
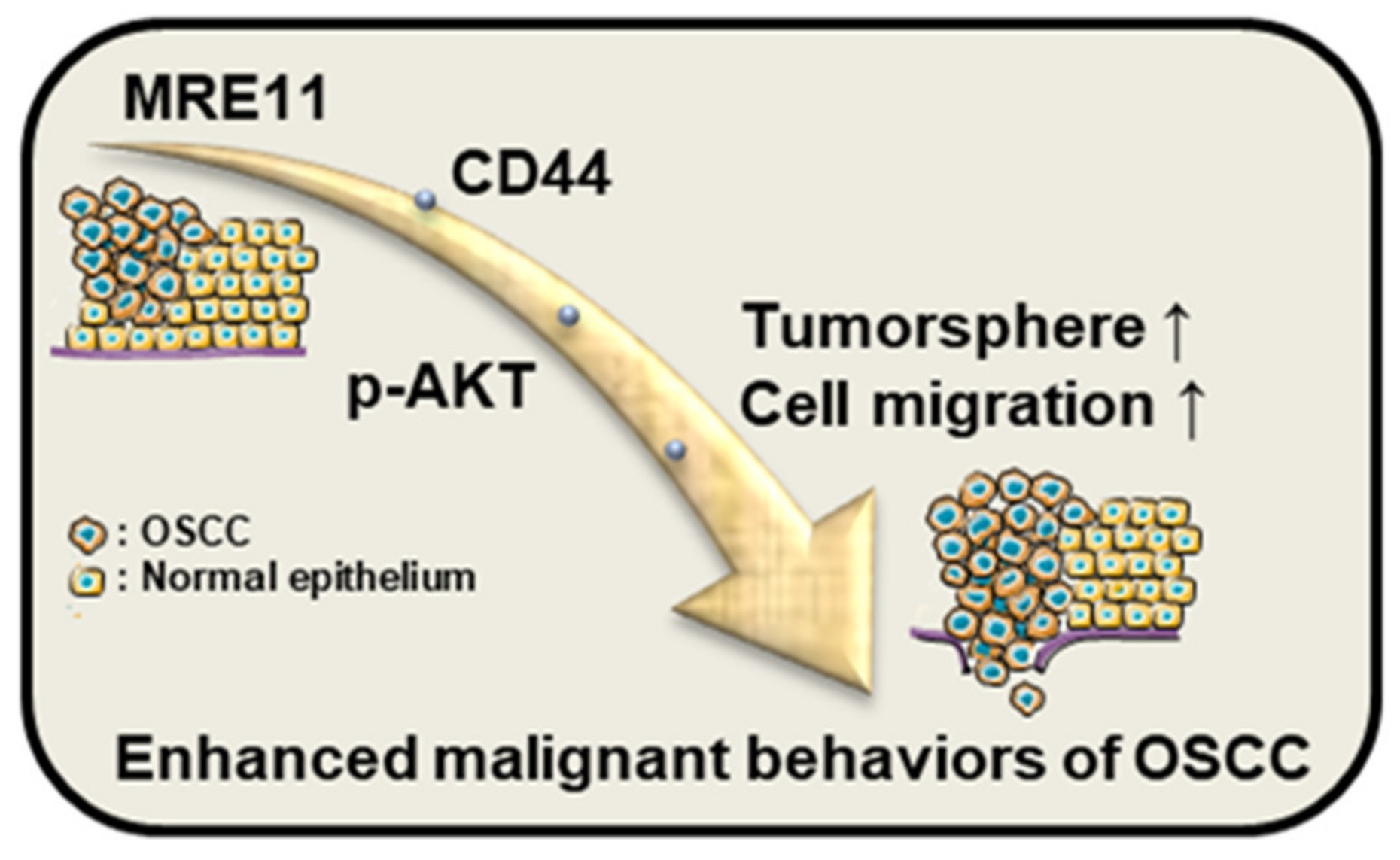
CD44 Mediates Oral Squamous Cell Carcinoma-Promoting Activity of MRE11 via AKT Signaling

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Specimens
2.2. Cell Culture
2.3. Gene Overexpression and Knockdown
2.4. Western Blot
2.5. Tumorsphere Formation Assay
2.6. Transwell Cell Migration Assay
2.7. Immunohistochemistry (IHC)
2.8. Metastatic Mouse Model
2.9. Oncomine Database Analysis
2.10. The Cancer Genome Atlas (TCGA) Database Analysis
2.11. Statistical Analysis
3. Results
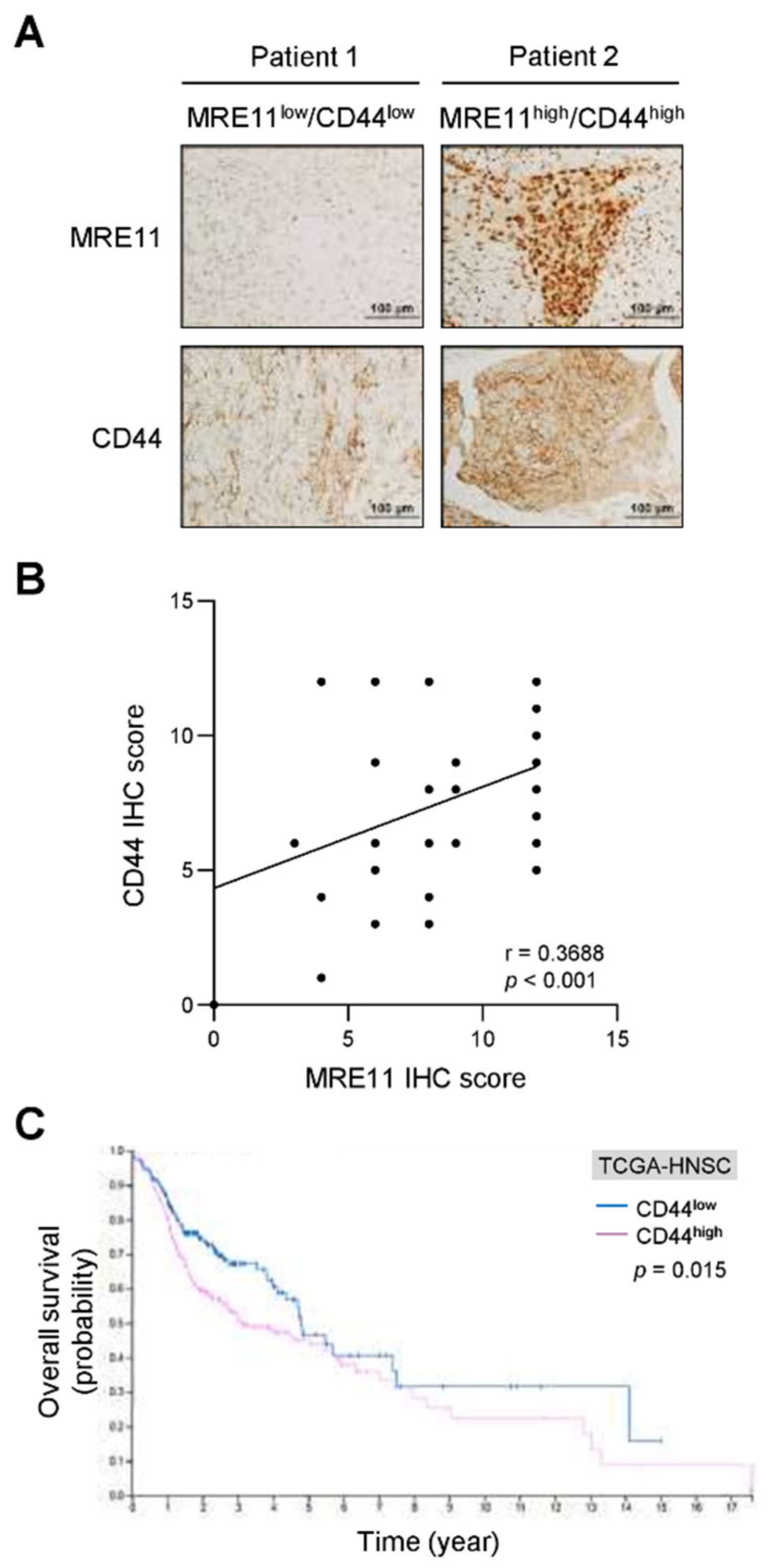
3.1. Expression of MRE11 and CD44 in OSCC Patients
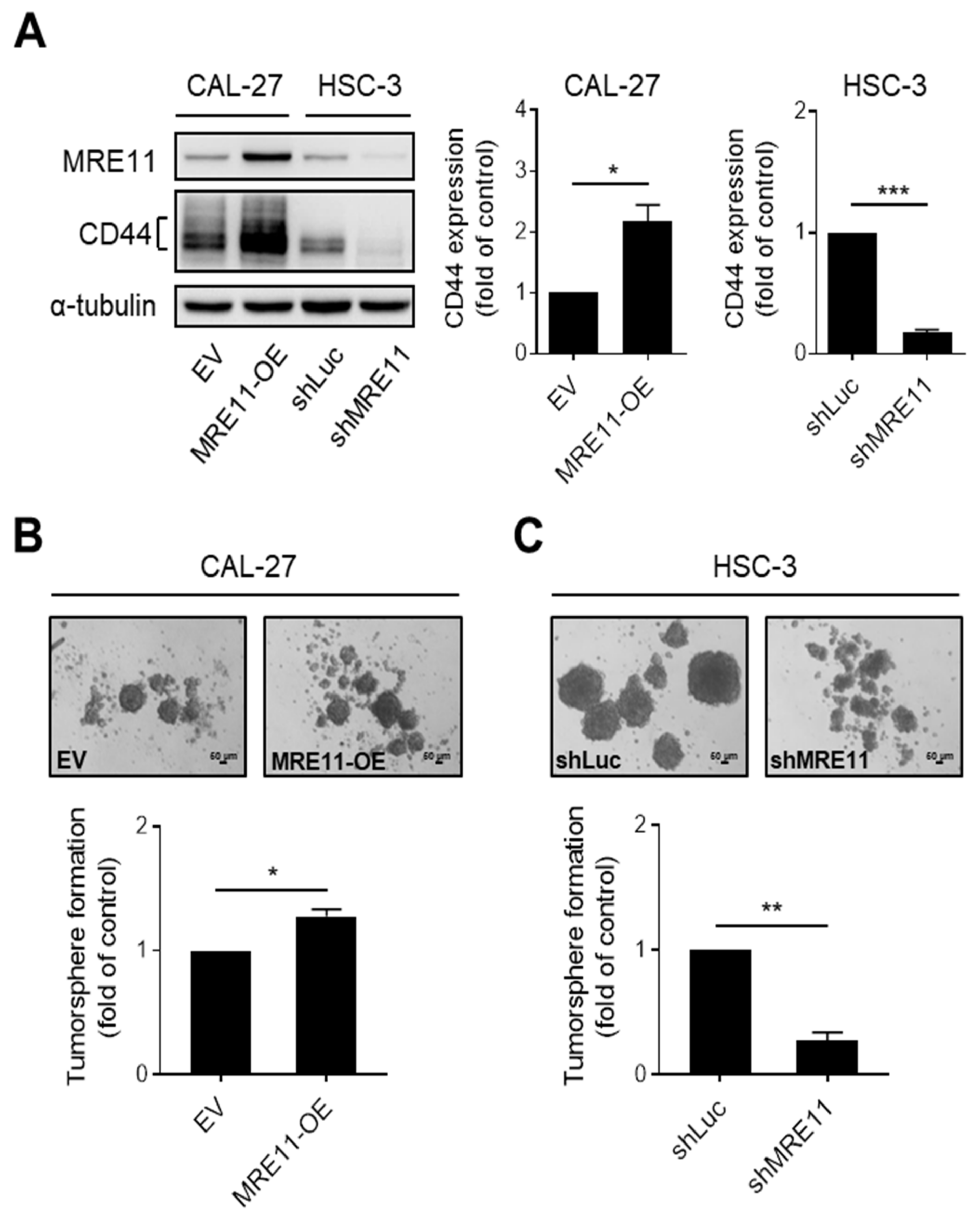
3.2. MRE11 Regulates CD44 Expression and Tumorsphere Formation in OSCC Cells
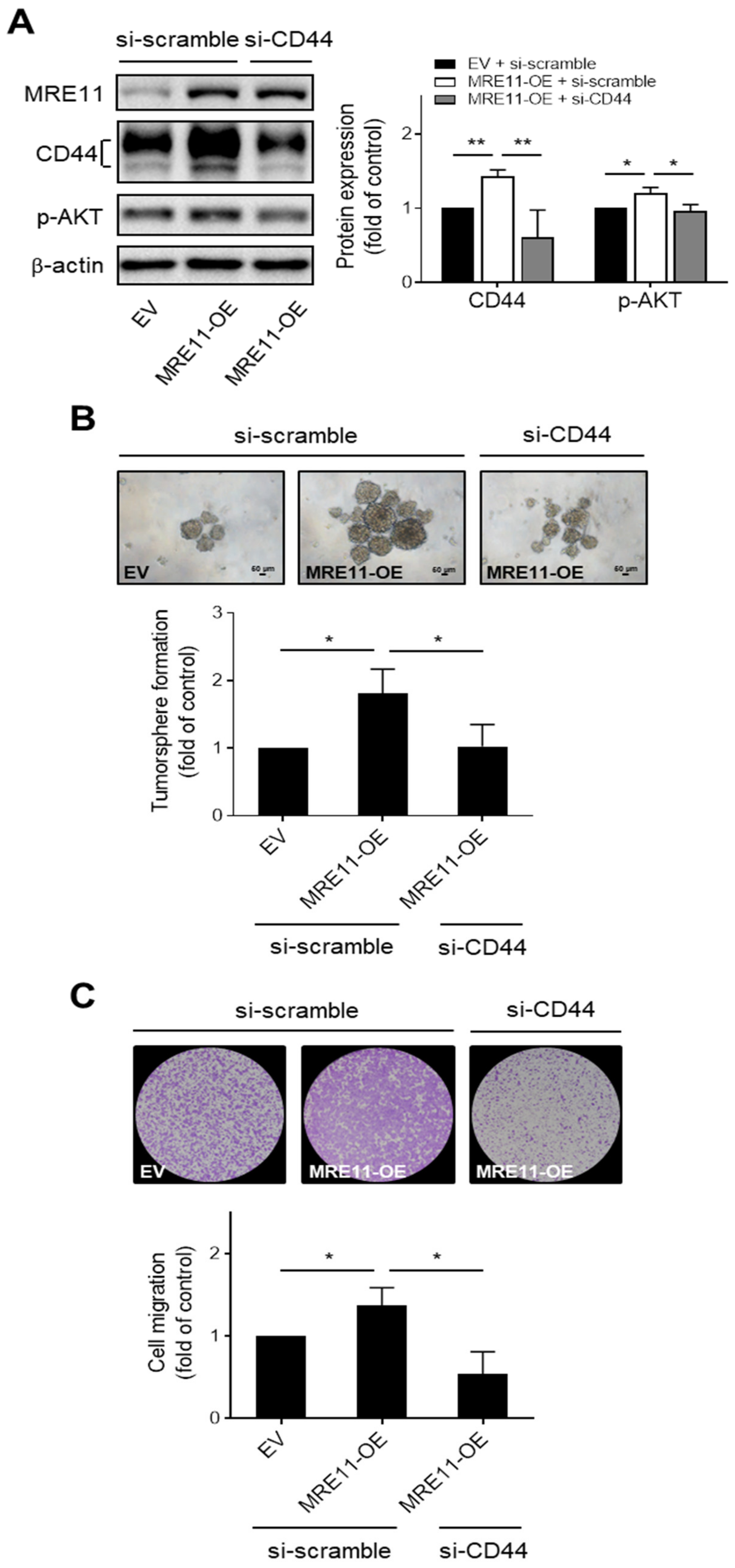
3.3. CD44 Mediates MRE11-Promoted AKT Phosphorylation, Tumorsphere Formation, and Migration Ability in OSCC Cells
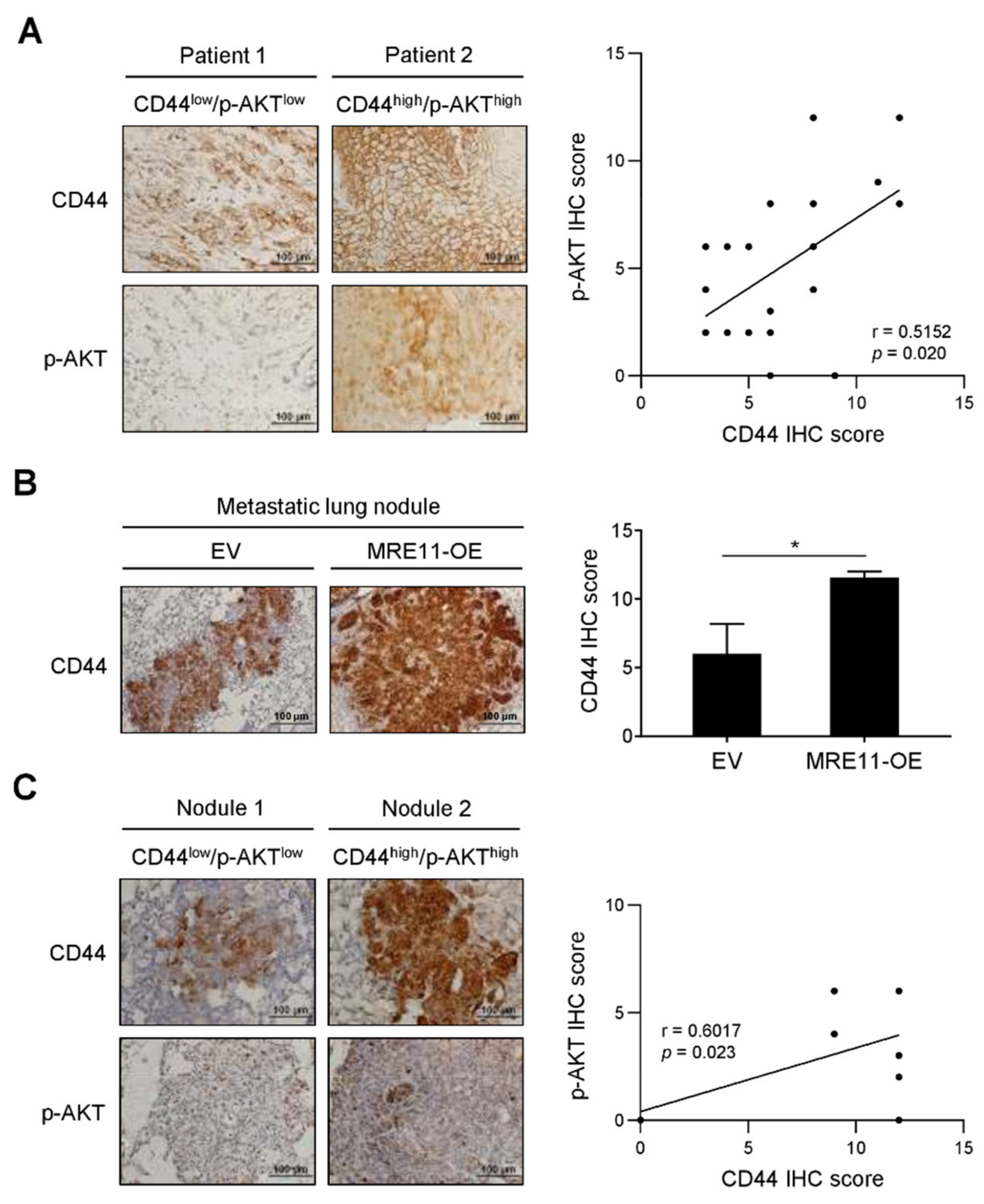
3.4. Correlation between CD44 Expression and AKT Phosphorylation in Tumor Tissues from OSCC Patients and In Vivo Metastatic Xenograft Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Du, M.; Nair, R.; Jamieson, L.; Liu, Z.; Bi, P. Incidence trends of lip, oral cavity, and pharyngeal cancers: Global burden of disease 1990–2017. J. Dent. Res. 2020, 99, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Rivera, C. Essentials of oral cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 11884–11894. [Google Scholar] [PubMed]
