

Discovering Breast Cancer Biomarkers Candidates through mRNA Expression Analysis Based on The Cancer Genome Atlas Database


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Public Data Collection
2.2. mRNA Expression Analysis
2.3. Diagnostic Performance Analysis
2.4. Statistical Analysis
3. Results
3.1. Characteristics of Breast Cancer Patients
3.2. Differential Expression of Breast Cancer mRNA Profiles
3.3. Diagnostic Performance Analysis via mRNA Expression Profile Verification
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, W.; Li, X.; Wang, W.; Chen, B.; Wang, L.; Zhang, N.; Wang, Z.; Yang, Q. Association of Preoperative Serum Levels of CEA and CA15-3 with Molecular Subtypes of Breast Cancer. Dis. Markers 2021, 2021, 5529106. [Google Scholar] [CrossRef] [PubMed]
- O’Hanlon, D.M.; Kerin, M.J.; Kent, P.; Maher, D.; Grimes, H.; Given, H.F. An evaluation of preoperative CA 15-3 measurement in primary breast carcinoma. Br. J. Cancer 1995, 71, 1288–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uehara, M.; Kinoshita, T.; Hojo, T.; Akashi-Tanaka, S.; Iwamoto, E.; Fukutomi, T. Long-term prognostic study of carcinoembryonic antigen (CEA) and carbohydrate antigen 15-3 (CA 15-3) in breast cancer. Int. J. Clin. Oncol. 2008, 13, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Hashim, Z.M. The significance of CA15-3 in breast cancer patients and its relationship to HER-2 receptor status. Int. J. Immunopathol. Pharmacol. 2014, 27, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Hammarstrom, S. The carcinoembryonic antigen (CEA) family: Structures, suggested functions and expression in normal and malignant tissues. Semin. Cancer Biol. 1999, 9, 67–81. [Google Scholar] [CrossRef]
- Molina, R.; Barak, V.; van Dalen, A.; Duffy, M.J.; Einarsson, R.; Gion, M.; Goike, H.; Lamerz, R.; Nap, M.; Soletormos, G.; et al. Tumor markers in breast cancer—European Group on Tumor Markers recommendations. Tumour Biol. 2005, 26, 281–293. [Google Scholar] [CrossRef] [Green Version]
- Gradishar, W.J.; Moran, M.S.; Abraham, J.; Aft, R.; Agnese, D.; Allison, K.H.; Anderson, B.; Burstein, H.J.; Chew, H.; Dang, C.; et al. Breast Cancer, Version 3.2022, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Canc. Netw. 2022, 20, 691–722. [Google Scholar] [CrossRef]
- Harris, L.; Fritsche, H.; Mennel, R.; Norton, L.; Ravdin, P.; Taube, S.; Somerfield, M.R.; Hayes, D.F.; Bast, R.C., Jr.; American Society of Clinical, O. American Society of Clinical Oncology 2007 update of recommendations for the use of tumor markers in breast cancer. J. Clin. Oncol. 2007, 25, 5287–5312. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Lichtenberg, T.; Hoadley, K.A.; Poisson, L.M.; Lazar, A.J.; Cherniack, A.D.; Kovatich, A.J.; Benz, C.C.; Levine, D.A.; Lee, A.V.; et al. An Integrated TCGA Pan-Cancer Clinical Data Resource to Drive High-Quality Survival Outcome Analytics. Cell 2018, 173, 400–416. [Google Scholar] [CrossRef] [Green Version]
