotes proliferation and invasion of bladder cancer cells by downregulating LASS2. Gene 2019, 701, 23–31. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, W.; Zuo, Y.; Ding, M.; Ke, C.; Yan, R.; Zhan, H.; Liu, J.; Wang, J. miR-9 promotes cell proliferation and inhibits apoptosis by targeting LASS2 in bladder cancer. Tumor Biol. 2015, 36, 9631–9640. [Google Scholar] [CrossRef]
- Luan, T.; Fu, S.; Huang, L.; Zuo, Y.; Ding, M.; Li, N.; Chen, J.; Wang, H.; Wang, J. MicroRNA-98 promotes drug resistance and regulates mitochondrial dynamics by targeting LASS2 in bladder cancer cells. Exp. Cell Res. 2018, 373, 188–197. [Google Scholar] [CrossRef]
- Wang, Z.; Shang, J.; Li, Z.; Li, H.; Zhang, C.; He, K.; Li, S.; Ju, W. PIK3CA Is Regulated by CUX1, Promotes Cell Growth and Metastasis in Bladder Cancer via Activating Epithelial-Mesenchymal Transition. Front. Oncol. 2020, 10, 536072. [Google Scholar] [CrossRef] [PubMed]
- Beukers, W.; Kandimalla, R.; van Houwelingen, D.; Kovacic, H.; Chin, J.-F.D.; Lingsma, H.F.; Dyrskjot, L.; Zwarthoff, E.C. The Use of Molecular Analyses in Voided Urine for the Assessment of Patients with Hematuria. PLoS ONE 2013, 8, e77657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Kessel, K.E.; Beukers, W.; Lurkin, I.; Der Made, A.Z.-V.; Van Der Keur, K.A.; Boormans, J.L.; Dyrskjøt, L.; Márquez, M.; Ørntoft, T.F.; Real, F.X.; et al. Validation of a DNA Methylation-Mutation Urine Assay to Select Patients with Hematuria for Cystoscopy. J. Urol. 2017, 197, 590–595. [Google Scholar] [CrossRef] [PubMed]
- Van Kessel, K.E.M.; Van Neste, L.; Lurkin, I.; Zwarthoff, E.C.; Van Criekinge, W. Evaluation of an Epigenetic Profile for the Detection of Bladder Cancer in Patients with Hematuria. J. Urol. 2016, 195, 601–607. [Google Scholar] [CrossRef]
- Choudhary, D.; Clement, J.M.; Choudhary, S.; Voznesensky, O.; Pilbeam, C.C.; Woolbright, B.L.; Taylor, J.A. SATB1 and bladder cancer: Is there a functional link? Urol. Oncol. Semin. Orig. Investig. 2018, 36, 93.e13–93.e21. [Google Scholar] [CrossRef]
- Hussein, S.; Fathi, A.; Abouhashem, N.S.; Amer, S.; Hemeda, M.; Mosaad, H. SATB-1 and Her2 as predictive molecular and immunohistochemical markers for urothelial cell carcinoma of the bladder. Cancer Biomark. 2021, 30, 249–259. [Google Scholar] [CrossRef]
- Wan, F.; Cheng, C.; Wang, Z.; Xiao, X.; Zeng, H.; Xing, S.; Chen, X.; Wang, J.; Li, S.; Zhang, Y.; et al. SATB1 Overexpression Regulates the Development and Progression in Bladder Cancer through EMT. PLoS ONE 2015, 10, e0117518. [Google Scholar] [CrossRef]
- Han, B.; Luan, L.; Xu, Z.; Wu, B. Expression and biological roles of SATB1 in human bladder cancer. Tumor Biol. 2013, 34, 2943–2949. [Google Scholar] [CrossRef] [PubMed]