- Cheng, Y.; Li, S.; Gao, L.; Zhi, K.; Ren, W. The molecular basis and therapeutic aspects of cisplatin resistance in oral squamous cell carcinoma. Front. Oncol. 2021, 11, 761379. [Google Scholar] [CrossRef] [PubMed]
- Vig, N.; Mackenzie, I.C.; Biddle, A. Phenotypic plasticity and epithelial-to-mesenchymal transition in the behaviour and therapeutic response of oral squamous cell carcinoma. J. Oral Pathol. Med. 2015, 44, 649–655. [Google Scholar] [CrossRef]
- Bai, Y.; Sha, J.; Okui, T.; Moriyama, I.; Ngo, H.X.; Tatsumi, H.; Kanno, T. The epithelial-mesenchymal transition influences the resistance of oral squamous cell carcinoma to monoclonal antibodies via its effect on energy homeostasis and the tumor microenvironment. Cancers 2021, 13, 5905. [Google Scholar] [CrossRef]
- Coletta, R.D.; Yeudall, W.A.; Salo, T. Grand challenges in oral cancers. Front. Oral Health 2020, 1, 3. [Google Scholar] [CrossRef]
- Mughees, M.; Sengupta, A.; Khowal, S.; Wajid, S. Mechanism of tumour microenvironment in the progression and development of oral cancer. Mol. Biol. Rep. 2021, 48, 1773–1786. [Google Scholar] [CrossRef]
- Richard, V.; Sebastian, P.; Nair, M.G.; Nair, S.N.; Malieckal, T.T.; Santhosh Kumar, T.R.; Pillai, M.R. Multiple drug resistant, tumorigenic stem-like cells in oral cancer. Cancer Lett. 2013, 338, 300–316. [Google Scholar] [CrossRef]
