
Comprehensive Analysis and Experimental Validation of a Novel Estrogen/Progesterone-Related Prognostic Signature for Endometrial Cancer

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
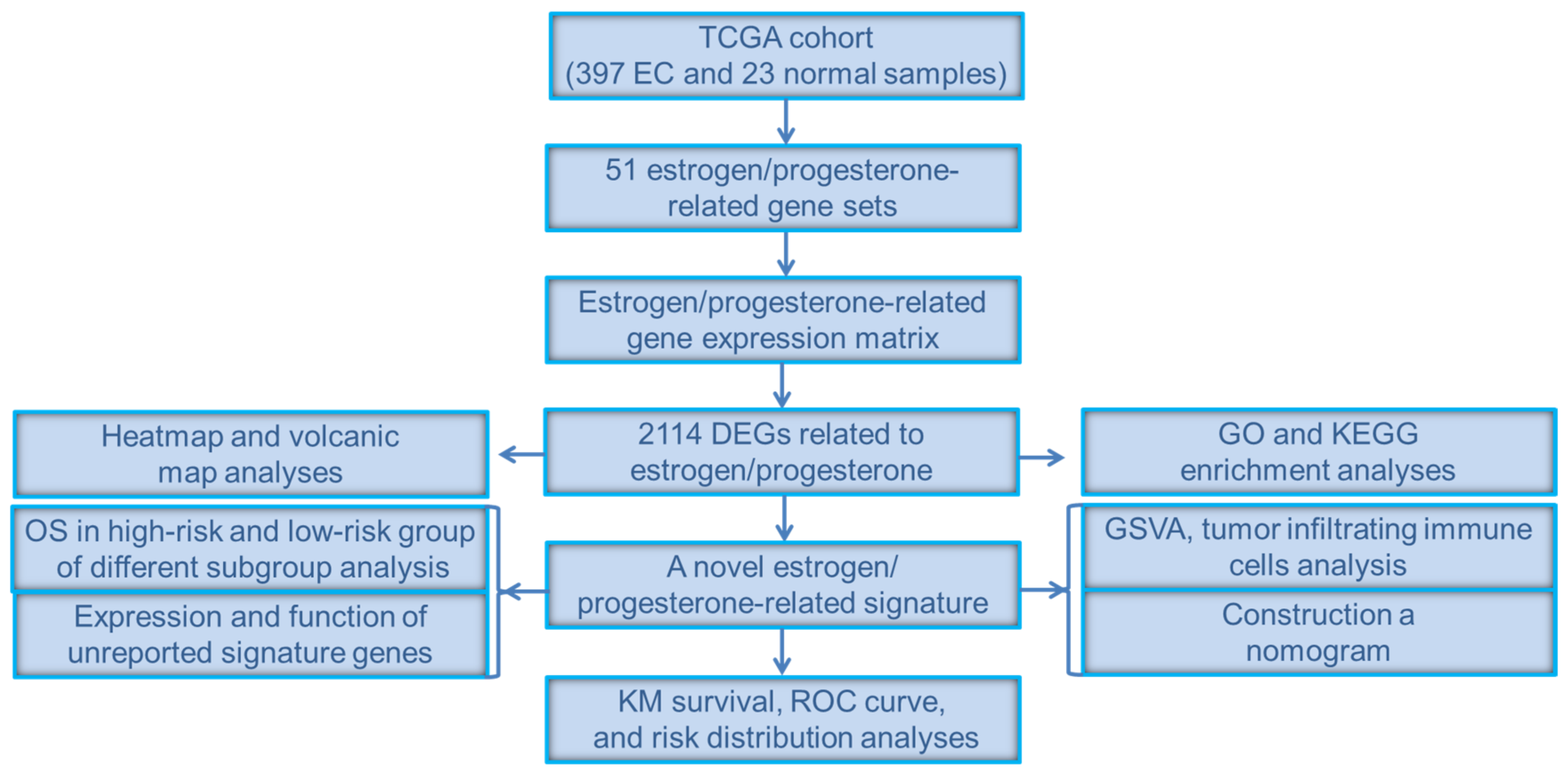
2. Materials and Methods
2.1. Data Extraction and Preprocessing
2.2. Heatmap, Gene Set Enrichment Analysis (GSEA), Gene Ontology (GO) Analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analysis
2.3. Establishment and Validation of a Novel Estrogen/Progesterone Gene Signature
2.4. OS Assessment of the High-Risk and Low-Risk Groups in Different Clinical Subgroups
2.5. Gene Set Variation Analysis (GSVA), Tumor-Infiltrating Immune Cells Analysis and Nomogram Construction
2.6. Clinical Samples
2.7. Total RNA Extraction
2.8. Quantitative Real-Time Polymerase Chain Reaction (qRT–PCR) Assay
2.9. Western Blotting Assay
2.10. Cell Culture, Transfection and Lentiviral Transduction
2.11. Cell Proliferation, Cell Cycle and Colony Formation Assays
2.12. Xenograft Model Assay
2.13. Statistical Analysis
3. Results
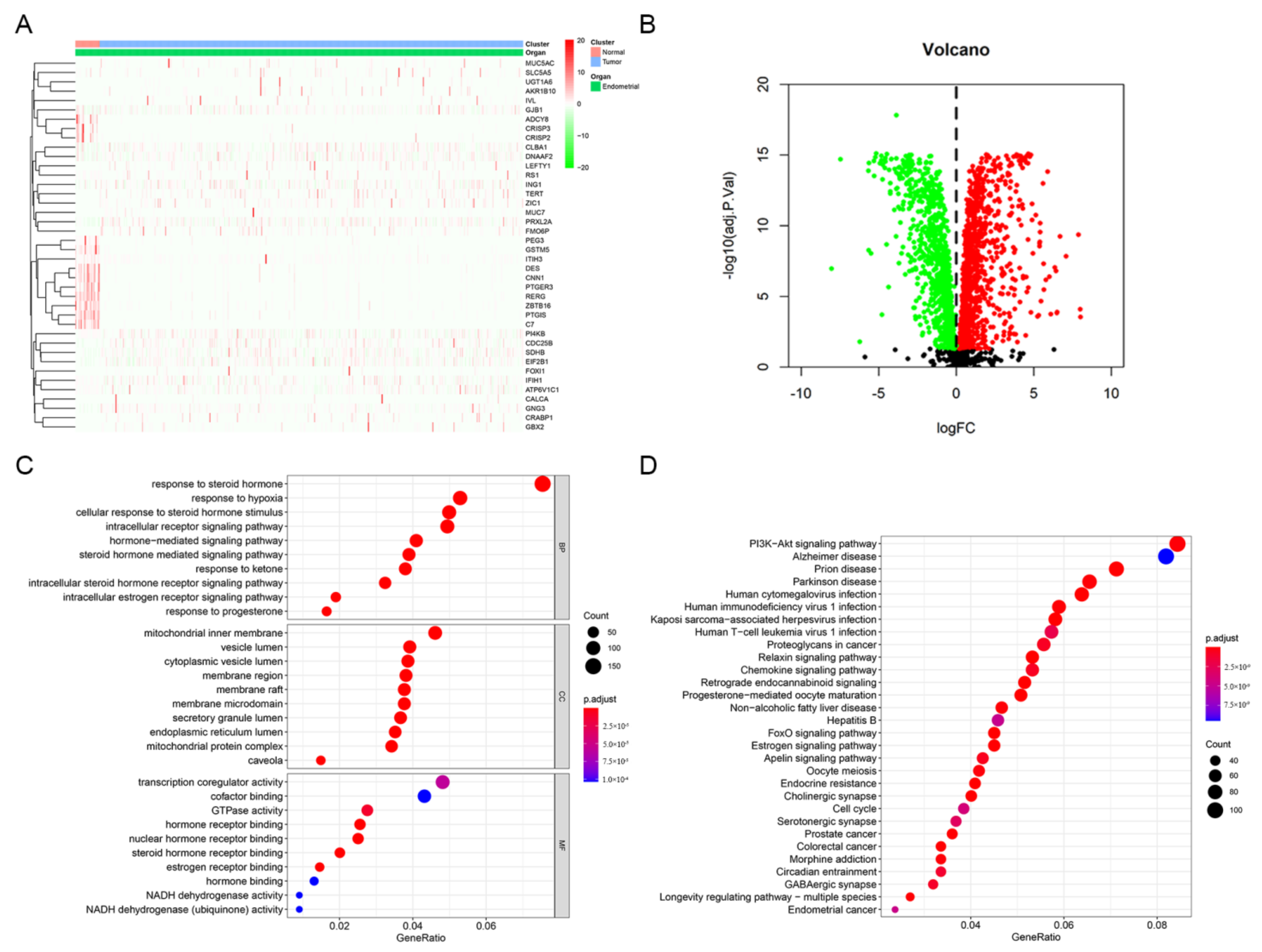
3.1. Identification and Enrichment Analysis of Estrogen/Progesterone-Related DEGs
3.2. Construction of a Signature Incorporating Five Estrogen/Progesterone-Related Genes
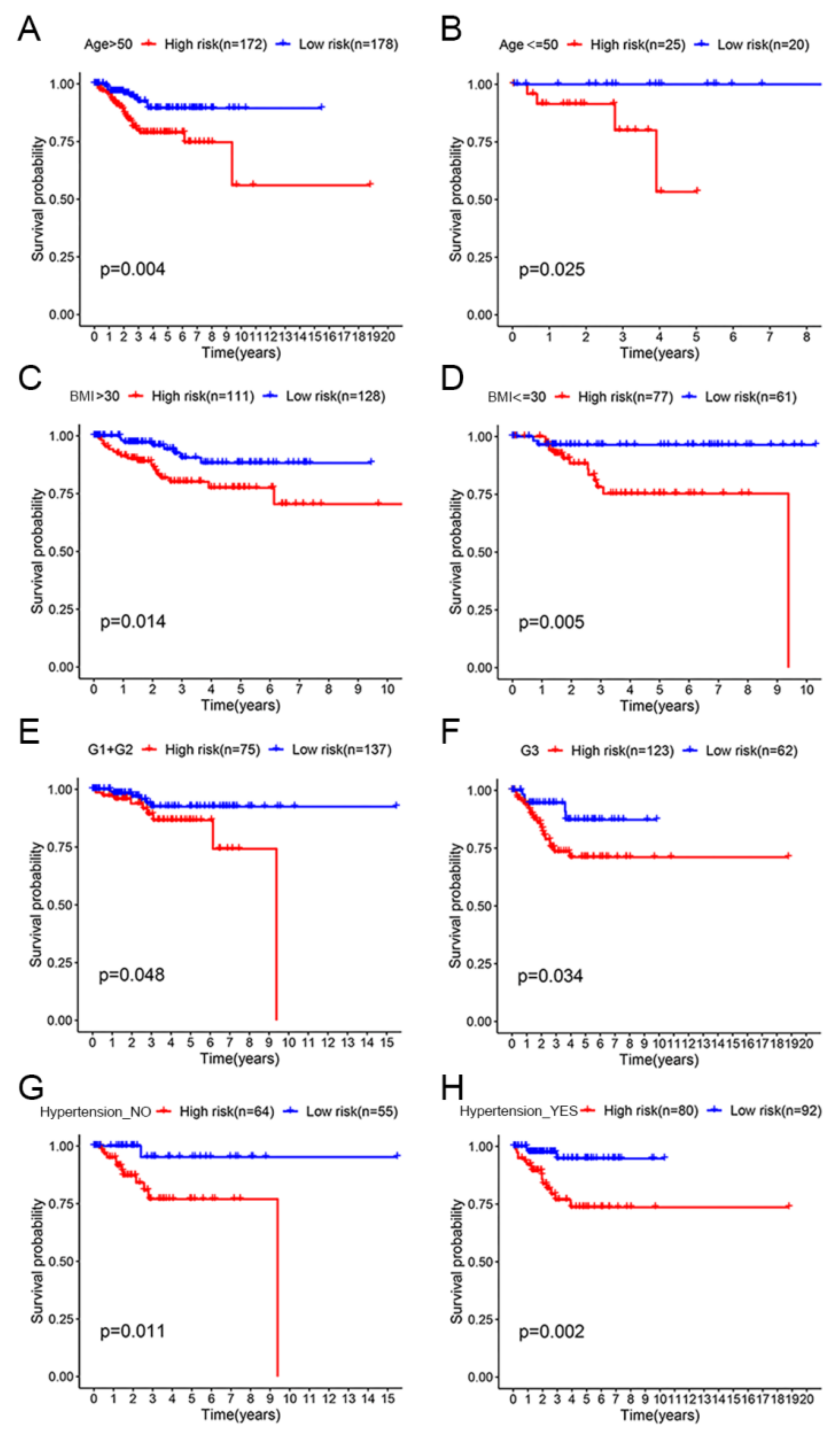
3.3. OS Assessment of the High-Risk and Low-Risk Groups in Different Clinical Subgroups
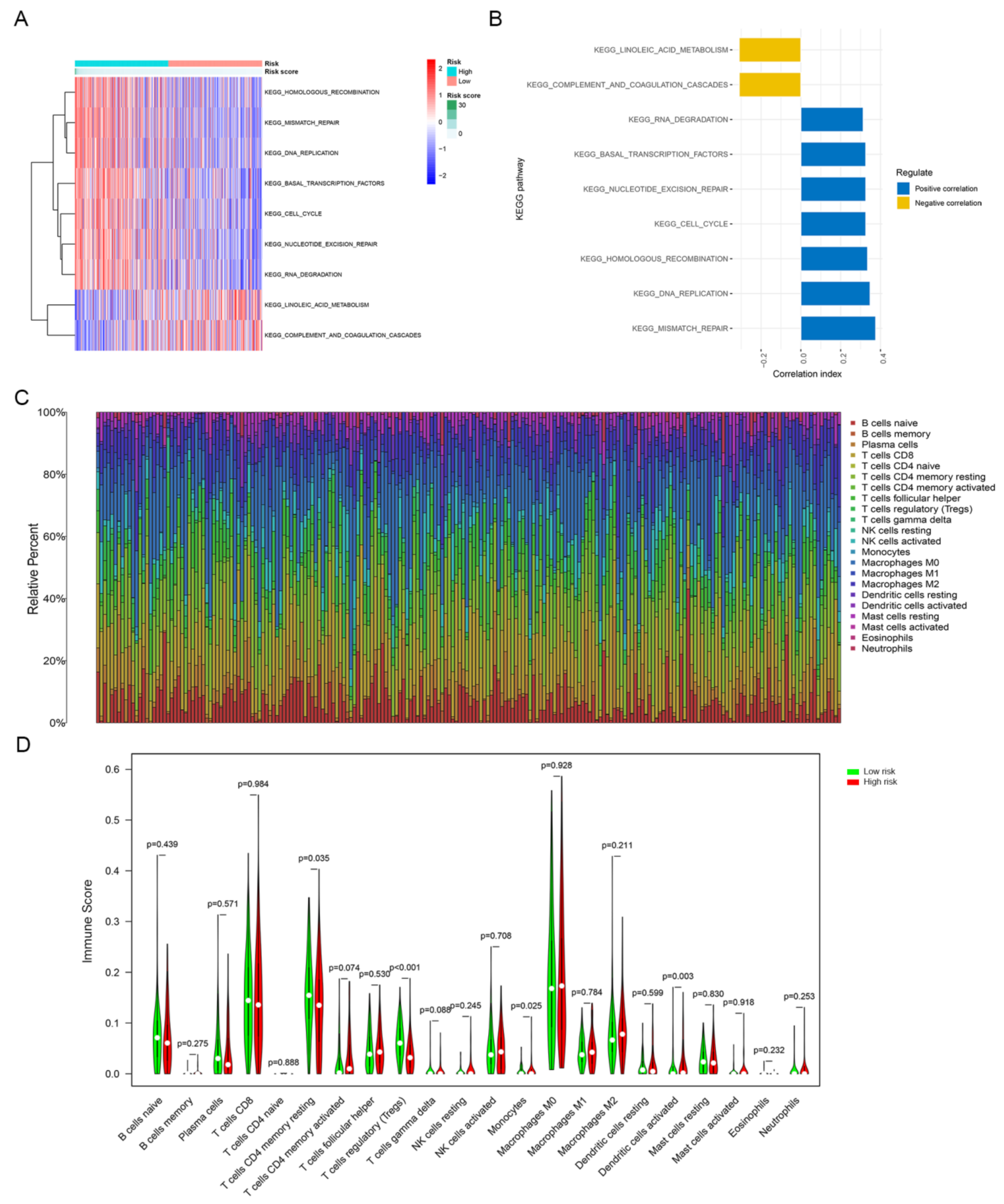
3.4. GSVA and Tumor-Infiltrating Immune Cells Analysis of the Signature between the High-Risk and Low-Risk Groups
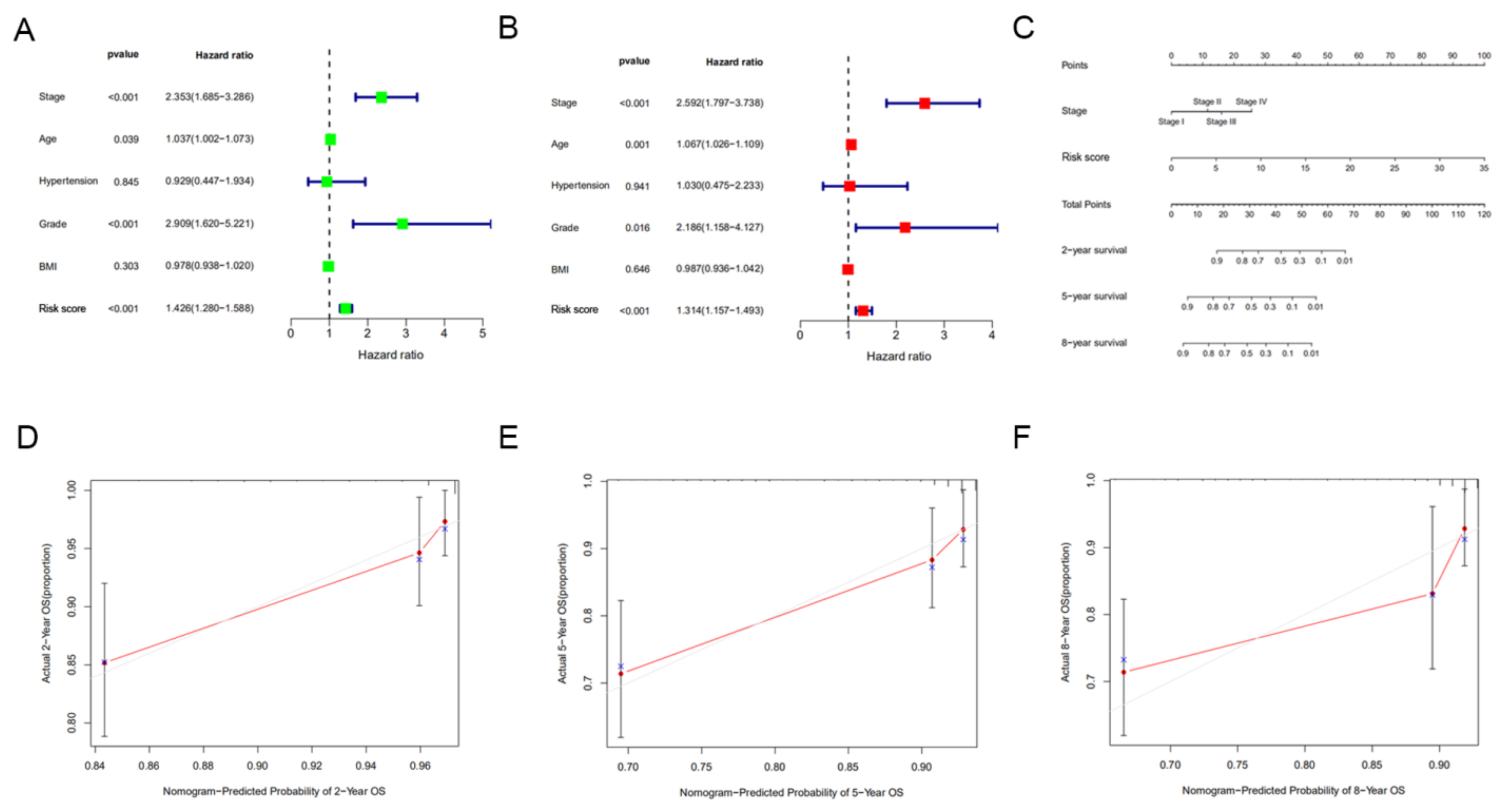
3.5. Construction of a Nomogram Incorporating the Signature for Predicting OS
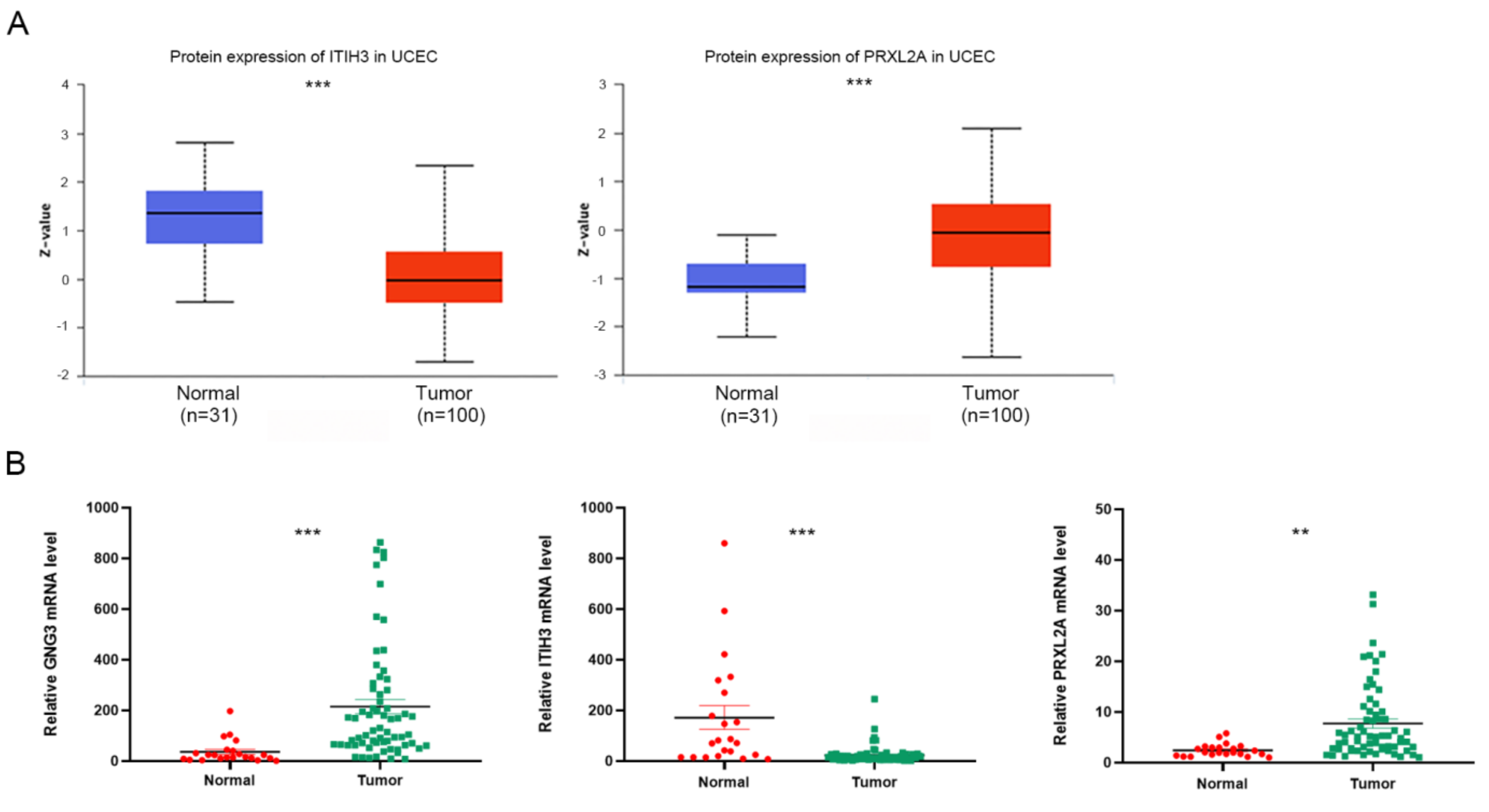
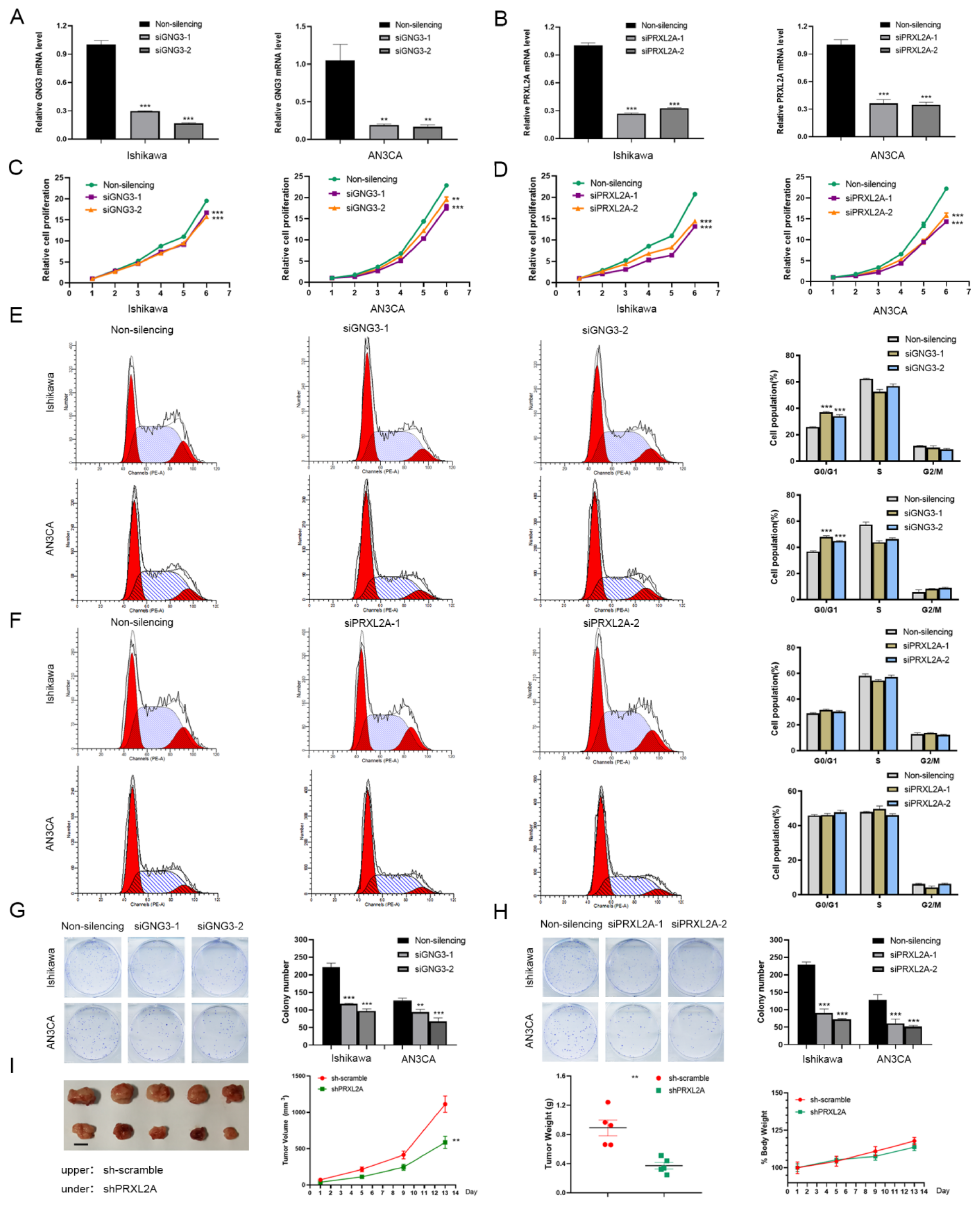