- Akhir, M.K.A.M.; Choy, C.S.; Abdullah, M.A.; Ghani, F.A.; Veerakumarasivam, A.; Hussin, H. The Role of ISL1 and LHX5 LIM Homeobox Genes in Bladder Tumourigenesis. Malays. J. Med. Sci. 2020, 27, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-C.; Shieh, G.-S.; Wu, P.; Lin, C.-C.; Shiau, A.-L.; Wu, C.-L. Oct-3/4 Expression Reflects Tumor Progression and Regulates Motility of Bladder Cancer Cells. Cancer Res. 2008, 68, 6281–6291. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-L.; Shieh, G.-S.; Chang, C.-C.; Yo, Y.-T.; Su, C.-H.; Chang, M.-Y.; Huang, Y.-H.; Wu, P.; Shiau, A.-L. Tumor-Selective Replication of an Oncolytic Adenovirus Carrying Oct-3/4 Response Elements in Murine Metastatic Bladder Cancer Models. Clin. Cancer Res. 2008, 14, 1228–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, P.; Chen, J.; Wang, L.; Na, Y.; Kaku, H.; Ueki, H.; Sasaki, K.; Yamaguchi, K.; Zhang, K.; Saika, T.; et al. Implications of transcriptional factor, OCT-4, in human bladder malignancy and tumor recurrence. Med. Oncol. 2011, 29, 829–834. [Google Scholar] [CrossRef]
- Xu, K.; Zhu, Z.; Zeng, F.; Dong, J. Expression and significance of Oct4 in bladder cancer. J. Huazhong Univ. Sci. Technol. 2007, 27, 675–677. [Google Scholar] [CrossRef]
- Atlasi, Y.; Mowla, S.J.; Ziaee, S.A.; Bahrami, A.-R. OCT-4, an embryonic stem cell marker, is highly expressed in bladder cancer. Int. J. Cancer 2007, 120, 1598–1602. [Google Scholar] [CrossRef]
- Asadzadeh, J.; Asadi, M.H.; Shakhssalim, N.; Rafiee, M.-R.; Kalhor, H.R.; Tavallaei, M.; Mowla, S.J. A plausible anti-apoptotic role of up-regulated OCT4B1 in bladder tumors. Urol. J. 2012, 9, 574–581. [Google Scholar]
- Sedaghat, S.; Gheytanchi, E.; Asgari, M.; Roudi, R.; Keymoosi, H.; Madjd, Z. Expression of Cancer Stem Cell Markers OCT4 and CD133 in Transitional Cell Carcinomas. Appl. Immunohistochem. Mol. Morphol. 2017, 25, 196–202. [Google Scholar] [CrossRef]
- Lu, C.-S.; Shieh, G.-S.; Wang, C.-T.; Su, B.-H.; Su, Y.-C.; Chen, Y.-C.; Su, W.-C.; Wu, P.; Yang, W.-H.; Shiau, A.-L.; et al. Chemotherapeutics-induced Oct4 expression contributes to drug resistance and tumor recurrence in bladder cancer. Oncotarget 2016, 8, 30844–30858. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Dong, D.; Cheng, R.; Wang, Y.; Jiang, S.; Zhu, Y.; Fan, L.; Mao, X.; Gui, Y.; Li, Z.; et al. Aberrant expression of KPNA2 is associated with a poor prognosis and contributes to OCT4 nuclear transportation in bladder cancer. Oncotarget 2016, 7, 72767–72776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Zhu, T.; Wang, Z.; Zhang, H.; Qian, Z.; Xu, H.; Gao, B.; Wang, W.; Gu, L.; Meng, J.; et al. A Novel Set of DNA Methylation Markers in Urine Sediments for Sensitive/Specific Detection of Bladder Cancer. Clin. Cancer Res. 2007, 13, 7296–7304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, X.-Y.; Ma, L.; Chen, J.-F.; Yu, R.; Li, Y.; Yan, Z.; Cheng, Y.; Ma, Q. Knockdown of DUXAP10 inhibits proliferation and promotes apoptosis in bladder cancer cells via PI3K/Akt/mTOR signaling pathway. Int. J. Oncol. 2017, 52, 288–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Zuo, Z.; Lin, J.; Yang, C. Orthodenticle homeobox OTX1 is a potential prognostic biomarker for bladder cancer. Bioengineered 2021, 12, 6559–6571. [Google Scholar] [CrossRef]
- Muratovska, A.; Zhou, C.; He, S.; Goodyer, P.; Eccles, M.R. Paired-Box genes are frequently expressed in cancer and often required for cancer cell survival. Oncogene 2003, 22, 7989–7997. [Google Scholar] [CrossRef] [Green Version]