- Sinha, N.; Mukhopadhyay, S.; Das, D.N.; Panda, P.K.; Bhutia, S.K. Relevance of cancer initiating/stem cells in carcinogenesis and therapy resistance in oral cancer. Oral Oncol. 2013, 49, 854–862. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, M.; Luo, J.; Zhou, H. Radiotherapy targeting cancer stem cells “awakens” them to induce tumour relapse and metastasis in oral cancer. Int. J. Oral Sci. 2020, 12, 19. [Google Scholar] [CrossRef]
- Goodison, S.; Urquidi, V.; Tarin, D. CD44 cell adhesion molecules. Mol. Pathol. 1999, 52, 189–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponta, H.; Sherman, L.; Herrlich, P.A. CD44: From adhesion molecules to signalling regulators. Nat. Rev. Mol. Cell Biol. 2003, 4, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Tahmasebi, E.; Alikhani, M.; Yazdanian, A.; Yazdanian, M.; Tebyanian, H.; Seifalian, A. The current markers of cancer stem cell in oral cancers. Life Sci. 2020, 249, 117483. [Google Scholar] [CrossRef] [PubMed]
- Hassn Mesrati, M.; Syafruddin, S.E.; Mohtar, M.A.; Syahir, A. CD44: A multifunctional mediator of cancer progression. Biomolecules 2021, 11, 1850. [Google Scholar] [CrossRef]
- Morath, I.; Hartmann, T.N.; Orian-Rousseau, V. CD44: More than a mere stem cell marker. Int. J. Biochem. Cell Biol. 2016, 81, 166–173. [Google Scholar] [CrossRef]
- Wang, S.J.; Bourguignon, L.Y. Role of hyaluronan-mediated CD44 signaling in head and neck squamous cell carcinoma progression and chemoresistance. Am. J. Pathol. 2011, 178, 956–963. [Google Scholar] [CrossRef]
- Lee, J.W.; Lee, H.Y. Targeting cancer stem cell markers or pathways: A potential therapeutic strategy for oral cancer treatment. Int. J. Stem Cells 2021, 14, 386–399. [Google Scholar] [CrossRef]