3.6. Expression and Functional Analysis of GNG3, ITIH3 and PRXL2A in EC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Yin, X.H.; Jia, H.Y.; Xue, X.R.; Yang, S.Z.; Wang, Z.Q. Clinical analysis of endometrial cancer patients with obesity, diabetes, and hypertension. Int. J. Clin. Exp. Med. 2014, 7, 736–743. [Google Scholar] [PubMed]
- Weiderpass, E.; Persson, I.; Adami, H.O.; Magnusson, C.; Lindgren, A.; Baron, J.A. Body size in different periods of life, diabetes mellitus, hypertension, and risk of postmenopausal endometrial cancer (Sweden). Cancer Causes Control 2000, 11, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Purdie, D.M.; Green, A.C. Epidemiology of endometrial cancer. Best Pract. Res. Clin. Obstet. Gynaecol. 2001, 15, 341–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunderson, C.C.; Fader, A.N.; Carson, K.A.; Bristow, R.E. Oncologic and reproductive outcomes with progestin therapy in women with endometrial hyperplasia and grade 1 adenocarcinoma: A systematic review. Gynecol. Oncol. 2012, 125, 477–482. [Google Scholar] [CrossRef]
- Zhang, G.; Cheng, Y.; Zhang, Q.; Li, X.; Zhou, J.; Wang, J.; Wei, L. ATXLPA axis facilitates estrogeninduced endometrial cancer cell proliferation via MAPK/ERK signaling pathway. Mol. Med. Rep. 2018, 17, 4245–4252. [Google Scholar]
- Busch, E.L.; Crous-Bou, M.; Prescott, J.; Chen, M.M.; Downing, M.J.; Rosner, B.A.; Mutter, G.L.; De Vivo, I. Endometrial Cancer Risk Factors, Hormone Receptors, and Mortality Prediction. Cancer Epidemiol. Prev. Biomark. 2017, 26, 727–735. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Dai, X.; Chen, L.; Lee, A.C.; Tong, M.; Wise, M.; Chen, Q. Body Mass Index Is Positively Associated with Endometrial Cancer in Chinese Women, Especially Prior to Menopause. J. Cancer 2016, 7, 1169–1173. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.G.; Wu, X.X.; Hua, T.; Xin, X.Y.; Feng, D.L.; Chi, S.Q.; Wang, X.X.; Wang, H.B. NLRP3 inflammasome activation by estrogen promotes the progression of human endometrial cancer. OncoTargets Ther. 2019, 12, 6927–6936. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Lei, H.; Yi, T.; Su, P.; Tang, S.; Tong, Y.; Dong, B.; Ruan, G.; Mustea, A.; Sehouli, J.; et al. Lipid reprogramming induced by the TFEB-ERRalpha axis enhanced membrane fluidity to promote EC progression. J. Exp. Clin. Cancer Res. 2022, 41, 28. [Google Scholar] [CrossRef] [PubMed]
- Miki, Y.; Iwabuchi, E.; Takagi, K.; Suzuki, T.; Sasano, H.; Yaegashi, N.; Ito, K. Co-expression of nuclear heterogeneous nuclear ribonucleic protein K and estrogen receptor alpha in endometrial cancer. Pathol.-Res. Pract. 2022, 231, 153795. [Google Scholar] [CrossRef] [PubMed]
- Poole, A.J.; Li, Y.; Kim, Y.; Lin, S.J.; Lee, W.; Lee, E.Y.P. Prevention of Brca1-mediated mammary tumorigenesis in mice by a progesterone antagonist. Science 2006, 314, 1467–1470. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Kurita, T.; Bulun, S.E. Progesterone action in endometrial cancer, endometriosis, uterine fibroids, and breast cancer. Endocr. Rev. 2013, 34, 130–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.; Li, M.; Shao, K.; Yang, Y.; Wang, Q.; Cai, M. Progesterone induces cell apoptosis via the CACNA2D3/Ca2+/p38 MAPK pathway in endometrial cancer. Oncol. Rep. 2020, 43, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Hanifi-Moghaddam, P.; Hanekamp, E.E.; Kloosterboer, H.J.; Franken, P.; Veldscholte, J.; van Doorn, H.C.; Ewing, P.C.; Kim, J.J.; Grootegoed, J.A.; et al. Progesterone inhibition of Wnt/beta-catenin signaling in normal endometrium and endometrial cancer. Clin. Cancer Res. 2009, 15, 5784–5793. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ma, X.; Wang, Y.; Liu, Y.; Liu, C. Comparison of Different Scoring Systems in the Assessment of Estrogen Receptor Status for Predicting Prognosis in Endometrial Cancer. Int. J. Gynecol. Pathol. 2019, 38, 111–118. [Google Scholar] [CrossRef]
- Van Weelden, W.J.; Reijnen, C.; Kusters-Vandevelde, H.V.N.; Bulten, J.; Bult, P.; Leung, S.; Visser, N.C.M.; Santacana, M.; Bronsert, P.; Hirschfeld, M.; et al. The cutoff for estrogen and progesterone receptor expression in endometrial cancer revisited: A European Network for Individualized Treatment of Endometrial Cancer collaboration study. Hum. Pathol. 2021, 109, 80–91. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research Network; Kandoth, C.; Schultz, N. Integrated genomic characterization of endometrial carcinoma. Nature 2013, 497, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mesirov, J.P. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.Y.; Yu, J.; Liu, T.T.; Yang, K.X.; Yang, L.Y.; Chen, Q.; Shi, F.; Hao, J.J.; Cai, Y.; Wang, M.R.; et al. Plumbagin inhibits the proliferation and survival of esophageal cancer cells by blocking STAT3-PLK1-AKT signaling. Cell Death Dis. 2018, 9, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, X.L.; Jiang, T.; Zhao, W.B.; Wang, F.; Wang, G.L.; Cui, M.; Wen, Z.Q. Gene alterations in tumor-associated endothelial cells from endometrial cancer. Int. J. Mol. Med. 2008, 22, 619–632. [Google Scholar] [PubMed]
- Xie, D.; Liang, Y.; Su, Y.; An, Y.; Pengpeng, Q. miR-152 inhibits proliferation of human endometrial cancer cells via inducing G2/M phase arrest by suppressing CDC25B expression. Biomed Pharm. 