
Chronic Venous Disease in Pregnant Women Causes an Increase in ILK in the Placental Villi Associated with a Decrease in E-Cadherin

 ,
,  ,
, 
 ,
,  , ,
, ,  ,
,  ,
,
Abstract
:1. Introduction
2. Patients and Methods
2.1. Experimental Design
2.2. Tissue Samples
2.3. Gene Expression Studies Using Reverse Transcription-Quantitative PCR (RT-qPCR)
2.4. Immunohistochemistry Studies for Protein Expression Analysis
2.5. Statistical Analysis
3. Results
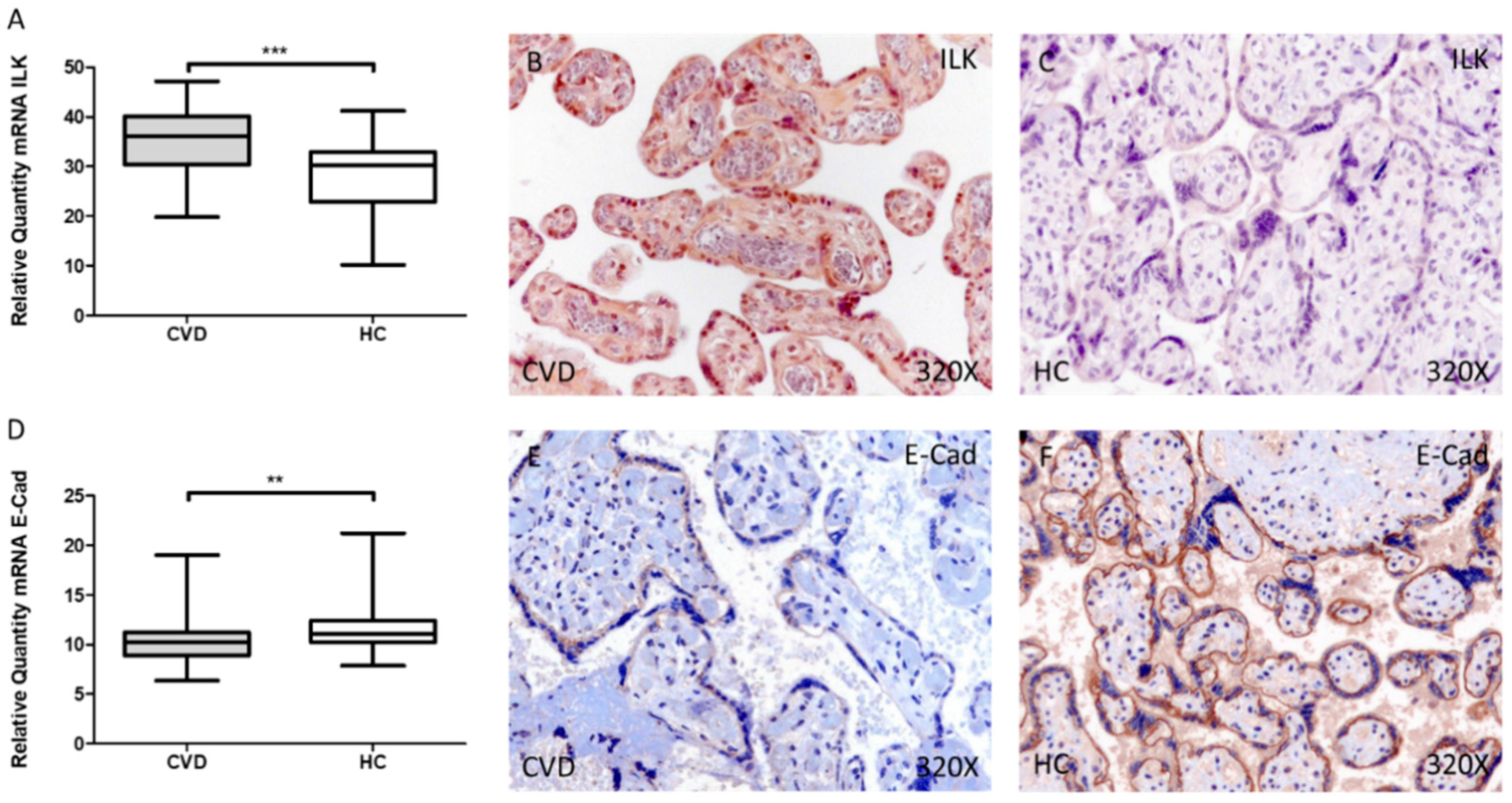
3.1. Women with CVD during Pregnancy Show an Increase in ILK Expression in Placental Villi Associated with a Decrease in E-Cadherin
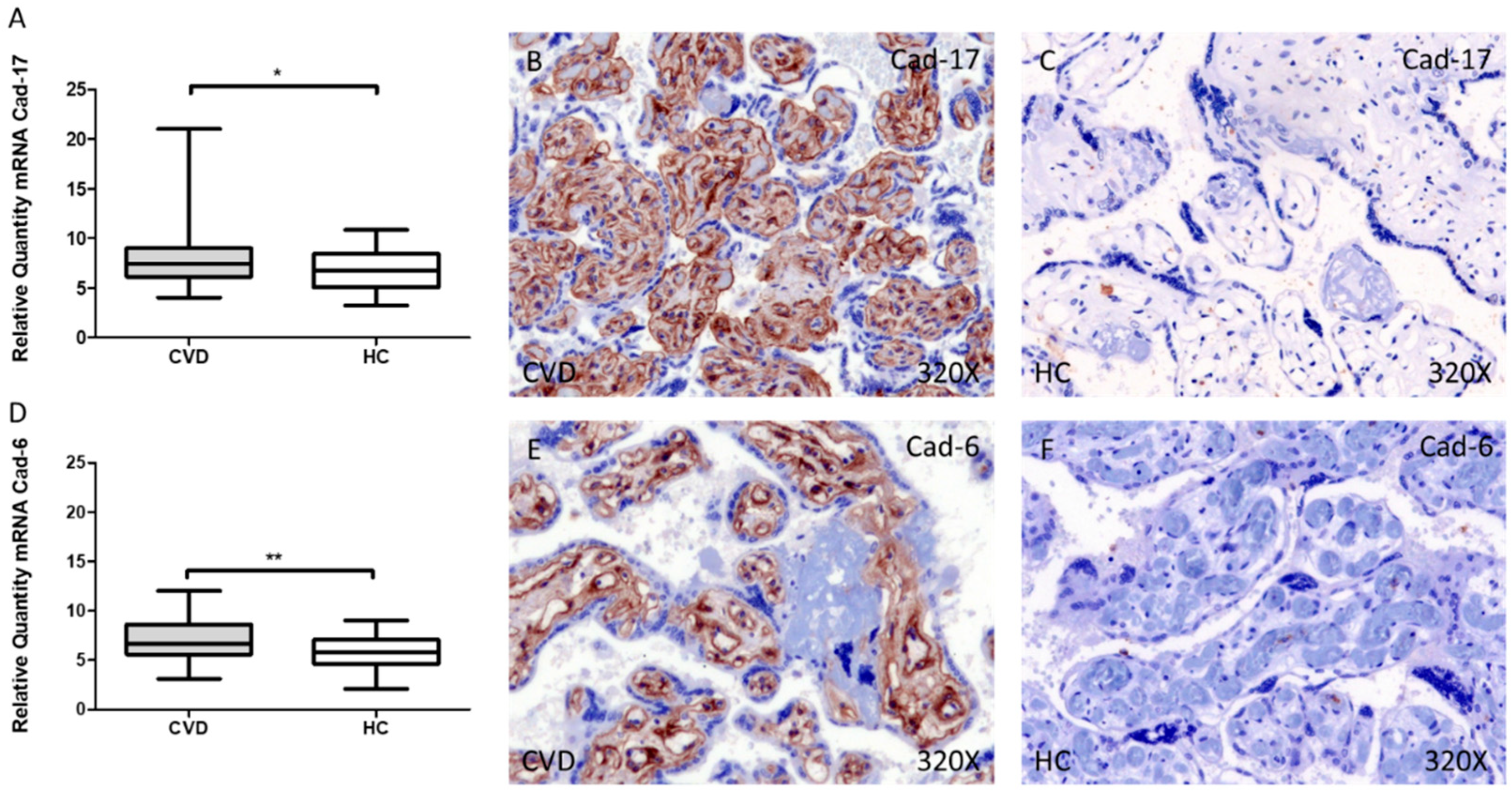
3.2. Cadherin 17 and Cadherin 6 Expression Level Is Increased in the Placental Villi of Women with CVD during Pregnancy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Youn, Y.J.; Lee, J. Chronic Venous Insufficiency and Varicose Veins of the Lower Extremities. Korean J. Intern. Med. 2019, 34, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.K.; Tan, E.L. Alterations in Physiology and Anatomy during Pregnancy. Best Pract. Res. Clin. Obstet. Gynaecol. 2013, 27, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Troiano, N.H. Physiologic and Hemodynamic Changes During Pregnancy. AACN Adv. Crit. Care 2018, 29, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Sanghavi, M.; Rutherford, J.D. Cardiovascular Physiology of Pregnancy. Circulation 2014, 130, 1003–1008. [Google Scholar] [CrossRef]
- Ortega, M.A.; Fraile-Martínez, O.; García-Montero, C.; Álvarez-Mon, M.A.; Chaowen, C.; Ruiz-Grande, F.; Pekarek, L.; Monserrat, J.; Asúnsolo, A.; García-Honduvilla, N.; et al. Understanding Chronic Venous Disease: A Critical Overview of Its Pathophysiology and Medical Management. J. Clin. Med. 2021, 10, 3239. [Google Scholar] [CrossRef]
- Tuncer Çoban, P.; Dirimeşe, E. Evaluation of quality of life after minimally invasive varicose vein treatment. Turk Gogus Kalp Damar Cerrahisi Derg. 2019, 27, 49–56. [Google Scholar] [CrossRef]
- Ortega, M.A.; Asúnsolo, Á.; Álvarez-Rocha, M.J.; Romero, B.; de León-Luis, J.; Álvarez-Mon, M.; Buján, J.; García-Honduvilla, N. Remodelling of Collagen Fibres in the Placentas of Women with Venous Insufficiency during Pregnancy. Histol. Histopathol. 2018, 33, 567–576. [Google Scholar] [CrossRef]