- Shahoumi, L.A. Oral cancer stem cells: Therapeutic implications and challenges. Front. Oral Health 2021, 2, 685236. [Google Scholar] [CrossRef]
- Hosoya, N.; Miyagawa, K. Targeting DNA damage response in cancer therapy. Cancer Sci. 2014, 105, 370–388. [Google Scholar] [CrossRef] [Green Version]
- Abad, E.; Graifer, D.; Lyakhovich, A. DNA damage response and resistance of cancer stem cells. Cancer Lett. 2020, 474, 106–117. [Google Scholar] [CrossRef]
- Williams, R.S.; Moncalian, G.; Williams, J.S.; Yamada, Y.; Limbo, O.; Shin, D.S.; Groocock, L.M.; Cahill, D.; Hitomi, C.; Guenther, G.; et al. Mre11 dimers coordinate DNA end bridging and nuclease processing in double-strand-break repair. Cell 2008, 135, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.Y.; Hung, A.C.; Lo, S.; Hsieh, Y.C.; Yuan, S.F. MRE11 as a molecular signature and therapeutic target for cancer treatment with radiotherapy. Cancer Lett. 2021, 514, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Situ, Y.; Chung, L.; Lee, C.S.; Ho, V. MRN (MRE11-RAD50-NBS1) complex in human cancer and prognostic implications in colorectal cancer. Int. J. Mol. Sci. 2019, 20, 816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.Y.; Chen, Y.K.; Lo, S.; Chi, T.C.; Chen, Y.H.; Hu, S.C.; Chen, Y.W.; Jiang, S.S.; Tsai, F.Y.; Liu, W.; et al. MRE11 promotes oral cancer progression through RUNX2/CXCR4/AKT/FOXA2 signaling in a nuclease-independent manner. Oncogene 2021, 40, 3510–3532. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.; Chen, H.; Lo, P.K. In vitro tumorsphere formation assays. Bio-Protocol 2013, 3, e325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.Y.; Wang, Y.Y.; Lo, S.; Tseng, L.M.; Chen, D.R.; Wu, Y.C.; Hou, M.F.; Yuan, S.F. Visfatin mediates malignant behaviors through adipose-derived stem cells intermediary in breast cancer. Cancers 2020, 12, 29. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Y.; Chen, H.D.; Lo, S.; Chen, Y.K.; Huang, Y.C.; Hu, S.C.; Hsieh, Y.C.; Hung, A.C.; Hou, M.F.; Yuan, S.F. Visfatin enhances breast cancer progression through CXCL1 induction in tumor-associated macrophages. Cancers 2020, 12, 3526. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Vadhan, A.; Chen, P.H.; Lee, Y.L.; Chao, C.Y.; Cheng, K.H.; Chang, Y.C.; Hu, S.C.; Yuan, S.F. CD44 promotes lung cancer cell metastasis through ERK-ZEB1 signaling. Cancers 2021, 13, 4057. [Google Scholar] [CrossRef]
- Rhodes, D.R.; Yu, J.; Shanker, K.; Deshpande, N.; Varambally, R.; Ghosh, D.; Barrette, T.; Pandey, A.; Chinnaiyan, A.M. ONCOMINE: A cancer microarray database and integrated data-mining platform. Neoplasia 2004, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Emich, H.; Chapireau, D.; Hutchison, I.; Mackenzie, I. The potential of CD44 as a diagnostic and prognostic tool in oral cancer. J. Oral Pathol. Med. 2015, 44, 393–400. [Google Scholar] [CrossRef]
