
Functionally Significant Variants in Genes Associated with Abdominal Obesity: A Review


Abstract
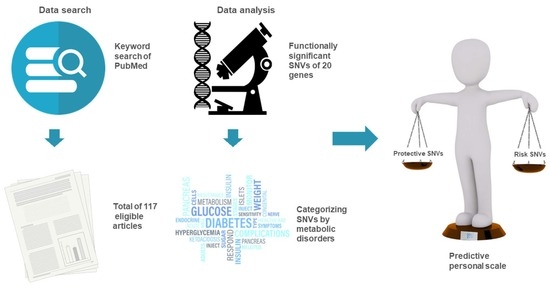
:
1. Introduction
2. Results
2.1. ADIPOQ
2.2. ADRB3
2.3. APLN and APLNR
2.4. CCL2 and CCL7
2.5. FTO
2.6. GCG and GLP1R
2.7. GHRL
2.8. GIP
2.9. INS
2.10. LEP
2.11. NAMPT
2.12. PYY and PPY
2.13. RBP4
2.14. RETN
2.15. SCT
2.16. UCP2
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caballero, B. Humans against Obesity: Who Will Win? Adv. Nutr. 2019, 10 (Suppl. S1), S4–S9. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Li, H. Obesity: Epidemiology, Pathophysiology, and Therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef]
- Ling, C.; Rönn, T. Epigenetics in Human Obesity and Type 2 Diabetes. Cell Metab. 2019, 9, 1028–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, B.M.; Lindgren, C.M. The genetics of obesity. Curr. Diab. Rep. 2010, 10, 498–505. [Google Scholar] [CrossRef] [Green Version]
- Vettori, A.; Pompucci, G.; Paolini, B.; Del Ciondolo, I.; Bressan, S.; Dundar, M.; Kenanoğlu, S.; Unfer, V.; Bertelli, M.; Geneob Project. Genetic background, nutrition and obesity: A review. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1751–1761. [Google Scholar] [CrossRef]
- Hurtado, A.M.D.; Acosta, A. Precision Medicine and Obesity. Gastroenterol. Clin. N. Am. 2021, 50, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Kumar, P.; Mahalingam, K. Molecular Genetics of Human Obesity: A Comprehensive Review. C R Biol. 2017, 340, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Hebebrand, J.; Hinney, A.; Knoll, N.; Volckmar, A.L.; Scherag, A. Molecular genetic aspects of weight regulation. Dtsch. Arztebl. Int. 2013, 110, 338–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, C.; Garcia-Vargas, G.G.; Morales, R.P. Monogenic, Polygenic and Multifactorial Obesity in Children: Genetic and Environmental Factor. Austin J. Nutr. Metab. 2017, 4, 1052. [Google Scholar]
- Littleton, S.H.; Berkowitz, R.I.; Grant, S.F.A. Genetic Determinants of Childhood Obesity. Mol. Diagn. Ther. 2020, 24, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, R.; Kimonis, V.; Butler, M.G. Genetics of Obesity in Humans: A Clinical Review. Int. J. Mol. Sci. 2022, 23, 11005. [Google Scholar] [CrossRef] [PubMed]
- GBD 2015 Obesity Collaborators. Health effects of overweight and obesity in 195 countries over 25 years. N. Engl. J. Med. 2017, 377, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Loos, R.J.F.; Yeo, G.S.H. The Genetics of Obesity: From Discovery to Biology. Nat. Rev. Genet. 2022, 23, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Heianza, Y.; Qi, L. Gene-Diet Interaction and Precision Nutrition in Obesity. Int. J. Mol. Sci. 2017, 18, 787. [Google Scholar] [CrossRef] [Green Version]
- Litwin, M.; Kułaga, Z. Obesity, metabolic syndrome, and primary hypertension. Pediatr. Nephrol. 2021, 36, 825–837. [Google Scholar] [CrossRef]
- Ahmed, B.; Sultana, R.; Greene, M.W. Adipose tissue and insulin resistance in obese. Biomed. Pharmacother. 2021, 137, 111315. [Google Scholar] [CrossRef]
- Brunner, K.T.; Henneberg, C.J.; Wilechansky, R.M.; Long, M.T. Nonalcoholic Fatty Liver Disease and Obesity Treatment. Curr. Obes. Rep. 2019, 8, 220–228. [Google Scholar] [CrossRef]