- Mhawech-Fauceglia, P.; Saxena, R.; Zhang, S.; Terracciano, L.; Sauter, G.; Chadhuri, A.; Herrmann, F.R.; Penetrante, R. Pax-5 immunoexpression in various types of benign and malignant tumours: A high-throughput tissue microarray analysis. J. Clin. Pathol. 2007, 60, 709–714. [Google Scholar] [CrossRef] [Green Version]
- Denzinger, S.; Burger, M.; Hammerschmied, C.G.; Wieland, W.F.; Hartmann, A.; Obermann, E.C.; Stoehr, R. Pax-5 protein expression in bladder cancer: A preliminary study that shows no correlation to grade, stage or clinical outcome. Pathology 2008, 40, 465–469. [Google Scholar] [CrossRef]
- Adshead, J.M.; Ogden, C.W.; Penny, M.; Stuart, E.T.; Kessling, A.M. The expression of PAX5 in human transitional cell carcinoma of the bladder: Relationship with de-differentiation. BJU Int. 2001, 83, 1039–1044. [Google Scholar] [CrossRef]
- Babjuk, M.; Soukup, V.; Mareš, J.; Dušková, J.; Pecen, L.; Pešl, M.; Pavlík, I.; DvorŘcek, J. Association of PAX5 expression with clinical outcome in patients with TaT1 transitional cell carcinoma of the bladder. Urology 2006, 67, 756–761. [Google Scholar] [CrossRef]
- Babjuk, M.; Soukup, V.; Mareš, J.; Dušková, J.; Sedláček, Z.; Trková, M.; Pecen, L.; Dvořáček, J.; Hanuš, T.; Kočvara, R.; et al. The expression of PAX5, p53 immunohistochemistry and p53 mutation analysis in superficial bladder carcinoma tissue. Correlation with pathological findings and clinical outcome. Int. Urol. Nephrol. 2002, 34, 495–501. [Google Scholar] [CrossRef]
- Dong, B.-W.; Zhang, W.-B.; Qi, S.-M.; Yan, C.-Y.; Gao, J. Transactivation of PTGS2 by PAX5 signaling potentiates cisplatin resistance in muscle-invasive bladder cancer cells. Biochem. Biophys. Res. Commun. 2018, 503, 2293–2300. [Google Scholar] [CrossRef] [PubMed]
- Hellwinkel, O.J.; Kedia, M.; Isbarn, H.; Budäus, L.; Friedrich, M.G. Methylation of the TPEF- and PAX6-promoters is increased in early bladder cancer and in normal mucosa adjacent to pTa tumours. BJU Int. 2008, 101, 753–757. [Google Scholar] [CrossRef]
- Sacristan, R.; Gonzalez, C.; Fernández-Gómez, J.M.; Fresno, F.; Escaf, S.; Sánchez-Carbayo, M. Molecular Classification of Non–Muscle-Invasive Bladder Cancer (pTa Low-Grade, pT1 Low-Grade, and pT1 High-Grade Subgroups) Using Methylation of Tumor-Suppressor Genes. J. Mol. Diagn. 2014, 16, 564–572. [Google Scholar] [CrossRef]
- Agundez, M.; Grau, L.; Palou, J.; Algaba, F.; Villavicencio, H.; Sanchez-Carbayo, M. Evaluation of the Methylation Status of Tumour Suppressor Genes for Predicting Bacillus Calmette-Guérin Response in Patients with T1G3 High-Risk Bladder Tumours. Eur. Urol. 2011, 60, 131–140. [Google Scholar] [CrossRef]
- Legesse, T.; Matoso, A.; Epstein, J.I. PAX8 positivity in nested variant of urothelial carcinoma: A potential diagnostic pitfall. Hum. Pathol. 2019, 94, 11–15. [Google Scholar] [CrossRef]
- Taylor, A.S.; McKenney, J.K.; Osunkoya, A.O.; Chan, M.P.; Al-Ahmadie, H.A.; Spratt, D.E.; Fullen, D.R.; Chinnaiyan, A.M.; Brown, N.A.; Mehra, R. PAX8 expression and TERT promoter mutations in the nested variant of urothelial carcinoma: A clinicopathologic study with immunohistochemical and molecular correlates. Mod. Pathol. 2020, 33, 1165–1171. [Google Scholar] [CrossRef]