2018, 99, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Yang, H.; Chang, K.; Zhang, B.; Xie, F.; Ye, J.; Chang, R.; Qiu, X.; Wang, Y.; Qu, Y.; et al. Melatonin alleviates progression of uterine endometrial cancer by suppressing estrogen/ubiquitin C/SDHB-mediated succinate accumulation. Cancer Lett. 2020, 476, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Dou, Y.; Kawaler, E.A.; Zhou, D.C.; Gritsenko, M.A.; Huang, C.; Blumenberg, L.; Karpova, A.; Petyuk, V.A.; Savage, S.R.; Satpathy, S.; et al. Proteogenomic Characterization of Endometrial Carcinoma. Cell 2020, 180, 729–748. [Google Scholar] [CrossRef]
- Yu, S.; Sun, Z.; Zong, L.; Yan, J.; Yu, M.; Chen, J.; Lu, Z. Clinicopathological and molecular characterization of high-grade endometrial carcinoma with POLE mutation: A single center study. J. Gynecol. Oncol. 2022, 33, 38. [Google Scholar] [CrossRef]
- Hause, R.J.; Pritchard, C.C.; Shendure, J.; Salipantel, S.J. Classification and characterization of microsatellite instability across 18 cancer types. Nat. Med. 2016, 22, 1342–1350. [Google Scholar] [CrossRef]
- Oaknin, A.; Gilbert, L.; Tinker, A.V.; Brown, J.; Mathews, C.; Press, J.; Sabatier, R.; O’Malley, D.M.; Samouelian, V.; Boni, V.; et al. Safety and antitumor activity of dostarlimab in patients with advanced or recurrent DNA mismatch repair deficient/microsatellite instability-high (dMMR/MSI-H) or proficient/stable (MMRp/MSS) endometrial cancer: Interim results from GARNET-a phase I, single-arm study. J. Immunother. Cancer 2022, 10, 003777. [Google Scholar]
- Marabelle, A.; Le, D.T.; Ascierto, P.A.; di Giacomo, A.M.; de Jesus-Acosta, A.; Delord, J.; Geva, R.; Gottfried, M.; Penel, N.; Hansen, A.R.; et al. Efficacy of Pembrolizumab in Patients With Noncolorectal High Microsatellite Instability/Mismatch Repair-Deficient Cancer: Results From the Phase II KEYNOTE-158 Study. J. Clin. Oncol. 2020, 38, 1. [Google Scholar] [CrossRef]