- de Barros Junior, N.; Del, M.; Janeiro Perez, C.; de Amorim, J.E.; Junior, F.M. Pregnancy and Lower Limb Varicose Veins: Prevalence and Risk Factors. J. Vasc. Bras. 2010, 9, 29–35. [Google Scholar]
- Rana, S.; Lemoine, E.; Granger, J.P.; Karumanchi, S.A. Preeclampsia: Pathophysiology, Challenges, and Perspectives. Circ. Res. 2019, 124, 1094–1112. [Google Scholar] [CrossRef]
- Redman, C.W.; Sargent, I.L. Pre-Eclampsia, the Placenta and the Maternal Systemic Inflammatory Response—A Review. Placenta 2003, 24, S21–S27. [Google Scholar] [CrossRef]
- Burton, G.J.; Redman, C.W.; Roberts, J.M.; Moffett, A. Pre-Eclampsia: Pathophysiology and Clinical Implications. BMJ 2019, 366, 2381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawfield, A.; Freedman, B.I. Pre-Eclampsia: The Pivotal Role of the Placenta in Its Pathophysiology and Markers for Early Detection. Ther. Adv. Cardiovasc. Dis. 2009, 3, 65–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Honduvilla, N.; Ortega, M.A.; Asúnsolo, Á.; Álvarez-Rocha, M.J.; Romero, B.; de León-Luis, J.; Álvarez-Mon, M.; Buján, J. Placentas from Women with Pregnancy-Associated Venous Insufficiency Show Villi Damage with Evidence of Hypoxic Cellular Stress. Hum. Pathol. 2018, 77, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Ortega, M.A.; Romero, B.; Asúnsolo, Á.; Martínez-Vivero, C.; Sainz, F.; Bravo, C.; de León-Luis, J.; Álvarez-Mon, M.; Buján, J.; García-Honduvilla, N. Pregnancy-Associated Venous Insufficiency Course with Placental and Systemic Oxidative Stress. J. Cell. Mol. Med. 2020, 24, 4157–4170. [Google Scholar] [CrossRef] [Green Version]
- Ortega, M.A.; Saez, M.A.; Fraile-Martínez, O.; Asúnsolo, Á.; Pekarek, L.; Bravo, C.; Coca, S.; Sainz, F.; Álvarez-Mon, M.; Buján, J.; et al. Increased Angiogenesis and Lymphangiogenesis in the Placental Villi of Women with Chronic Venous Disease during Pregnancy. Int. J. Mol. Sci. 2020, 21, 2487. [Google Scholar] [CrossRef] [Green Version]
- Ortega, M.A.; Asúnsolo, Á.; Fraile-Martínez, O.; Sainz, F.; Saez, M.A.; Bravo, C.; De León-Luis, J.A.; Alvarez-Mon, M.A.; Coca, S.; Álvarez-Mon, M.; et al. An Increase in Elastogenic Components in the Placental Villi of Women with Chronic Venous Disease during Pregnancy Is Associated with Decreased EGFL7 Expression Level. Mol. Med. Rep. 2021, 24, 1–9. [Google Scholar] [CrossRef]