- Pellizzari, L.; Puppin, C.; Mariuzzi, L.; Saro, F.; Pandolfi, M.; Di Lauro, R.; Beltrami, C.A.; Damante, G. PAX8 expression in human bladder cancer. Oncol. Rep. 2006, 16, 1015–1020. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Cao, J.; Li, J.; Yao, Z.; Han, D.; Ying, L.; Wang, Z.; Tian, J. Identification of prognostic biomarkers associated with stromal cell infiltration in muscle-invasive bladder cancer by bioinformatics analyses. Cancer Med. 2020, 9, 7253–7267. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Tang, Y.; Ren, H.; Lei, Y. Identification of Prognosis-Related Genes in Bladder Cancer Microenvironment across TCGA Database. BioMed Res. Int. 2020, 2020, 9143695. [Google Scholar] [CrossRef]
- Chen, C.; Luo, Y.; He, W.; Zhao, Y.; Kong, Y.; Liu, H.; Zhong, G.; Li, Y.; Li, J.; Huang, J.; et al. Exosomal long noncoding RNA LNMAT2 promotes lymphatic metastasis in bladder cancer. J. Clin. Investig. 2019, 130, 404–421. [Google Scholar] [CrossRef] [PubMed]
- Na, X.Y.; Shang, X.S.; Zhao, Y.; Ren, P.P.; Hu, X.Q. MiR-203a functions as a tumor suppressor in bladder cancer by targeting SIX4. Neoplasma 2019, 66, 211–221. [Google Scholar] [CrossRef]
- Huang, H.-S.; Liu, Z.-M.; Chen, P.-C.; Tseng, H.-Y.; Yeh, B.-W. TG-interacting factor-induced superoxide production from NADPH oxidase contributes to the migration/invasion of urothelial carcinoma. Free Radic. Biol. Med. 2012, 53, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Yeh, B.-W.; Li, W.-M.; Li, C.-C.; Kang, W.-Y.; Huang, C.-N.; Hour, T.-C.; Liu, Z.-M.; Wu, W.-J.; Huang, H.-S. Histone deacetylase inhibitor trichostatin A resensitizes gemcitabine resistant urothelial carcinoma cells via suppression of TG-interacting factor. Toxicol. Appl. Pharmacol. 2016, 290, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Zhu, S.; Zang, J.; Wu, G.; Wen, Y.; Liang, Y.; Long, Y.; Guo, W.; Zang, C.; Hu, X.; et al. ADNP prompts the cisplatin-resistance of bladder cancer via TGF-β-mediated epithelial-mesenchymal transition (EMT) pathway. J. Cancer 2021, 12, 5114–5124. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Xu, Z.; Zeng, Y.; Long, Y.; Fan, G.; Ding, Q.; Wen, Y.; Cao, J.; Dai, T.; Han, W.; et al. ADNP Upregulation Promotes Bladder Cancer Cell Proliferation via the AKT Pathway. Front. Oncol. 2020, 10, 491129. [Google Scholar] [CrossRef]
- Mahdavinezhad, A.; Yadegarazari, R.; Mousavi-Bahar, S.H.; Poorolajal, J.; Jafari, M.; Amirzargar, M.A.; Effatpanah, H.; Saidijam, M. Evaluation of zinc finger E-box binding homeobox 1 and transforming growth factor-beta2 expression in bladder cancer tissue in comparison with healthy adjacent tissue. Investig. Clin. Urol. 2017, 58, 140–145. [Google Scholar] [CrossRef]
- Liang, Y.; Ye, F.; Xu, C.; Zou, L.; Hu, Y.; Hu, J.; Jiang, H. A novel survival model based on a ferroptosis-related gene signature for predicting overall survival in bladder cancer. BMC Cancer 2021, 21, 943. [Google Scholar] [CrossRef]
- Li, B.; Mao, X.; Wang, H.; Su, G.; Mo, C.; Cao, K.; Qiu, S. Vasculogenic mimicry in bladder cancer and its association with the aberrant expression of ZEB1. Oncol. Lett. 2018, 15, 5193–5200. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhang, C.; Wu, Y.; He, W.; Gou, X. Identification and analysis of long non-coding RNA related miRNA sponge regulatory network in bladder urothelial carcinoma. Cancer Cell Int. 2019, 19, 327. [Google Scholar] [CrossRef]