- Concin, N.; Matias-Guiu, X.; Vergote, I.; Cibula, D.; Mirza, M.R.; Marnitz, S.; Ledermann, J.; Bosse, T.; Chargari, C.; Fagotti, A.; et al. ESGO/ESTRO/ESP guidelines for the management of patients with endometrial carcinoma. Int. J. Gynecol. Cancer 2021, 31, 12–39. [Google Scholar] [CrossRef]
- Rodriguez, A.C.; Blanchard, Z.; Maurer, K.A.; Gertz, J. Estrogen Signaling in Endometrial Cancer: A Key Oncogenic Pathway with Several Open Questions. Horm. Cancer 2019, 10, 51–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.F.; Yang, C.C.; Kao, S.Y.; Liu, C.J.; Lin, S.C.; Chang, K.W. MicroRNA-211 Enhances the Oncogenicity of Carcinogen-Induced Oral Carcinoma by Repressing TCF12 and Increasing Antioxidant Activity. Cancer Res. 2016, 76, 4872–4886. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Yang, L.; Zhang, X.; Chen, R.; Chen, X.; Tang, W.; Zhang, M. Identification of Potential Biomarkers in Glioblastoma through Bioinformatic Analysis and Evaluating Their Prognostic Value. BioMed. Res. Int. 2019, 2019, 6581576. [Google Scholar] [CrossRef]
- Cardaci, S.; Zheng, L.; MacKay, G.; van den Broek, N.J.F.; MacKenzie, E.D.; Nixon, C.; Stevenson, D.; Tumanov, S.; Bulusu, V.; Kamphorst, J.J.; et al. Pyruvate carboxylation enables growth of SDH-deficient cells by supporting aspartate biosynthesis. Nat. Cell Biol. 2015, 17, 1317–1326. [Google Scholar] [CrossRef] [PubMed]
- Andrews, K.A.; Ascher, D.B.; Pires, D.E.V.; Barnes, D.R.; Vialard, L.; Casey, R.T.; Bradshaw, N.; Adlard, J.; Aylwin, S.; Brennan, P.; et al. Tumour risks and genotype-phenotype correlations associated with germline variants in succinate dehydrogenase subunit genes SDHB, SDHC and SDHD. J. Med. Genet. 