- Ortega, M.A.; Fraile-Martínez, O.; Saez, M.A.; Álvarez-Mon, M.A.; Gómez-Lahoz, A.M.; Bravo, C.; Luis, J.A.L.; Sainz, F.; Coca, S.; Asúnsolo, Á.; et al. Abnormal Proinflammatory and Stressor Environmental with Increased the Regulatory Cellular IGF-1/PAPP-A/STC and Wnt-1/β-Catenin Canonical Pathway in Placenta of Women with Chronic Venous Disease during Pregnancy. Int. J. Med. Sci. 2021, 18, 2814–2827. [Google Scholar] [CrossRef]
- Wu, C.; Dedhar, S. Integrin-Linked Kinase (ILK) and Its Interactors: A New Paradigm for the Coupling of Extracellular Matrix to Actin Cytoskeleton and Signaling Complexes. J. Cell Biol. 2001, 155, 505–510. [Google Scholar] [CrossRef]
- Hannigan, G.E.; Coles, J.G.; Dedhar, S. Integrin-Linked Kinase at the Heart of Cardiac Contractility, Repair, and Disease. Circ. Res. 2007, 100, 1408–1414. [Google Scholar] [CrossRef] [Green Version]
- Elustondo, P.A.; Hannigan, G.E.; Caniggia, I.; MacPhee, D.J. Integrin-Linked Kinase (ILK) Is Highly Expressed in First Trimester Human Chorionic Villi and Regulates Migration of a Human Cytotrophoblast-Derived Cell Line. Biol. Reprod. 2006, 74, 959–968. [Google Scholar] [CrossRef] [Green Version]
- Yen, C.-F.; Wang, H.-S.; Lee, C.-L.; Liao, S.-K. Roles of Integrin-Linked Kinase in Cell Signaling and Its Perspectives as a Therapeutic Target. Gynecol. Minim. Invasive Ther. 2014, 3, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Van Roy, F.; Berx, G. The cell-cell adhesion molecule E-cadherin. Cell Mol. Life Sci. 2008, 65, 3756–3788. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.M.; Lacey, H.A.; Baker, P.N.; Crocker, I.P. E-Cadherin in the Assessment of Aberrant Placental Cytotrophoblast Turnover in Pregnancies Complicated by Pre-Eclampsia. Histochem. Cell Biol. 2005, 124, 499–506. [Google Scholar] [CrossRef]
- Casal, J.I.; Bartolomé, R.A. Beyond N-Cadherin, Relevance of Cadherins 5, 6 and 17 in Cancer Progression and Metastasis. Int. J. Mol. Sci. 2019, 20, 3373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lurie, F.; Passman, M.; Meisner, M.; Dalsing, M.; Masuda, E.; Welch, H.; Bush, R.L.; Blebea, J.; Carpentier, P.H.; de Maeseneer, M.; et al. The 2020 Update of the CEAP Classification System and Reporting Standards. J. Vasc. Surg. Venous Lymphat. Disord. 2020, 8, 342–352. [Google Scholar] [CrossRef]