- Sun, J.; Yue, W.; You, J.; Wei, X.; Huang, Y.; Ling, Z.; Hou, J. Identification of a Novel Ferroptosis-Related Gene Prognostic Signature in Bladder Cancer. Front. Oncol. 2021, 11, 730716. [Google Scholar] [CrossRef]
- Yi, K.; Liu, J.; Rong, Y.; Wang, C.; Tang, X.; Zhang, X.; Xiong, Y.; Wang, F. Biological Functions and Prognostic Value of Ferroptosis-Related Genes in Bladder Cancer. Front. Mol. Biosci. 2021, 8, 631152. [Google Scholar] [CrossRef] [PubMed]
- Matsui, Y.; Assi, K.; Ogawa, O.; Raven, P.A.; Dedhar, S.; Gleave, M.E.; Salh, B.; So, A.I. The importance of integrin-linked kinase in the regulation of bladder cancer invasion. Int. J. Cancer 2011, 130, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Genitsch, V.; Kollár, A.; Vandekerkhove, G.; Blarer, J.; Furrer, M.; Annala, M.; Herberts, C.; Pycha, A.; de Jong, J.J.; Liu, Y.; et al. Morphologic and genomic characterization of urothelial to sarcomatoid transition in muscle-invasive bladder cancer. Urol. Oncol. Semin. Orig. Investig. 2019, 37, 826–836. [Google Scholar] [CrossRef]
- Morelli, M.B.; Amantini, C.; de Vermandois, J.A.R.; Gubbiotti, M.; Giannantoni, A.; Mearini, E.; Maggi, F.; Nabissi, M.; Marinelli, O.; Santoni, M.; et al. Correlation between High PD-L1 and EMT/Invasive Genes Expression and Reduced Recurrence-Free Survival in Blood-Circulating Tumor Cells from Patients with Non-Muscle-Invasive Bladder Cancer. Cancers 2021, 13, 5989. [Google Scholar] [CrossRef]
- Wu, K.; Fan, J.; Zhang, L.; Ning, Z.; Zeng, J.; Zhou, J.; Li, L.; Chen, Y.; Zhang, T.; Wang, X.; et al. PI3K/Akt to GSK3β/β-catenin signaling cascade coordinates cell colonization for bladder cancer bone metastasis through regulating ZEB1 transcription. Cell Signal. 2012, 24, 2273–2282. [Google Scholar] [CrossRef]
- Shen, J.; Lv, X.; Zhang, L. GRHL2 Acts as an Anti-Oncogene in Bladder Cancer by Regulating ZEB1 in Epithelial-Mesenchymal Transition (EMT) Process. OncoTargets Ther. 2020, 13, 2511–2522. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Huang, Z.; Zhang, M.; Wang, W.; Liang, H.; Zeng, J.; Wu, K.; Wang, X.; Hsieh, J.; Guo, P.; et al. HIF-1α promotes ZEB1 expression and EMT in a human bladder cancer lung metastasis animal model. Oncol. Lett. 2018, 15, 3482–3489. [Google Scholar] [CrossRef]
- Majid, S.; Dar, A.A.; Saini, S.; Deng, G.; Chang, I.; Greene, K.; Tanaka, Y.; Dahiya, R.; Yamamura, S. MicroRNA-23b Functions as a Tumor Suppressor by Regulating Zeb1 in Bladder Cancer. PLoS ONE 2013, 8, e67686. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-L.; Ho, J.-Y.; Chou, S.-C.; Yu, D.-S. MiR-429 reverses epithelial-mesenchymal transition by restoring E-cadherin expression in bladder cancer. Oncotarget 2016, 7, 26593–26603. [Google Scholar] [CrossRef]
- Zhi, Y.; Pan, J.; Shen, W.; He, P.; Zheng, J.; Zhou, X.; Lu, G.; Chen, Z.; Zhou, Z. Ginkgolide B Inhibits Human Bladder Cancer Cell Migration and Invasion Through MicroRNA-223-3p. Cell Physiol. Biochem. 2016, 39, 1787–1794. [Google Scholar] [CrossRef]