- Gao, G.F.; Parker, J.S.; Reynolds, S.M.; Silva, T.C.; Wang, L.B.; Zhou, W.; Akbani, R.; Bailey, M.; Balu, S.; Berman, B.P.; et al. Before and After: Comparison of Legacy and Harmonized TCGA Genomic Data Commons’ Data. Cell Syst. 2019, 9, 24–34. [Google Scholar] [CrossRef]
- The ICGC/TCGA Pan-Cancer Analysis of Whole Genomes Consortium. Pan-cancer analysis of whole genomes. Nature 2020, 578, 82–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cancer Genome Atlas Research Network; Albert Einstein College of Medicine; Analytical Biological Services; Barretos Cancer Hospital; Baylor College of Medicine; Beckman Research Institute of City of Hope; Buck Institute for Research on Aging; Canada’s Michael Smith Genome Sciences Centre; Harvard Medical School; Helen F. Graham Cancer Center &Research Institute at Christiana Care Health Services; et al. Integrated genomic and molecular characterization of cervical cancer. Nature 2017, 543, 378–384. [Google Scholar] [CrossRef] [Green Version]
- Ai, D.; Yao, J.; Yang, F.; Huo, L.; Chen, H.; Lu, W.; Soto, L.M.S.; Jiang, M.; Raso, M.G.; Wang, S.; et al. TRPS1: A highly sensitive and specific marker for breast carcinoma, especially for triple-negative breast cancer. Mod. Pathol. 2021, 34, 710–719. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wu, Y.; Liu, Y.; Pan, F.; Zeng, H.; Li, X.; Yu, L. Tumor Suppressor Effect of RBMS3 in Breast Cancer. Technol. Cancer Res. Treat. 2021, 20, 15330338211004921. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Tang, T.; Zhang, G.; Liang, T. Identification of tumor antigens and immune subtypes of cholangiocarcinoma for mRNA vaccine development. Mol. Cancer 2021, 20, 50. [Google Scholar] [CrossRef]
- Liang, W.; Mo, C.; Wei, J.; Chen, W.; Gong, W.; Shi, J.; Hou, X.; Li, C.; Deng, Y.; Ou, M. FAM65A as a novel prognostic biomarker in human tumors reveal by a pan-cancer analysis. Discov. Oncol. 2021, 12, 60. [Google Scholar] [CrossRef]
- GTEx Consortium; Laboratory, Data Analysis & Coordinating Center (LDACC)—Analysis Working Group; Statistical Methods groups—Analysis Working Group; Enhancing GTEx (eGTEx) groups; NIH Common Fund; NIH/NCI; NIH/NHGRI; NIH/NIMH; NIH/NIDA; Biospecimen Collection Source Site—NDRI; et al. Genetic effects on gene expression across human tissues. Nature 2017, 550, 204–213. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Liu, L.; Feng, Z.; Wang, X.; Huang, Y.; Dai, H.; Zhang, L.; Song, F.; Wang, D.; Zhang, P.; et al. Tumor markers CA15-3, CA125, CEA and breast cancer survival by molecular subtype: A cohort study. Breast Cancer 2020, 27, 621–630. [Google Scholar] [CrossRef]