- Ortega, M.A.; Asúnsolo, Á.; Pekarek, L.; Alvarez-Mon, M.A.; Delforge, A.; Sáez, M.A.; Coca, S.; Sainz, F.; Mon, M.Á.; Buján, J.; et al. Histopathological Study of JNK in Venous Wall of Patients with Chronic Venous Insufficiency Related to Osteogenesis Process. Int. J. Med. Sci. 2021, 18, 1921–1934. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Chomczynski, P.; Sacchi, N. The Single-Step Method of RNA Isolation by Acid Guanidinium Thiocyanate-Phenol-Chloroform Extraction: Twenty-Something Years On. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef]
- Ortega, M.A.; Asúnsolo, Á.; Leal, J.; Romero, B.; Alvarez-Rocha, M.J.; Sainz, F.; Álvarez-Mon, M.; Buján, J.; García-Honduvilla, N. Implication of the PI3K/Akt/MTOR Pathway in the Process of Incompetent Valves in Patients with Chronic Venous Insufficiency and the Relationship with Aging. Oxidative Med. Cell. Longev. 2018, 2018, 1–14. [Google Scholar] [CrossRef]
- Vallone, P.M.; Butler, J.M. AutoDimer: A Screening Tool for Primer-Dimer and Hairpin Structures. BioTechniques 2004, 37, 226–231. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A Tool to Design Target-Specific Primers for Polymerase Chain Reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.J.; Jeon, R.H.; Kim, H.D.; Hwang, J.C.; Lee, H.J.; Bae, S.G.; Lee, S.L.; Rho, G.J.; Kim, S.J.; Lee, W.J. TATA Box Binding Protein and Ribosomal Protein 4 Are Suitable Reference Genes for Normalization during Quantitative Polymerase Chain Reaction Study in Bovine Mesenchymal Stem Cells. Asian-Australas. J. Anim. Sci. 2020, 33, 2021–2030. [Google Scholar] [CrossRef] [PubMed]
- Ortega, M.A.; Asúnsolo, Á.; Romero, B.; Álvarez-Rocha, M.J.; Sainz, F.; Leal, J.; Álvarez-Mon, M.; Buján, J.; García-Honduvilla, N. Unravelling the Role of Mapks (Erk1/2) in Venous Reflux in Patients with Chronic Venous Disorder. Cells Tissues Organs 2019, 206, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Ortega, M.A.; Fraile-Martínez, O.; Pekarek, L.; Alvarez-Mon, M.A.; Asúnsolo, Á.; Sanchez-Trujillo, L.; Coca, S.; Buján, J.; Álvarez-Mon, M.; García-Honduvilla, N.; et al. Defective Expression of the Peroxisome Regulators PPARα Receptors and Lysogenesis with Increased Cellular Senescence in the Venous Wall of Chronic Venous Disorder. Histol. Histopathol. 