- Xie, X.; Rui, W.; He, W.; Shao, Y.; Sun, F.; Zhou, W.; Wu, Y.; Zhu, Y. Discoidin domain receptor 1 activity drives an aggressive phenotype in bladder cancer. Am. J. Transl. Res. 2017, 9, 2500–2507. [Google Scholar]
- Wu, K.; Ning, Z.; Zeng, J.; Fan, J.; Zhou, J.; Zhang, T.; Zhang, L.; Chen, Y.; Gao, Y.; Wang, B.; et al. Silibinin inhibits β-catenin/ZEB1 signaling and suppresses bladder cancer metastasis via dual-blocking epithelial–mesenchymal transition and stemness. Cell Signal. 2013, 25, 2625–2633. [Google Scholar] [CrossRef]
- Shan, Y.; Zhang, L.; Bao, Y.; Li, B.; He, C.; Gao, M.; Feng, X.; Xu, W.; Zhang, X.; Wang, S. Epithelial-mesenchymal transition, a novel target of sulforaphane via COX-2/MMP2, 9/Snail, ZEB1 and miR-200c/ZEB1 pathways in human bladder cancer cells. J. Nutr. Biochem. 2013, 24, 1062–1069. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, D.; Ding, Y.; Zhou, J.; Liu, G.; Ji, Z. lncRNA ZEB1-AS1 promotes migration and metastasis of bladder cancer cells by post-transcriptional activation of ZEB1. Int. J. Mol. Med. 2019, 44, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.-K.; Wang, J.-M.; Zhang, P.; Wang, Y.-Q. MicroRNA-138 Regulates Metastatic Potential of Bladder Cancer Through ZEB2. Cell Physiol. Biochem. 2015, 37, 2366–2374. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, G.; Zheng, W.; Xue, Q.; Wei, D.; Zheng, Y.; Yuan, J. MiR-454-3p and miR-374b-5p suppress migration and invasion of bladder cancer cells through targetting ZEB2. Biosci. Rep. 2018, 38, BSR20181436. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Qiu, K.; Li, M.; Liang, Y. Double-negative feedback loop between long non-coding RNA TUG1 and miR-145 promotes epithelial to mesenchymal transition and radioresistance in human bladder cancer cells. FEBS Lett. 2015, 589, 3175–3181. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liu, H.; Cheng, H.; Li, Y.; Li, X.; Zhu, C. Downregulation of long noncoding RNA TUG1 inhibits proliferation and induces apoptosis through the TUG1/miR-142/ZEB2 axis in bladder cancer cells. OncoTargets Ther. 2017, 10, 2461–2471. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.-Y.; Liu, Q.-H.; Wang, T.-J.; Zhao, J.; Cheng, X.-H.; Wang, J.-S. CircZFR serves as a prognostic marker to promote bladder cancer progression by regulating miR-377/ZEB2 signaling. Biosci. Rep. 2019, 39, BSR20192779. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yan, T.; Wang, Z.; Wu, X.; Cao, G.; Zhang, C. LncRNA ZEB2-AS1 promotes bladder cancer cell proliferation and inhibits apoptosis by regulating miR-27b. Biomed. Pharmacother. 2017, 96, 299–304. [Google Scholar] [CrossRef]
- Kenney, P.A.; Wszolek, M.F.; Rieger-Christ, K.M.; Neto, B.S.; Gould, J.J.; Harty, N.J.; Mosquera, J.M.; Zeheb, R.; Loda, M.; Darling, D.S.; et al. Novel ZEB1 expression in bladder tumorigenesis. BJU Int. 2011, 107, 656–663. [Google Scholar] [CrossRef]
- Sayan, A.E.; Griffiths, T.R.; Pal, R.; Browne, G.J.; Ruddick, A.; Yagci, T.; Edwards, R.; Mayer, N.J.; Qazi, H.; Goyal, S.; et al. SIP1 protein protects cells from DNA damage-induced apoptosis and has independent prognostic value in bladder cancer. Proc. Natl. Acad. Sci. USA 2009, 106, 14884–14889. [Google Scholar] [CrossRef]