- Milić, S.; Lulić, D.; Štimac, D. Non-alcoholic fatty liver disease and obesity: Biochemical, metabolic and clinical presentations. World J. Gastroenterol. 2014, 20, 9330–9337. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, J.; Tang, X.; Luo, Q.; Xu, D.; Yu, B. Interaction between adipocytes and high-density lipoprotein:new insights into the mechanism of obesity-induced dyslipidemia and atherosclerosis. Lipids Health Dis. 2019, 18, 223. [Google Scholar] [CrossRef] [Green Version]
- Yan, B.; Yang, J.; Zhao, B.; Wu, Y.; Bai, L.; Ma, X. Causal Effect of Visceral Adipose Tissue Accumulation on the Human Longevity: A Mendelian Randomization Study. Front Endocrinol. 2021, 1, 722187. [Google Scholar] [CrossRef]
- Cao, H. Adipocytokines in obesity and metabolic disease. J. Endocrinol. 2014, 220, 47–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maximus, P.S.; Al Achkar, Z.; Hamid, P.F.; Hasnain, S.S.; Peralta, C.A. Adipocytokines: Are they the Theory of Everything? Cytokine 2020, 133, 155144. [Google Scholar] [CrossRef]
- Zorena, K.; Jachimowicz-Duda, O.; Ślęzak, D.; Robakowska, M.; Mrugacz, M. Adipokines and Obesity. Potential Link to Metabolic Disorders and Chronic Complications. Int. J. Mol. Sci. 2020, 21, 3570. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Kato, N.; Hwang, J.Y.; Guo, X.; Tabara, Y.; Li, H.; Dorajoo, R.; Yang, X.; Tsai, F.J.; Li, S. Genome-wide association studies in East Asians identify new loci for waist-hip ratio and waist circumference. Sci. Rep. 2016, 20, 17958. [Google Scholar] [CrossRef] [PubMed]
- Dedov, I.I.; Shestakova, M.V.; Melnichenko, G.A.; Mazurina, N.V.; Andreeva, E.N.; Bondarenko, I.Z.; Gusova, Z.R.; Dzgoeva, F.K.; Eliseev, M.S.; Ershova, E.V.; et al. Interdisciplinary Clinical Practice Guidelines “Management of obesity and its comorbidities”. Obes. Metab. 2021, 18, 5–99. [Google Scholar] [CrossRef]
- Yumuk, V.; Tsigos, C.; Fried, M.; Schindler, K.; Busetto, L.; Micic, D.; Toplak, H. Obesity Management Task Force of the European Association for the Study of Obesity. European Guidelines for Obesity Management in Adults. Obes. Facts 2015, 8, 402–424. [Google Scholar] [CrossRef]
- Serra-Juhé, C.; Martos-Moreno, G.; De Pieri, F.B.; Flores, R.; Chowen, J.A.; Pérez-Jurado, L.A.; Argente, J. Heterozygous rare genetic variants in non-syndromic early-onset obesity. Int. J. Obes. 2020, 44, 830–841. [Google Scholar] [CrossRef] [Green Version]
- Ghoshal, K.; Chatterjee, T.; Chowdhury, S.; Sengupta, S.; Bhattacharyya, M. Adiponectin Genetic Variant and Expression Coupled with Lipid Peroxidation Reveal New Signatures in Diabetic Dyslipidemia. Biochem. Genet. 2021, 59, 781–798. [Google Scholar] [CrossRef]
- Iwabu, M.; Okada-Iwabu, M.; Yamauchi, T.; Kadowaki, T. Adiponectin/AdipoR Research and Its Implications for Lifestyle-Related Diseases. Front. Cardiovasc. Med. 2019, 6, 116. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Zhang, T.; Wang, S.; He, Y.; Ye, Q.; Cao, W. The Trp64Arg Polymorphism in Β3 Adrenergic Receptor (ADRB3) Gene Is Associated with Adipokines and Plasma Lipids: A Systematic Review, Meta-Analysis, and Meta-Regression. Lipids Health Dis. 2020, 19, 99. [Google Scholar] [CrossRef]
- Li, C.; Cheng, H.; Adhikari, B.K.; Wang, S.; Yang, N.; Liu, W.; Sun, J.; Wang, Y. The Role of Apelin-APJ System in Diabetes and Obesity. Front. Endocrinol. 2022, 13, 820002. [Google Scholar] [CrossRef]