2018, 55, 384–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, J.; Li, H.; Chu, J.; Sheng, J.; Wang, C.; Jia, Z.; Meng, W.; Yin, H.; Wan, J.; He, F. LncRNA FAM83A-AS1 promotes ESCC progression by regulating miR-214/CDC25B axis. J. Cancer 2021, 12, 1200–1211. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Zeng, B.H.; Xu, L.H.; Wang, J.Y.; Li, M.Z.; Zeng, M.S.; Liu, W.L. Anti-CDC25B autoantibody predicts poor prognosis in patients with advanced esophageal squamous cell carcinoma. J. Transl. Med. 2010, 8, 81. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Chua, M.S.; He, J.; So, S.K. Small interfering RNA targeting CDC25B inhibits liver tumor growth in vitro and in vivo. Mol. Cancer 2008, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Yu, Y.; Gao, D.; Jin, W.; Jiang, P.; Li, Y.; Wang, C.; Song, Y.; Zhan, P.; Gu, F.; et al. Inhibition of CDC25B With WG-391D Impedes the Tumorigenesis of Ovarian Cancer. Front. Oncol. 2019, 9, 236. [Google Scholar] [CrossRef]
- Cerchia, C.; Nasso, R.; Mori, M.; Villa, S.; Gelain, A.; Capasso, A.; Aliotta, F.; Simonetti, M.; Rullo, R.; Masullo, M.; et al. Discovery of Novel Naphthylphenylketone and Naphthylphenylamine Derivatives as Cell Division Cycle 25B (CDC25B) Phosphatase Inhibitors: Design, Synthesis, Inhibition Mechanism, and in Vitro Efficacy against Melanoma Cell Lines. J. Med. Chem. 2019, 62, 7089–7110. [Google Scholar] [CrossRef]
- George Rosenker, K.M.; Paquette, W.D.; Johnston, P.A.; Sharlow, E.R.; Vogt, A.; Bakan, A.; Lazo, J.S.; Wipf, P. Synthesis and biological evaluation of 3-aminoisoquinolin-1(2H)-one based inhibitors of the dual-specificity phosphatase Cdc25B. Bioorg. Med. Chem. 2015, 23, 2810–2818. [Google Scholar] [CrossRef]