2021, 36, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Sanmartín-Salinas, P.; Guijarro, L.G. Overexpression of IRS-4 Correlates with Procaspase 3 Levels in Tumoural Tissue of Patients with Colorectal Cancer. J. Oncol. 2018, 2018, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cristóbal, L.; Ortega, M.A.; Asúnsolo, Á.; Romero, B.; Álvarez-Mon, M.; Buján, J.; Maldonado, A.A.; García-Honduvilla, N. Human Skin Model for Mimic Dermal Studies in Pathology with a Clinical Implication in Pressure Ulcers. Histol. Histopathol. 2018, 33, 959–970. [Google Scholar] [CrossRef]
- Yan, Z.; Yin, H.; Wang, R.; Wu, D.; Sun, W.; Liu, B.; Su, Q. Overexpression of Integrin-Linked Kinase (ILK) Promotes Migration and Invasion of Colorectal Cancer Cells by Inducing Epithelial-Mesenchymal Transition via NF-ΚB Signaling. Acta Histochem. 2014, 116, 527–533. [Google Scholar] [CrossRef]
- Cui, K.; Yan, T.; Luo, Q.; Zheng, Y.; Liu, X.; Huang, X.; Zou, L. Ultrasound Microbubble-Mediated Delivery of Integrin-Linked Kinase Gene Improves Endothelial Progenitor Cells Dysfunction in Pre-Eclampsia. DNA Cell Biol. 2014, 33, 301–310. [Google Scholar] [CrossRef] [Green Version]
- Petropoulos, S.; Guillemin, C.; Ergaz, Z.; Dimov, S.; Suderman, M.; Weinstein-Fudim, L.; Ornoy, A.; Szyf, M. Gestational Diabetes Alters Offspring DNA Methylation Profiles in Human and Rat: Identification of Key Pathways Involved in Endocrine System Disorders, Insulin Signaling, Diabetes Signaling, and ILK Signaling. Endocrinology 2015, 156, 2222–2238. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Zhang, Y.; Wu, C. Integrin-Linked Kinase Is Localized to Cell-Matrix Focal Adhesions but Not Cell-Cell Adhesion Sites and the Focal Adhesion Localization of Integrin-Linked Kinase Is Regulated by the PINCH-Binding ANK Repeats. J. Cell Sci. 1999, 112, 4589–4599. [Google Scholar] [CrossRef]