- Ignacio, R.M.; Gibbs, C.R.; Lee, E.S.; Son, D.S. Differential Chemokine Signature between Human Preadipocytes and Adipocytes. Immune Netw. 2016, 16, 189–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claussnitzer, M.; Dankel, S.N.; Kim, K.-H.; Quon, G.; Meuleman, W.; Haugen, C.; Glunk, V.; Sousa, I.S.; Beaudry, J.L.; Puviindran, V.; et al. FTO Obesity Variant Circuitry and Adipocyte Browning in Humans. N. Engl. J. Med. 2015, 373, 895–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Zhang, M.; Wang, J.J.; Wang, C.J.; Ren, Y.C.; Wang, B.Y.; Zhang, H.Y.; Yang, X.Y.; Zhao, Y.; Han, C.Y.; et al. Association of TCF7L2 and GCG Gene Variants with Insulin Secretion, Insulin Resistance, and Obesity in New-Onset Diabetes. Biomed. Environ. Sci. 2016, 29, 814–817. [Google Scholar] [CrossRef] [PubMed]
- Mayendraraj, A.; Rosenkilde, M.M.; Gasbjerg, L.S. GLP-1 and GIP Receptor Signaling in Beta Cells—A Review of Receptor Interactions and Co-Stimulation. Peptides 2022, 151, 170749. [Google Scholar] [CrossRef] [PubMed]
- Landgren, S.; Simms, J.A.; Thelle, D.S.; Strandhagen, E.; Bartlett, S.E.; Engel, J.A.; Jerlhag, E. The Ghrelin Signalling System Is Involved in the Consumption of Sweets. PLoS ONE 2011, 6, e18170. [Google Scholar] [CrossRef] [Green Version]
- Nauck, M.A.; Quast, D.R.; Wefers, J.; Pfeiffer, A.F.H. The evolving story of incretins (GIP and GLP-1) in metabolic and cardiovascular disease: A pathophysiological update. Diabetes Obes. Metab. 2021, 23 (Suppl. S3), 5–29. [Google Scholar] [CrossRef] [PubMed]
- Støy, J.; Edghill, E.L.; Flanagan, S.E.; Ye, H.; Paz, V.P.; Pluzhnikov, A.; Below, J.E.; Hayes, M.G.; Cox, N.J.; Lipkind, G.M.; et al. Insulin gene mutations as a cause of permanent neonatal diabetes. Proc. Natl. Acad. Sci. USA 2007, 104, 15040–15044. [Google Scholar] [CrossRef] [Green Version]
- Wabitsch, M.; Pridzun, L.; Ranke, M.; von Schnurbein, J.; Moss, A.; Brandt, S.; Kohlsdorf, K.; Moepps, B.; Schaab, M.; Funcke, J.B.; et al. Measurement of immunofunctional leptin to detect and monitor patients with functional leptin deficiency. Eur. J. Endocrinol. 2017, 176, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Curat, C.A.; Wegner, V.; Sengenes, C.; Miranville, A.; Tonus, C.; Busse, R.; Bouloumie, A. Macrophages in human visceral adipose tissue: Increased accumulation in obesity and a source of resistin and visfatin. Diabetologia 2006, 49, 744–747. [Google Scholar] [CrossRef] [Green Version]
- Stromsdorfer, K.L.; Yamaguchi, S.; Yoon, M.J.; Moseley, A.C.; Franczyk, M.P.; Kelly, S.C.; Qi, N.; Imai, S.I.; Yoshino, J. NAMPT-Mediated NAD+ Biosynthesis in Adipocytes Regulates Adipose Tissue Function and Multi-Organ Insulin Sensitivity in Mice. Cell Rep. 2016, 16, 1851–1860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, C.D.; Lyon, H.N.; Nemesh, J.; Drake, J.A.; Tuomi, T.; Gaudet, D.; Zhu, X.; Cooper, R.S.; Ardlie, K.G.; Groop, L.C.; et al. Association studies of BMI and type 2 diabetes in the neuropeptide Y pathway: A possible role for NPY2R as a candidate gene for type 2 diabetes in men. Diabetes 2007, 56, 1460–14607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaner, W.S. Vitamin A signaling and homeostasis in obesity, diabetes, and metabolic disorders. Pharmacol. Ther. 2019, 197, 153–178. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.; Menzaghi, C.; Smith, S.; Liang, L.; de Rekeneire, N.; Garcia, M.E.; Lohman, K.K.; Miljkovic, I.; Strotmeyer, E.S.; Cummings, S.R.; et al. Genome-wide association analysis identifies TYW3/CRYZ and NDST4 loci associated with circulating resistin levels. Hum. Mol. Genet. 2012, 21, 4774–4780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathwa, N.; Patel, R.; Palit, S.P.; Ramachandran, A.V.; Begum, R. Genetic variants of resistin and its plasma levels: Association with obesity and dyslipidemia related to type 2 diabetes susceptibility. Genomics 2019, 111, 980–985. [Google Scholar] [CrossRef]
- Schnabl, K.; Li, Y.; Klingenspor, M. The gut hormone secretin triggers a gut-brown fat-brain axis in the control of food intake. Exp. Physiol. 2020, 105, 1206–1213. [Google Scholar] [CrossRef]
- Oliveira, M.S.; Rheinheimer, J.; Moehlecke, M.; Rodrigues, M.; Assmann, T.S.; Leitão, C.B.; Trindade, M.R.M.; Crispim, D.; de Souza, B.M. UCP2, IL18, and miR-133a-3p are dysregulated in subcutaneous adipose tissue of patients with obesity. Mol. Cell. Endocrinol. 2020, 509, 110805. [Google Scholar] [CrossRef]
- Available online: https://pubmed.ncbi.nlm.nih.gov (accessed on 14 November 2022).
- Khoramipour, K.; Chamari, K.; Hekmatikar, A.A.; Ziyaiyan, A.; Taherkhani, S.; Elguindy, N.M.; Bragazzi, N.L. Adiponectin: Structure, Physiological Functions, Role in Diseases, and Effects of Nutrition. Nutrients 2021, 13, 1180. [Google Scholar] [CrossRef]
- Pruitt, K.D.; Tatusova, T.; Brown, G.R.; Maglott, D.R. NCBI Reference Sequences (RefSeq): Current Status, New Features and Genome Annotation Policy. Nucleic. Acids Res. 2012, 40, D130–D135. [Google Scholar] [CrossRef] [Green Version]
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef] [Green Version]
- Bueno, A.C.; Sun, K.; Martins, C.S.; Elias, J.; Miranda, W.; Tao, C.; Foss-Freitas, M.C.; Barbieri, M.A.; Bettiol, H.; de Castro, M.; et al. A Novel ADIPOQ Mutation (p.M40K) Impairs Assembly of High-Molecular-Weight Adiponectin and Is Associated with Early-Onset Obesity and Metabolic Syndrome. J. Clin. Endocrinol. Metab. 2014, 99, E683–E693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apalasamy, Y.D.; Rampal, S.; Salim, A.; Moy, F.M.; Bulgiba, A.; Mohamed, Z. Association of ADIPOQ Gene with Obesity and Adiponectin Levels in Malaysian Malays. Mol. Biol. Rep. 2014, 41, 2917–2921. [Google Scholar] [CrossRef] [PubMed]
- Olusegun, E.; Ogundele, O.E.; Adekoya, K.O.; Osinubi, A.A.A.; Awofala, A.A.; Oboh, B.O. Association of Adiponectin Gene (ADIPOQ) Polymorphisms with Measures of Obesity in Nigerian Young Adults. Egypt. J. Med. Hum. Genet. 2018, 19, 123–127. [Google Scholar] [CrossRef]
- Siitonen, N.; Pulkkinen, L.; Lindström, J.; Kolehmainen, M.; Eriksson, J.G.; Venojärvi, M.; Ilanne-Parikka, P.; Keinänen-Kiukaanniemi, S.; Tuomilehto, J.; Uusitupa, M. Association of ADIPOQ Gene Variants with Body Weight, Type 2 Diabetes and Serum Adiponectin Concentrations: The Finnish Diabetes Prevention Study. BMC Med. Genet. 2011, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Palit, S.P.; Patel, R.; Jadeja, S.D.; Rathwa, N.; Mahajan, A.; Ramachandran, A.V.; Dhar, M.K.; Sharma, S.; Begum, R. A Genetic Analysis Identifies a Haplotype at Adiponectin Locus: Association with Obesity and Type 2 Diabetes. Sci. Rep. 2020, 10, 2904. [Google Scholar] [CrossRef] [Green Version]