- Fan, Y.; Ren, X.; Liu, X.; Shi, D.; Xu, E.; Wang, S.; Liu, Y. Combined detection of CA15-3, CEA, and SF in serum and tissue of canine mammary gland tumor patients. Sci. Rep. 2021, 11, 6651. [Google Scholar] [CrossRef]
- Nam, S.E.; Lim, W.; Jeong, J.; Lee, S.; Choi, J.; Park, H.; Jung, Y.S.; Jung, S.P.; Bae, S.Y. The prognostic significance of preoperative tumor marker (CEA, CA15-3) elevation in breast cancer patients: Data from the Korean Breast Cancer Society Registry. Breast Cancer Res. Treat. 2019, 177, 669–678. [Google Scholar] [CrossRef]
- Greiner, M.; Pfeiffer, D.; Smith, R.D. Principles and practical application of the receiver-operating characteristic analysis for diagnostic tests. Prev. Vet. Med. 2000, 45, 23–41. [Google Scholar] [CrossRef]
- Yan, S.; Deng, Y.; Qiang, Y.; Xi, Q.; Liu, R.; Yang, S.; Liu, J.; Tang, C.; Zhong, J.; Wang, Y. SYF2 is upregulated in human epithelial ovarian cancer and promotes cell proliferation. Tumour Biol. 2015, 36, 4633–4642. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Cai, F.F.; Cai, L.; Lin, X.Y.; Zhang, W.; Wang, Q.Q.; Zhao, Y.J.; Ni, Q.C.; Wang, H.; He, Z.X. Overexpression of SYF2 promotes cell proliferation and correlates with poor prognosis in human breast cancer. Oncotarget 2017, 8, 88453–88463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.; Zhang, Z.; Chen, H.; Cai, R.; Yang, Q.; Liu, Q.; Fan, Y.; Liu, W.; Yao, C. Carboxypeptidase N2 as a Novel Diagnostic and Prognostic Biomarker for Lung Adenocarcinoma. Front. Oncol. 2022, 12, 843325. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Y.; Chen, T.; Kuklina, A.S.; Bernard, P.; Esteva, F.J.; Shen, H.; Ferrari, M.; Hu, Y. Circulating proteolytic products of carboxypeptidase N for early detection of breast cancer. Clin. Chem. 2014, 60, 233–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, J.; Tanikawa, C.; Ohnishi, N.; Funauchi, Y.; Toyoshima, O.; Ueda, K.; Matsuda, K. EPSIN 3, A Novel p53 Target, Regulates the Apoptotic Pathway and Gastric Carcinogenesis. Neoplasia 2017, 19, 185–195. [Google Scholar] [CrossRef]
- Wang, Y.; Song, W.; Kan, P.; Huang, C.; Ma, Z.; Wu, Q.; Yao, X.; Zhang, B. Overexpression of Epsin 3 enhances migration and invasion of glioma cells by inducing epithelialmesenchymal transition. Oncol. Rep. 2018, 40, 3049–3059. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Li, Q.; Zhu, W.; Zhang, X.; Li, H. Epsin 3 potentiates the NFkappaB signaling pathway to regulate apoptosis in breast cancer. Mol. Med. Rep. 2022, 25, 15. [Google Scholar] [CrossRef]
- Roversi, F.M.; Olalla Saad, S.T.; Machado-Neto, J.A. Serine peptidase inhibitor Kunitz type 2 (SPINT2) in cancer development and progression. Biomed. Pharmacother. 2018, 101, 278–286. [Google Scholar] [CrossRef]