- Choi, W.; Shah, J.B.; Tran, M.; Svatek, R.; Marquis, L.; Lee, I.-L.; Yu, D.; Adam, L.; Wen, S.; Shen, Y.; et al. p63 Expression Defines a Lethal Subset of Muscle-Invasive Bladder Cancers. PLoS ONE 2012, 7, e30206. [Google Scholar] [CrossRef] [Green Version]
- Adam, L.; Zhong, M.; Choi, W.; Qi, W.; Nicoloso, M.; Arora, A.; Calin, G.; Wang, H.; Siefker-Radtke, A.; McConkey, D.; et al. miR-200 Expression Regulates Epithelial-to-Mesenchymal Transition in Bladder Cancer Cells and Reverses Resistance to Epidermal Growth Factor Receptor Therapy. Clin. Cancer Res. 2009, 15, 5060–5072. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Fernández, M.; Dueñas, M.; Feber, A.; Segovia, C.; García-Escudero, R.; Rubio, C.; López-Calderón, F.F.; Díaz-García, C.; Villacampa, F.; Duarte, J.; et al. A Polycomb-mir200 loop regulates clinical outcome in bladder cancer. Oncotarget 2015, 6, 42258–42275. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Jiang, T.; Mao, H.; Gao, R.; Zhang, H.; He, Y.; Liu, C.; Chen, Q. SNHG16 regulates invasion and migration of bladder cancer through induction of epithelial-to-mesenchymal transition. Hum. Cell 2020, 33, 737–749. [Google Scholar] [CrossRef]
- Lee, H.; Jun, S.-Y.; Lee, Y.-S.; Lee, H.J.; Lee, W.S.; Park, C.S. Expression of miRNAs and ZEB1 and ZEB2 correlates with histopathological grade in papillary urothelial tumors of the urinary bladder. Virchows Arch. 2014, 464, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, G.; Cheng, B.; Jiang, R. ZFAS1 functions as an oncogenic long non-coding RNA in bladder cancer. Biosci. Rep. 2018, 38, BSR20180475. [Google Scholar] [CrossRef] [Green Version]
- Dong, B.; Liang, J.; Li, D.; Song, W.; Song, J.; Zhu, M.; Zhao, S.; Ma, Y.; Yang, T. Identification of a Prognostic Signature Associated With the Homeobox Gene Family for Bladder Cancer. Front. Mol. Biosci. 2021, 8, 688298. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Wei, W.; Duan, J.; Chen, R.; Wang, N.; He, L.; Peng, Y.; Ma, X.; Wu, Z.; Liu, J.; et al. ZHX3 promotes the progression of urothelial carcinoma of the bladder via repressing of RGS2 and is a novel substrate of TRIM21. Cancer Sci. 2021, 112, 1758–1771. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chin, F.-W.; Chan, S.-C.; Veerakumarasivam, A. Homeobox Gene Expression Dysregulation as Potential Diagnostic and Prognostic Biomarkers in Bladder Cancer. Diagnostics 2023, 13, 2641. https://doi.org/10.3390/diagnostics13162641
Chin F-W, Chan S-C, Veerakumarasivam A. Homeobox Gene Expression Dysregulation as Potential Diagnostic and Prognostic Biomarkers in Bladder Cancer. Diagnostics. 2023; 13(16):2641. https://doi.org/10.3390/diagnostics13162641
Chicago/Turabian StyleChin, Fee-Wai, Soon-Choy Chan, and Abhi Veerakumarasivam. 2023. "Homeobox Gene Expression Dysregulation as Potential Diagnostic and Prognostic Biomarkers in Bladder Cancer" Diagnostics 13, no. 16: 2641. https://doi.org/10.3390/diagnostics13162641
APA StyleChin, F. -W., Chan, S. -C., & Veerakumarasivam, A. (2023). Homeobox Gene Expression Dysregulation as Potential Diagnostic and Prognostic Biomarkers in Bladder Cancer. Diagnostics, 13(16), 2641. https://doi.org/10.3390/diagnostics13162641