- Vasseur, F.; Helbecque, N.; Dina, C.; Lobbens, S.; Delannoy, V.; Gaget, S.; Boutin, P.; Vaxillaire, M.; Leprêtre, F.; Dupont, S.; et al. Single-Nucleotide Polymorphism Haplotypes in the Both Proximal Promoter and Exon 3 of the APM1 Gene Modulate Adipocyte-Secreted Adiponectin Hormone Levels and Contribute to the Genetic Risk for Type 2 Diabetes in French Caucasians. Hum. Mol. Genet. 2002, 11, 2607–2614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, H.; Shimomura, L.; Matsukawa, Y.; Kumada, M.; Takahashi, M.; Matsuda, M.; Ouchi, N.; Kihara, S.; Kawamoto, T.; Sumitsuji, S.; et al. Association of Adiponectin Mutation With Type 2 Diabetes A Candidate Gene for the Insulin Resistance Syndrome. Diabetes 2002, 51, 2325–2328. [Google Scholar] [CrossRef] [Green Version]
- Hammad, S.S.; Eck, P.; Sihag, J.; Chen, X.; Connelly, P.W.; Lamarche, B.; Couture, P.; Guay, V.; Maltais-Giguère, J.; West, S.G.; et al. Common Variants in Lipid Metabolism-Related1 Genes Associate with Fat Mass Changes in Response to Dietary Monounsaturated Fatty Acids in Adults with Abdominal Obesity. J. Nutr. 2019, 149, 1749–1756. [Google Scholar] [CrossRef]
- ADRB3 Adrenoceptor Beta 3 [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/155 (accessed on 14 November 2022).
- Takenaka, A.; Nakamura, S.; Mitsunaga, F.; Inoue-Murayama, M.; Udono, T.; Suryobroto, B. Human-specific SNP in obesity genes, adrenergic receptor beta2 (ADRB2), Beta3 (ADRB3), and PPAR γ2 (PPARG), during primate evolution. PLoS ONE 2012, 7, e43461. [Google Scholar] [CrossRef] [Green Version]
- Kadowaki, H.; Yasuda, K.; Iwamoto, K.; Otabe, S.; Shimokawa, K.; Silver, K.; Walston, J.; Yoshinaga, H.; Kosaka, K.; Yamada, N.; et al. A Mutation in the Β3-Adrenergic Receptor Gene Is Associated with Obesity and Hyperinsulinemia in Japanese Subjects. Biochem. Biophys. Res. Commun 1995, 215, 555–560. [Google Scholar] [CrossRef]
- Xie, C.; Hua, W.; Zhao, Y.; Rui, J.; Feng, J.; Chen, Y.; Liu, Y.; Liu, J.; Yang, X.; Xu, X. The ADRB3 Rs4994 Polymorphism Increases Risk of Childhood and Adolescent Overweight/Obesity for East Asia’s Population: An Evidence-Based Meta-Analysis. Adipocyte 2020, 9, 77–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryukm, J.A.; Zhang, X.; Ko, B.S.; Daily, J.W.; Park, S. Association of β3-adrenergic receptor rs4994 polymorphisms with the risk of type 2 diabetes: A systematic review and meta-analysis. Diabetes Res. Clin. Pract. 2017, 129, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Yang, T.L.; Tang, B.S.; Chen, X.; Huang, X.; Luo, X.H.; Zhu, Y.S.; Chen, X.P.; Hu, P.C.; Chen, J.; et al. Two Novel Functional Single Nucleotide Polymorphisms of ADRB3 Are Associated with Type 2 Diabetes in the Chinese Population. J. Clin. Endocrinol. Metab. 2013, 98, E1272–E1277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, J.C.; Zhang, Z.Z.; Wang, W.; McKinnie, S.M.K.; Vederas, J.C.; Oudit, G.Y. Targeting the Apelin Pathway as a Novel Therapeutic Approach for Cardiovascular Diseases. Biochim. Biophys. Acta Mol. Basis. Dis. 2017, 1863, 1942–1950. [Google Scholar] [CrossRef]