- Wang, N.; Che, Y.; Yin, F.; Yu, F.; Bi, X.; Wang, Y. Study on the methylation status of SPINT2 gene and its expression in cervical carcinoma. Cancer Biomark. 2018, 22, 435–442. [Google Scholar] [CrossRef]
- Pereira, M.S.; de Almeida, G.C.; Pinto, F.; Viana-Pereira, M.; Reis, R.M. SPINT2 Deregulation in Prostate Carcinoma. J. Histochem. Cytochem. 2016, 64, 32–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Yin, G.; Luo, J.; Zhang, Y.; Ai, T.; Tian, J.; Jin, Y.; Lei, J.; Liu, S. Comprehensive Analysis of the Expression and Prognostic Value of SPINT1/2 in Breast Carcinoma. Front. Endocrinol. 2021, 12, 665666. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Chen, C.; Luo, B.; Yan, D.; Yan, H.; Chen, F.; Guan, F.; Wu, H.; Yuan, J. Nuclear ING3 Expression Is Correlated with a Good Prognosis of Breast Cancer. Front. Oncol. 2020, 10, 589009. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, H.; Tan, X.; Liu, D.; Guo, R.; Wang, M.; Tang, Y.; Zheng, K.; Chen, W.; Li, H.; et al. Overexpression of ING3 is associated with attenuation of migration and invasion in breast cancer. Exp. Ther. Med. 2021, 22, 699. [Google Scholar] [CrossRef]
- Lu, M.; Chen, F.; Wang, Q.; Wang, K.; Pan, Q.; Zhang, X. Downregulation of inhibitor of growth 3 is correlated with tumorigenesis and progression of hepatocellular carcinoma. Oncol. Lett. 2012, 4, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Gunduz, M.; Beder, L.B.; Gunduz, E.; Nagatsuka, H.; Fukushima, K.; Pehlivan, D.; Cetin, E.; Yamanaka, N.; Nishizaki, K.; Shimizu, K.; et al. Downregulation of ING3 mRNA expression predicts poor prognosis in head and neck cancer. Cancer Sci. 2008, 99, 531–538. [Google Scholar] [CrossRef]
- Gou, W.F.; Sun, H.Z.; Zhao, S.; Niu, Z.F.; Mao, X.Y.; Takano, Y.; Zheng, H.C. Downregulated inhibitor of growth 3 (ING3) expression during colorectal carcinogenesis. Indian J. Med. Res. 2014, 139, 561–567. [Google Scholar]
- Lu, Z.; Mao, W.; Yang, H.; Santiago-O’Farrill, J.M.; Rask, P.J.; Mondal, J.; Chen, H.; Ivan, C.; Liu, X.; Liu, C.G.; et al. SIK2 inhibition enhances PARP inhibitor activity synergistically in ovarian and triple-negative breast cancers. J. Clin. Investig. 2022, 132, e146471. [Google Scholar] [CrossRef]
- Maxfield, K.E.; Macion, J.; Vankayalapati, H.; Whitehurst, A.W. SIK2 Restricts Autophagic Flux to Support Triple-Negative Breast Cancer Survival. Mol. Cell Biol. 2016, 36, 3048–3057. [Google Scholar] [CrossRef] [Green Version]
- Xin, L.; Liu, C.; Liu, Y.; Mansel, R.E.; Ruge, F.; Davies, E.; Jiang, W.G.; Martin, T.A. SIKs suppress tumor function and regulate drug resistance in breast cancer. Am. J. Cancer Res. 2021, 11, 3537–3557. [Google Scholar]