- Elad, N.; Volberg, T.; Patla, I.; Hirschfeld-Warneken, V.; Grashoff, C.; Spatz, J.P.; Fässler, R.; Geiger, B.; Medalia, O. The Role of Integrin-Linked Kinase in the Molecular Architecture of Focal Adhesions. J. Cell Sci. 2013, 126, 4099–4107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, R.; Macklin, W.B. Integrin-Linked Kinase (ILK) Deletion Disrupts Oligodendrocyte Development by Altering Cell Cycle. J. Neurosci. 2017, 37, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Radeva, G.; Petrocelli, T.; Behrend, E.; Leung-Hagesteijn, C.; Filmus, J.; Slingerland, J.; Dedhar, S. Overexpression of the Integrin-Linked Kinase Promotes Anchorage-Independent Cell Cycle Progression. J. Biol. Chem. 1997, 272, 13937–13944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, Y.; Zhong, X.; Flynn, D.C.; Zheng, J.Z.; Qiao, M.; Wu, C.; Dedhar, S.; Shi, X.; Jiang, B.H. ILK Mediates Actin Filament Rearrangements and Cell Migration and Invasion through PI3K/Akt/Rac1 Signaling. Oncogene 2005, 24, 3154–3165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dedhar, S. Cell-Substrate Interactions and Signaling through ILK. Curr. Opin. Cell Biol. 2000, 12, 250–256. [Google Scholar] [CrossRef]
- Attwell, S.; Mills, J.; Troussard, A.; Wu, C.; Dedhar, S. Integration of Cell Attachment, Cytoskeletal Localization, and Signaling by Integrin-Linked Kinase (ILK), CH-ILKBP, and the Tumor Suppressor PTEN. Mol. Biol. Cell 2003, 14, 4813–4825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troussard, A.A.; Costello, P.; Yoganathan, T.N.; Kumagai, S.; Roskelley, C.D.; Dedhar, S. The Integrin Linked Kinase (ILK) Induces an Invasive Phenotype via AP-1 Transcription Factor-Dependent Upregulation of Matrix Metalloproteinase 9 (MMP-9). Oncogene 2000, 19, 5444–5452. [Google Scholar] [CrossRef] [Green Version]
- Kokkinos, M.I.; Murthi, P.; Wafai, R.; Thompson, E.W.; Newgreen, D.F. Cadherins in the Human Placenta—Epithelial-Mesenchymal Transition (EMT) and Placental Development. Placenta 2010, 31, 747–755. [Google Scholar] [CrossRef]
- Incebiyik, A.; Kocarslan, S.; Camuzcuoglu, A.; Hilali, N.G.; Incebiyik, H.; Camuzcuoglu, H. Trophoblastic E-Cadherin and TGF-Beta Expression in Placenta Percreta and Normal Pregnancies. J. Matern.-Fetal Neonatal Med. 2016, 29, 126–129. [Google Scholar] [CrossRef]