- Boutros, R.; Mondesert, O.; Lorenzo, C.; Astuti, P.; McArthur, G.; Chircop, M.; Ducommun, B.; Gabrielli, B. CDC25B overexpression stabilises centrin 2 and promotes the formation of excess centriolar foci. PLoS ONE 2013, 8, 67822. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.Q.; Liu, Z.; Ngan, E.S.; Tsai, S.Y. Cdc25B functions as a novel coactivator for the steroid receptors. Mol. Cell Biol. 2001, 21, 8056–8067. [Google Scholar] [CrossRef] [Green Version]
- Chua, S.S.; Ma, Z.; Ngan, E.; Tsai, S.Y. Cdc25B as a steroid receptor coactivator. Vitam. Horm. 2004, 68, 231–256. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Yao, H.-W.; Liu, T.-T.; Wang, D.; Shi, J.-H.; Yuan, G.-W.; Ma, S.; Wu, L.-Y. Comprehensive Analysis and Experimental Validation of a Novel Estrogen/Progesterone-Related Prognostic Signature for Endometrial Cancer. J. Pers. Med. 2022, 12, 914. https://doi.org/10.3390/jpm12060914
Yu J, Yao H-W, Liu T-T, Wang D, Shi J-H, Yuan G-W, Ma S, Wu L-Y. Comprehensive Analysis and Experimental Validation of a Novel Estrogen/Progesterone-Related Prognostic Signature for Endometrial Cancer. Journal of Personalized Medicine. 2022; 12(6):914. https://doi.org/10.3390/jpm12060914
Chicago/Turabian StyleYu, Jing, Hong-Wen Yao, Ting-Ting Liu, Di Wang, Jian-Hong Shi, Guang-Wen Yuan, Sai Ma, and Ling-Ying Wu. 2022. "Comprehensive Analysis and Experimental Validation of a Novel Estrogen/Progesterone-Related Prognostic Signature for Endometrial Cancer" Journal of Personalized Medicine 12, no. 6: 914. https://doi.org/10.3390/jpm12060914
APA StyleYu, J., Yao, H. -W., Liu, T. -T., Wang, D., Shi, J. -H., Yuan, G. -W., Ma, S., & Wu, L. -Y. (2022). Comprehensive Analysis and Experimental Validation of a Novel Estrogen/Progesterone-Related Prognostic Signature for Endometrial Cancer. Journal of Personalized Medicine, 12(6), 914. https://doi.org/10.3390/jpm12060914