- Dai, X.M.; Zhang, Y.H.; Lin, X.H.; Huang, X.X.; Zhang, Y.; Xue, C.R.; Chen, W.N.; Ye, J.X.; Lin, X.J.; Lin, X. SIK2 represses AKT/GSK3beta/beta-catenin signaling and suppresses gastric cancer by inhibiting autophagic degradation of protein phosphatases. Mol. Oncol. 2021, 15, 228–245. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.; Liu, S.; Wang, C.; Gu, X.; Linghu, E.; Xue, X. Mass spectrum analysis of membrane proteins reveals that CASK, CD36 and EPB42 are differentially expressed in pancreatic adenocarcinoma. Oncol. Lett. 2020, 20, 376. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Jin, J.; Chen, J.; Lou, W. MUC14-Related ncRNA-mRNA Network in Breast Cancer. Genes 2021, 12, 1677. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, X.; Xu, L.; Zhang, J. PBX1, EMCN and ERG are associated with the sub-clusters and the prognosis of VHL mutant clear cell renal cell carcinoma. Sci. Rep. 2022, 12, 8955. [Google Scholar] [CrossRef]
- Jiang, H.; Chen, H.; Wan, P.; Chen, N. Decreased expression of HADH is related to poor prognosis and immune infiltration in kidney renal clear cell carcinoma. Genomics 2021, 113, 3556–3564. [Google Scholar] [CrossRef]
- Shen, C.; Song, Y.H.; Xie, Y.; Wang, X.; Wang, Y.; Wang, C.; Liu, S.; Xue, S.L.; Li, Y.; Liu, B.; et al. Downregulation of HADH promotes gastric cancer progression via Akt signaling pathway. Oncotarget 2017, 8, 76279–76289. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.; Zhang, X.; Gao, W.; Yang, J. Differentially expressed genes PCCA, ECHS1, and HADH are potential prognostic biomarkers for gastric cancer. Sci. Prog. 2021, 104, 368504211011344. [Google Scholar] [CrossRef]
- Yang, P.J.; Hou, M.F.; Tsai, E.M.; Liang, S.S.; Chiu, C.C.; Ou-Yang, F.; Kan, J.Y.; Peng, C.Y.; Wang, T.N. Breast cancer is associated with methylation and expression of the a disintegrin and metalloproteinase domain 33 (ADAM33) gene affected by endocrinedisrupting chemicals. Oncol. Rep. 2018, 40, 2766–2777. [Google Scholar] [CrossRef]
- Seniski, G.G.; Camargo, A.A.; Ierardi, D.F.; Ramos, E.A.; Grochoski, M.; Ribeiro, E.S.; Cavalli, I.J.; Pedrosa, F.O.; de Souza, E.M.; Zanata, S.M.; et al. ADAM33 gene silencing by promoter hypermethylation as a molecular marker in breast invasive lobular carcinoma. BMC Cancer 2009, 9, 80. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Yue, Y.; Pan, M.; Sun, J.; Chu, J.; Lin, X.; Xu, W.; Feng, L.; Chen, Y.; Chen, D.; et al. Histone deacetylase 3 inhibits new tumor suppressor gene DTWD1 in gastric cancer. Am. J. Cancer Res. 2015, 5, 663–673. [Google Scholar]