- Duzyj, C.M.; Buhimschi, I.A.; Motawea, H.; Laky, C.A.; Cozzini, G.; Zhao, G.; Funai, E.F.; Buhimschi, C.S. The Invasive Phenotype of Placenta Accreta Extravillous Trophoblasts Associates with Loss of E-Cadherin. Placenta 2015, 36, 645–651. [Google Scholar] [CrossRef]
- Li, H.W.; Cheung, A.N.; Tsao, S.W.; Cheung, A.L.M. Expression of E-Cadherin and Beta-Catenin in Trophoblastic Tissue in Normal and Pathological Pregnancies. Int. J. Gynecol. Pathol. 2003, 22, 63–70. [Google Scholar] [CrossRef] [PubMed]
- McPhee, T.R.; McDonald, P.C.; Oloumi, A.; Dedhar, S. Integrin-Linked Kinase Regulates E-Cadherin Expression through PARP-1. Dev. Dyn. 2008, 237, 2737–2747. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Santos, L.; Dimitriadis, E. Characterization of the role for cadherin 6 in the regulation of human endometrial receptivity. Reprod. Biol. Endocrinol. 2020, 18, 1–10. [Google Scholar] [CrossRef] [PubMed]
- MacCalman, C.D.; Getsios, S.; Chen, G.T. Type 2 cadherins in the human endometrium and placenta: Their putative roles in human implantation and placentation. Am. J. Reprod. Immunol. 1998, 39, 96–107. [Google Scholar] [CrossRef]
- Ordóñez, N.G. Cadherin 17 is a novel diagnostic marker for adenocarcinomas of the digestive system. Adv. Anat. Pathol. 2014, 21, 131–137. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| CVD (n = 62) | HC (n = 52) | |
|---|---|---|
| Median age (IQR), years | 33 (22–40) | 34 (27–41) |
| Median gestational age (IQR), weeks | 40.5 (39–41.5) | 41 (39–42) |
| C-section delivery, n (%) | 12 (19.4) | 9 (17.3) |
| Vaginal delivery, n (%) | 50 (80.6) | 43 (82.7) |
| Varicose vein (CEAP), n (%) | ||
| CEAP 1 | 37 (59.7) | 0 (0) |
| CEAP 2 | 21 (33.8) | 0 (0) |
| CEAP 3 | 4 (6.5) | 0 (0) |
| Previous pregnancies, n (%) | 33 (53.2) | 19 (36.5) |
| Previous abortions, n (%) | 14 (22.6) | 9 (17.3) |
| Regular menstrual cycles, n (%) | 50 (80.6) | 42 (80.7) |
| Sedentary profession, n (%) | 41 (66.1) | 40 (76.9) |
| GENE | SEQUENCE Fwd (5′→3′) | SEQUENCE Rev (5′→3′) | Temp |
|---|---|---|---|
| TBP | TGCACAGGAGCCAAGAGTGAA | CACATCACAGCTCCCCACCA | 60 °C |
| ILK | TCCCAAGTAAGGAACGGAGC | CACCACCAGACATGAGCACT | 59 °C |
| E-Cad | GTGAACACCTACAATGCCGC | CCCAGGGGACAAGGGTATGA | 59 °C |
| Cad-17 | GCTCCTGGGAGGTAAGTAGA | ACCCTCGGCAAAGCTCC | 57 °C |
| Cad-6 | AGCTATTTCCTGCTTTCAGGGT | GGTGGGAAGGAAGTGAGACG | 60 °C |