- Zheng, H.; Fan, X.; Li, X.; Zhang, Y.; Fan, Y.; Zhang, N.; Song, Y.; Ren, F.; Shen, C.; Shen, J.; et al. The Association between Single Nucleotide Polymorphisms of the Apelin Gene and Diabetes Mellitus in a Chinese Population. J. Pediatr. Endocrinol. Metab. 2016, 29, 1397–1402. [Google Scholar] [CrossRef]
- Liao, Y.C.; Chou, W.W.; Li, Y.N.; Chuang, S.C.; Lin, W.Y.; Lakkakula, B.V.K.S.; Yu, M.L.; Juo, S.H.H. Apelin gene polymorphism influences apelin expression and obesity phenotypes in Chinese women. Am. J. Clin. Nutr. 2011, 94, 921–928. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Peng, F.; Lin, L.; Yu, M.; Huang, C.; Hu, D.; Guo, Q.; Xu, C.; Lin, J.; Zhang, M.; et al. Association Study of Apelin-APJ System Genetic Polymorphisms with Incident Metabolic Syndrome in a Chinese Population: A Case-Control Study. Oncotarget 2019, 10, 3807–3817. [Google Scholar] [CrossRef]
- Galon-Tilleman, H.; Yang, H.; Bednarek, M.A.; Spurlock, S.M.; Paavola, K.J.; Ko, B.; To, C.; Luo, J.; Tian, H.; Jermutus, L.; et al. Apelin-36 Modulates Blood Glucose and Body Weight Independently of Canonical APJ Receptor Signaling. J. Biol. Chem. 2017, 292, 1925–1933. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, M.; Asaba, K.; Nakayama, T. The APLNR gene polymorphism rs7119375 is associated with an increased risk of development of essential hypertension in the Chinese population: A meta-analysis. Medicine 2020, 99, e22418. [Google Scholar] [CrossRef]
- Available online: https://www.ncbi.nlm.nih.gov/gene/ (accessed on 14 November 2022).
- Mikhailova, S.V.; Ivanoshchuk, D.E. Innate-Immunity Genes in Obesity. J. Pers. Med. 2021, 11, 1201. [Google Scholar] [CrossRef]
- Guzmán-Ornelas, M.O.; Petri, M.H.; Vázquez-Del Mercado, M.; Chavarría-Ávila, E.; Corona-Meraz, F.I.; Ruíz-Quezada, S.L.; Madrigal-Ruíz, P.M.; Castro-Albarrán, J.; Sandoval-García, F.; Navarro-Hernández, R.E. CCL2 Serum Levels and Adiposity Are Associated with the Polymorphic Phenotypes -2518A on CCL2 and 64ILE on CCR2 in a Mexican Population with Insulin Resistance. J. Diabetes Res. 2016, 2016, 5675739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teler, J.; Tarnowski, M.; Safranow, K.; Maciejewska, A.; Sawczuk, M.; Dziedziejko, V.; Sluczanowska-Glabowska, S.; Pawlik, A. CCL2, CCL5, IL4 and IL15 Gene Polymorphisms in Women with Gestational Diabetes Mellitus. Horm. Metab. Res. 2017, 49, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Huber, J.; Kiefer, F.W.; Zeyda, M.; Ludvik, B.; Silberhumer, G.R.; Prager, G.; Zlabinger, G.J.; Stulnig, T.M. CC Chemokine and CC Chemokine Receptor Profiles in Visceral and Subcutaneous Adipose Tissue Are Altered in Human Obesity. J. Clin. Endocrinol. Metab. 2008, 93, 3215–3221. [Google Scholar] [CrossRef] [PubMed]
- Sindhu, S.; Thomas, R.; Kochumon, S.; Wilson, A.; Abu-Farha, M.; Bennakhi, A.; Al-Mulla, F.; Ahmad, R. Increased Adipose Tissue Expression of Interferon Regulatory Factor (IRF)-5 in Obesity: Association with Metabolic Inflammation. Cells 2019, 8, 1418. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Liu, F.; Lu, Z.; Fei, Q.; Ai, Y.; He, P.C.; Shi, H.; Cui, X.; Su, R.; Klungland, A.; et al. Differential m 6 A, m 6 A m, and m 1 A Demethylation Mediated by FTO in the Cell Nucleus and Cytoplasm. Mol. Cell 2018, 71, 973–985.e5. [Google Scholar] [CrossRef] [Green Version]