- Jin, Y.; Yang, Y. Bioinformatics-based discovery of PYGM and TNNC2 as potential biomarkers of head and neck squamous cell carcinoma. Biosci. Rep. 2019, 39, BSR20191612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Almeida, B.P.; Apolonio, J.D.; Binnie, A.; Castelo-Branco, P. Roadmap of DNA methylation in breast cancer identifies novel prognostic biomarkers. BMC Cancer 2019, 19, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coon, B.G.; Burgner, J.; Camonis, J.H.; Aguilar, R.C. The epsin family of endocytic adaptors promotes fibrosarcoma migration and invasion. J. Biol. Chem. 2010, 285, 33073–33081. [Google Scholar] [CrossRef] [PubMed]
- Rakha, E.A.; Reis-Filho, J.S.; Baehner, F.; Dabbs, D.J.; Decker, T.; Eusebi, V.; Fox, S.B.; Ichihara, S.; Jacquemier, J.; Lakhani, S.R.; et al. Breast cancer prognostic classification in the molecular era: The role of histological grade. Breast Cancer Res. 2010, 12, 207. [Google Scholar] [CrossRef] [Green Version]
- Harbeck, N.; Gnant, M. Breast cancer. Lancet 2017, 389, 1134–1150. [Google Scholar] [CrossRef]
- Yersal, O.; Barutca, S. Biological subtypes of breast cancer: Prognostic and therapeutic implications. World J. Clin. Oncol. 2014, 5, 412–424. [Google Scholar] [CrossRef]
- Loibl, S.; Poortmans, P.; Morrow, M.; Denkert, C.; Curigliano, G. Breast cancer. Lancet 2021, 397, 1750–1769. [Google Scholar] [CrossRef]
- Kumar, P.; Aggarwal, R. An overview of triple-negative breast cancer. Arch. Gynecol. Obstet. 2016, 293, 247–269. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Breast Cancer, n = 526 n, (%) |
|---|---|
| Female | 520, (98.86) |
| Age (y, mean ± SD) | 57.91, ± 13.26 |
| Race | |
| Asian | 34, (6.46) |
| African American | 40, (7.60) |
| Caucasian | 361, (68.63) |
| American Indian or Alaska native | 1, (0.19) |
| Unknown | 90, (17.11) |
| Tumor stage | |
| Stage I | 89, (16.92) |
| Stage II | 296, (56.27) |
| Stage III | 110, (20.91) |
| Stage IV | 13, (2.47) |
| Unknown | 18, (3.43) |
| Molecular subtype | |
| Luminal A | 330, (62.74) |
| Luminal B | 79, (15.02) |
| HER2-positive | 25, (4.75) |
| TNBC | 92, (17.49) |
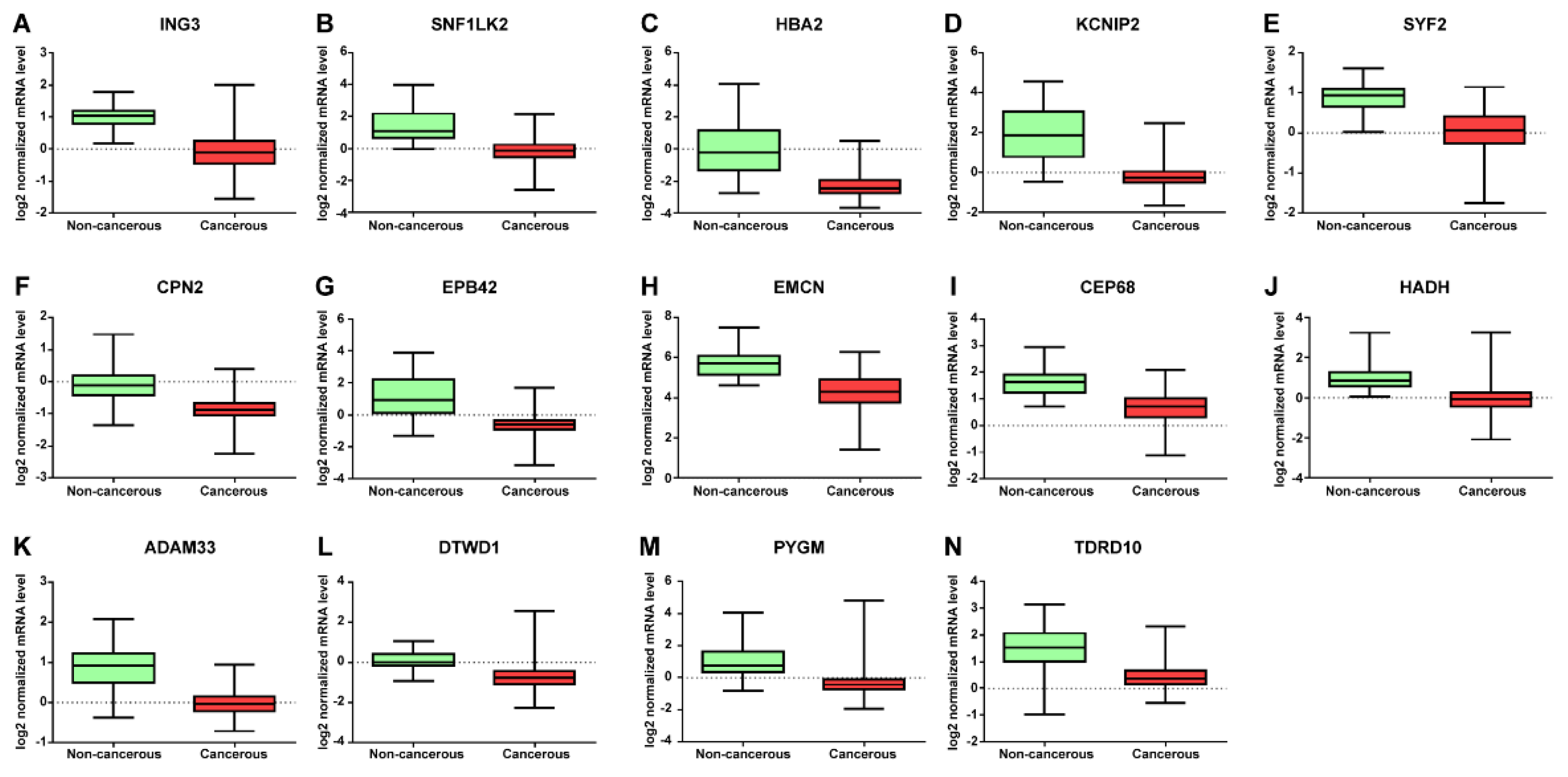
| mRNAs | Log2 Normalized mRNA Level | AUC | Cutoff | Sensitivity | Specificity | p-Value | |
|---|---|---|---|---|---|---|---|
| Non-Cancerous | Cancerous | ||||||
| ING3 | 0.9884 ± 0.03998 | −0.09470 ± 0.2354 | 0.96 (0.94–0.97) | <0.5343 | 87.71% (84.61–90.39) | 91.67% (81.61–97.24) | <0.0001 |
| SNF1LK2 | 1.426 ± 0.1338 | −0.1613 ± 0.02701 | 0.94 (0.92–0.97) | <0.4393 | 85.82% (82.56–88.68) | 88.33% (77.43–95.18) | <0.0001 |
| EVPL | −0.02857 ± 0.08834 | 0.8723 ± 0.01995 | 0.94 (0.92–0.97) | >0.4566 | 84.12% (80.72–87.13) | 93.33% (83.80–98.15) | <0.0001 |
| HBA2 | 0.06397 ± 0.2116 | −2.279 ± 0.02886 | 0.93 (0.90–0.97) | <−1.616 | 85.07% (81.74–88.00) | 86.67% (75.41–94.06) | <0.0001 |