- Xu, H.; Niu, M.; Yuan, X.; Wu, K.; Liu, A. CD44 as a tumor biomarker and therapeutic target. Exp. Hematol. Oncol. 2020, 9, 36. [Google Scholar] [CrossRef] [PubMed]
- Weiswald, L.B.; Bellet, D.; Dangles-Marie, V. Spherical cancer models in tumor biology. Neoplasia 2015, 17, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, R.C.; Edwards, N.S.; Burns, G.F.; Fisher, D.E. A CD44 survival pathway triggers chemoresistance via lyn kinase and phosphoinositide 3-kinase/Akt in colon carcinoma cells. Cancer Res. 2001, 61, 5275–5283. [Google Scholar] [PubMed]
- Yu, S.; Cai, X.; Wu, C.; Wu, L.; Wang, Y.; Liu, Y.; Yu, Z.; Qin, S.; Ma, F.; Thiery, J.P.; et al. Adhesion glycoprotein CD44 functions as an upstream regulator of a network connecting ERK, AKT and Hippo-YAP pathways in cancer progression. Oncotarget 2015, 6, 2951–2965. [Google Scholar] [CrossRef] [Green Version]
- Gomez, K.E.; Wu, F.; Keysar, S.B.; Morton, J.J.; Miller, B.; Chimed, T.S.; Le, P.N.; Nieto, C.; Chowdhury, F.N.; Tyagi, A.; et al. Cancer cell CD44 mediates macrophage/monocyte-driven regulation of head and neck cancer stem cells. Cancer Res. 2020, 80, 4185–4198. [Google Scholar] [CrossRef]
- Brabletz, T.; Jung, A.; Spaderna, S.; Hlubek, F.; Kirchner, T. Migrating cancer stem cells—An integrated concept of malignant tumour progression. Nat. Rev. Cancer 2005, 5, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Ingangi, V.; Minopoli, M.; Ragone, C.; Motti, M.L.; Carriero, M.V. Role of microenvironment on the fate of disseminating cancer stem cells. Front. Oncol. 