| Antigen | Species | Dilution | Provider | Protocol Specifications |
|---|---|---|---|---|
| ILK | Rabbit | 1:50 | Abcam (ab52,480) | 10 mM Sodium citrate pH = 6 before incubation with blocking solution |
| E-Cad | Mouse | 1:250 | Vitro (MAD-000761QD-3/V) | - |
| Cad-17 | Rabbit | 1:250 | Vitro (MAD-000737QD-3/V) | - |
| Cad-6 | Mouse | 1:250 | Vitro (MAD-000582QD-3/V) | - |
| IgG (Mouse) | Goat | 1:300 | Sigma-Aldrich (F2012/045K6072) | - |
| IgG (Rabbit) | Mouse | 1:1000 | Sigma-Aldrich (RG-96/B5283) | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortega, M.A.; Chaowen, C.; Fraile-Martinez, O.; García-Montero, C.; Saez, M.A.; Cruza, I.; Pereda-Cerquella, C.; Alvarez-Mon, M.A.; Guijarro, L.G.; Fatych, Y.; et al. Chronic Venous Disease in Pregnant Women Causes an Increase in ILK in the Placental Villi Associated with a Decrease in E-Cadherin. J. Pers. Med. 2022, 12, 277. https://doi.org/10.3390/jpm12020277
Ortega MA, Chaowen C, Fraile-Martinez O, García-Montero C, Saez MA, Cruza I, Pereda-Cerquella C, Alvarez-Mon MA, Guijarro LG, Fatych Y, et al. Chronic Venous Disease in Pregnant Women Causes an Increase in ILK in the Placental Villi Associated with a Decrease in E-Cadherin. Journal of Personalized Medicine. 2022; 12(2):277. https://doi.org/10.3390/jpm12020277
Chicago/Turabian StyleOrtega, Miguel A., Chen Chaowen, Oscar Fraile-Martinez, Cielo García-Montero, Miguel A. Saez, Iris Cruza, Claude Pereda-Cerquella, Miguel Angel Alvarez-Mon, Luis G. Guijarro, Yuliia Fatych, and et al. 2022. "Chronic Venous Disease in Pregnant Women Causes an Increase in ILK in the Placental Villi Associated with a Decrease in E-Cadherin" Journal of Personalized Medicine 12, no. 2: 277. https://doi.org/10.3390/jpm12020277
APA StyleOrtega, M. A., Chaowen, C., Fraile-Martinez, O., García-Montero, C., Saez, M. A., Cruza, I., Pereda-Cerquella, C., Alvarez-Mon, M. A., Guijarro, L. G., Fatych, Y., Menor-Salván, C., Alvarez-Mon, M., De Leon-Luis, J., Buján, J., Garcia-Honduvilla, N., Bravo, C., & Asúnsolo-del-Barco, A. (2022). Chronic Venous Disease in Pregnant Women Causes an Increase in ILK in the Placental Villi Associated with a Decrease in E-Cadherin. Journal of Personalized Medicine, 12(2), 277. https://doi.org/10.3390/jpm12020277