| KCNIP2 | 1.955 ± 0.1823 | −0.1219 ± 0.02623 | 0.93 (0.90–0.96) | <0.3937 | 85.82% (82.56–88.68) | 86.67% (75.41–94.06) | <0.0001 |
| SYF2 | 0.8908 ± 0.07179 | 0.06124 ± 0.02065 | 0.93 (0.90–0.96) | <0.5191 | 82.99% (79.51–86.09) | 90.00% (79.49–96.24) | <0.0001 |
| AP1M2 | −0.04386 ± 0.1699 | 1.649 ± 0.02720 | 0.93 (0.90–0.96) | >1.111 | 86.39% (83.17–89.20) | 86.67% (75.41–94.06) | <0.0001 |
| CPN2 | −0.08433 ± 0.08377 | −0.8673 ± 0.01309 | 0.93 (0.89–0.97) | <−0.5644 | 85.44% (82.15–88.34) | 88.33% (77.43–95.18) | <0.0001 |
| EPB42 | 1.208 ± 0.1597 | −0.5775 ± 0.02256 | 0.93 (0.89–0.97) | <−0.2037 | 82.99% (79.51–86.09) | 86.67% (75.41–94.06) | <0.0001 |
| TOMM40 | −1.474 ± 0.07094 | −0.6728 ± 0.01989 | 0.92 (0.89–0.96) | >−1.093 | 82.04% (78.50–85.22) | 86.67% (75.41–94.06) | <0.0001 |
| EMCN | 5.715 ± 0.1217 | 4.270 ± 0.03668 | 0.93 (0.90–0.95) | <5.048 | 81.66% (78.10–84.87) | 86.67% (75.41–94.06) | <0.0001 |
| CEP68 | 1.610 ± 0.08263 | 0.6641 ± 0.02243 | 0.92 (0.89–0.95) | <1.113 | 81.66% (78.10–84.87) | 83.33% (71.48–91.71) | <0.0001 |
| HADH | 1.056 ± 0.1059 | −0.08562 ± 0.02584 | 0.92 (0.87–0.95) | <0.4448 | 83.74% (80.32–86.79) | 90.00% (79.49–96.24) | <0.0001 |
| ADAM33 | 0.8487 ± 0.08717 | −0.01361 ± 0.01299 | 0.92 (0.87–0.97) | <0.3385 | 88.28% (85.23–90.90) | 88.33% (77.43–95.18) | <0.0001 |
| EPN3 | 0.5513 ± 0.1724 | 2.411 ± 0.04022 | 0.92 (0.89–0.95) | >1.596 | 82.04% (78.50–85.22) | 90.00% (79.49–96.24) | <0.0001 |
| ZNF8 | −0.6821 ± 0.06841 | 0.1177 ± 0.02107 | 0.92 (0.88–0.96) | >−0.3715 | 84.69% (81.33–87.65) | 88.33% (77.43–95.18) | <0.0001 |
| DTWD1 | 0.1174 ± 0.07557 | −0.7459 ± 0.02202 | 0.92 (0.89–0.95) | <−0.3011 | 84.12% (80.72–87.13) | 90.00% (79.49–96.24) | <0.0001 |
| PYGM | 1.132 ± 0.1535 | −0.3815 ± 0.02526 | 0.92 (0.88–0.96) | <0.09362 | 85.07% (81.74–88.00) | 83.33% (71.48–91.71) | <0.0001 |
| TDRD10 | 1.561 ± 0.1077 | 0.4343 ± 0.01890 | 0.91 (0.87–0.96) | <0.8479 | 84.88% (81.54–87.82) | 86.67% (75.41–94.06) | <0.0001 |
| SPINT2 | 0.9341 ± 0.1939 | 2.351 ± 0.02249 | 0.90 (0.87–0.93) | >1.981 | 81.85% (78.30–85.05) | 83.33% (71.48–91.71) | <0.0001 |
| Symbol | Description | NCBI Gene ID | Expression Pattern in This Study | Expression Patterns in Other Studies Related to Breast Cancer | Expression Patterns in Different Types of Cancer |
|---|---|---|---|---|---|
| ING3 | Inhibitor Of Growth Family Member 3 | 54556 | Downregulation | Downregulation [33,34] | Downregulated in liver cancer, head and neck cancer and colorectal cancer [35,36,37] |
| SNF1LK2 | Salt Inducible Kinase 2 | 23235 | Downregulation | Downregulation [38,39,40] | Downregulated in gastric cancer [41] |
| HBA2 | Hemoglobin Subunit Alpha 2 | 3040 | Downregulation | - | - |
| KCNIP2 | Potassium Voltage-Gated Channel Interacting Protein 2 | 30819 | Downregulation | - | - |