2019, 9, 82. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, J.; Ito, E.; Fujimoto, J.; Watanabe, S.; Semba, K. Comparative analysis of gene regulatory networks of highly metastatic breast cancer cells established by orthotopic transplantation and intra-circulation injection. Int. J. Oncol. 2017, 50, 497–504. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Anand, R.; Zhang, X.; Francia, S.; Michelini, F.; Galbiati, A.; Williams, H.; Ronato, D.A.; Masson, J.Y.; Rothenberg, E.; et al. MRE11-RAD50-NBS1 complex is sufficient to promote transcription by RNA polymerase II at double-strand breaks by melting DNA ends. Cell Rep. 2021, 34, 108565. [Google Scholar] [CrossRef]
- Dosio, F.; Arpicco, S.; Stella, B.; Fattal, E. Hyaluronic acid for anticancer drug and nucleic acid delivery. Adv. Drug Deliv. Rev. 2016, 97, 204–236. [Google Scholar] [CrossRef]
- Duan, H.; Liu, Y.; Gao, Z.; Huang, W. Recent advances in drug delivery systems for targeting cancer stem cells. Acta Pharm. Sin. B 2021, 11, 55–70. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CD44 | |||
|---|---|---|---|
| Variable | Low | High | p-Value * |
| n (%) | n (%) | ||
| Age | 41 (46.6) | 47 (53.4) | – |
| years (mean ± SD) | 52.0 ± 13.5 | 52.6 ± 13.2 | 0.830 |
| Gender | |||
| Female | 4 (80.0) | 1 (20.0) | 0.280 |
| Male | 37 (44.6) | 46 (55.4) | |
| Histologic grade | |||
| I | 37 (46.3) | 43 (53.7) | 0.866 |
| II | 4 (50.0) | 4 (50.0) | |
| Tumor size | |||
| T1 + T2 | 27 (43.5) | 35 (56.5) | 0.516 |