| SYF2 | SYF2 Pre-MRNA Splicing Factor | 25949 | Downregulation | Upregulation [23] | Upregulated in epithelial ovarian cancer [22] |
| CPN2 | Carboxypeptidase N Subunit 2 | 1370 | Downregulation | Upregulation [25] | Upregulated in lung cancer [24] |
| EPB42 | Erythrocyte Membrane Protein Band 4.2 | 2038 | Downregulation | - | Downregulated in pancreatic cancer [42] |
| EMCN | Endomucin | 51705 | Downregulation | Downregulation [43] | Downregulated in renal cancer [44] |
| CEP68 | Centrosomal Protein 68 | 23177 | Downregulation | - | - |
| HADH | Hydroxyacyl-CoA Dehydrogenase | 3033 | Downregulation | - | Downregulated in renal cancer and gastric cancer [45,46,47] |
| ADAM33 | ADAM Metallopeptidase Domain 33 | 80332 | Downregulation | Downregulation [48,49] | |
| DTWD1 | DTW Domain Containing 1 | 56986 | Downregulation | - | Downregulated in gastric cancer [50] |
| PYGM | Glycogen Phosphorylase, Muscle Associated | 5837 | Downregulation | - | Downregulated in head and neck cancer [51] |
| TDRD10 | Tudor Domain Containing 10 | 126668 | Downregulation | Downregulation [52] | - |
| AP1M2 | Adaptor Related Protein Complex 1 Subunit Mu 2 | 10053 | Upregulation | - | - |
| EVPL | Envoplakin | 2125 | Upregulation | - | - |
| TOMM40 | Translocase Of Outer Mitochondrial Membrane 40 | 10452 | Upregulation | - | - |
| EPN3 | Epsin 3 | 55040 | Upregulation | Upregulation [28] | Downregulated in gastric cancer [26] Upregulated in glioblastoma [27] |
| ZNF8 | Zinc Finger Protein 8 | 7554 | Upregulation | ||
| SPINT2 | Serine Peptidase Inhibitor, Kunitz Type 2 | 10653 | Upregulation | Upregulation [32] Downregulation [29] | Downregulated in liver, renal, gastric, cervical, prostate cancer and medulloblastoma [29,30,31] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.H.; Lee, K.E. Discovering Breast Cancer Biomarkers Candidates through mRNA Expression Analysis Based on The Cancer Genome Atlas Database. J. Pers. Med. 2022, 12, 1753. https://doi.org/10.3390/jpm12101753
Kim DH, Lee KE. Discovering Breast Cancer Biomarkers Candidates through mRNA Expression Analysis Based on The Cancer Genome Atlas Database. Journal of Personalized Medicine. 2022; 12(10):1753. https://doi.org/10.3390/jpm12101753
Chicago/Turabian StyleKim, Dong Hyeok, and Kyung Eun Lee. 2022. "Discovering Breast Cancer Biomarkers Candidates through mRNA Expression Analysis Based on The Cancer Genome Atlas Database" Journal of Personalized Medicine 12, no. 10: 1753. https://doi.org/10.3390/jpm12101753
APA StyleKim, D. H., & Lee, K. E. (2022). Discovering Breast Cancer Biomarkers Candidates through mRNA Expression Analysis Based on The Cancer Genome Atlas Database. Journal of Personalized Medicine, 12(10), 1753. https://doi.org/10.3390/jpm12101753