| T3 + T4 | 14 (53.8) | 12 (46.2) | |
| Lymph node metastasis | |||
| No | 35 (53.8) | 30 (46.2) | 0.040 |
| Yes | 6 (26.1) | 17 (73.9) | |
| Pathologic stage | |||
| I + II | 23 (45.1) | 28 (54.9) | 0.910 |
| III + IV | 18 (48.6) | 19 (51.4) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, S.-S.F.; Hung, A.C.; Hsu, C.-W.; Lan, T.-H.; Su, C.-W.; Chi, T.-C.; Chang, Y.-C.; Chen, Y.-K.; Wang, Y.-Y. CD44 Mediates Oral Squamous Cell Carcinoma-Promoting Activity of MRE11 via AKT Signaling. J. Pers. Med. 2022, 12, 841. https://doi.org/10.3390/jpm12050841
Yuan S-SF, Hung AC, Hsu C-W, Lan T-H, Su C-W, Chi T-C, Chang Y-C, Chen Y-K, Wang Y-Y. CD44 Mediates Oral Squamous Cell Carcinoma-Promoting Activity of MRE11 via AKT Signaling. Journal of Personalized Medicine. 2022; 12(5):841. https://doi.org/10.3390/jpm12050841
Chicago/Turabian StyleYuan, Shyng-Shiou F., Amos C. Hung, Ching-Wei Hsu, Ting-Hsun Lan, Chang-Wei Su, Tsung-Chen Chi, Yu-Chiuan Chang, Yuk-Kwan Chen, and Yen-Yun Wang. 2022. "CD44 Mediates Oral Squamous Cell Carcinoma-Promoting Activity of MRE11 via AKT Signaling" Journal of Personalized Medicine 12, no. 5: 841. https://doi.org/10.3390/jpm12050841
APA StyleYuan, S. -S. F., Hung, A. C., Hsu, C. -W., Lan, T. -H., Su, C. -W., Chi, T. -C., Chang, Y. -C., Chen, Y. -K., & Wang, Y. -Y. (2022). CD44 Mediates Oral Squamous Cell Carcinoma-Promoting Activity of MRE11 via AKT Signaling. Journal of Personalized Medicine, 12(5), 841. https://doi.org/10.3390/jpm12050841