- Povel, C.M.; Boer, J.M.; Onland-Moret, N.C.; Dollé, M.E.; Feskens, E.J.; van der Schouw, Y.T. Single nucleotide polymorphisms (SNPs) involved in insulin resistance, weight regulation, lipid metabolism and inflammation in relation to metabolic syndrome: An epidemiological study. Cardiovasc. Diabetol. 2012, 11, 133. [Google Scholar] [CrossRef] [Green Version]
- Hinney, A.; Nguyen, T.T.; Scherag, A.; Friedel, S.; Brönner, G.; Müller, T.D.; Grallert, H.; Illig, T.; Wichmann, H.E.; Rief, W.; et al. Genome Wide Association (GWA) Study for Early Onset Extreme Obesity Supports the Role of Fat Mass and Obesity Associated Gene (FTO) Variants. PLoS ONE 2007, 2, e1361. [Google Scholar] [CrossRef] [Green Version]
- Scott, L.J.; Mohlke, K.L.; Bonnycastle, L.L.; Willer, C.J.; Li, Y.; Duren, W.L.; Erdos, M.R.; Stringham, H.M.; Chines, P.S.; Jackson, A.U.; et al. A Genome-Wide Association Study of Type 2 Diabetes in Finns Detects Multiple Susceptibility Variants. Science 2007, 316, 1341–1345. [Google Scholar] [CrossRef] [Green Version]
- Larsson, S.C.; Burgess, S.; Michaëlsson, K. Genetic association between adiposity and gout: A Mendelian randomization study. Rheumatology 2018, 57, 2145–2148. [Google Scholar] [CrossRef]
- Fox, C.S.; Liu, Y.; White, C.C.; Feitosa, M.; Smith, A.V.; Heard-Costa, N.; Lohman, K.; GIANT Consortium; MAGIC Consortium; GLGC Consortium; et al. Genome-wide association for abdominal subcutaneous and visceral adipose reveals a novel locus for visceral fat in women. PLoS Genet. 2012, 8, e1002695. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, G.; Tabassum, R.; Mahajan, A.; Dwivedi, O.P.; Mahendran, Y.; Kaur, I.; Nigam, S.; Dubey, H.; Varma, B.; Madhu, S.V.; et al. Common Variants of FTO and the Risk of Obesity and Type 2 Diabetes in Indians. J. Hum. Genet. 2011, 56, 720–726. [Google Scholar] [CrossRef] [Green Version]
- Baturin, A.K.; Sorokina, E.Y.; Pogozheva, A.V.; Keshabyants, E.E.; Kobelkova, I.v.; Kambarov, A.O.; Elizarova, E.v.; Tutelyan, V.A. The Association of Rs993609 Polymorphisms of Gene FTO and Rs659366 Polymorphisms of Gene UCP2 with Obesity among Arctic Russian Population. Vopr. Pitan. 2017, 86, 32–39. [Google Scholar] [CrossRef]
- Matsuo, T.; Nakata, Y.; Hotta, K.; Tanaka, K. The FTO genotype as a useful predictor of body weight maintenance: Initial data from a 5-year follow-up study. Metabolism 2014, 63, 912–917. [Google Scholar] [CrossRef] [Green Version]
- Kamura, Y.; Iwata, M.; Maeda, S.; Shinmura, S.; Koshimizu, Y.; Honoki, H.; Fukuda, K.; Ishiki, M.; Usui, I.; Fukushima, Y.; et al. FTO Gene Polymorphism Is Associated with Type 2 Diabetes through Its Effect on Increasing the Maximum BMI in Japanese Men. PLoS ONE 2016, 11, e0165523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cyrus, C.; Ismail, M.H.; Chathoth, S.; Vatte, C.; Hasen, M.; al Ali, A. Analysis of the Impact of Common Polymorphisms of the FTO and MC4R Genes with the Risk of Severe Obesity in Saudi Arabian Population. Genet. Test Mol. Biomark. 2018, 22, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Xi, B.; Cheng, H.; Shen, Y.; Chandak, G.R.; Zhao, X.; Hou, D.; Wu, L.; Wang, X.; Mi, J. Study of 11 BMI-associated loci identified in GWAS for associations with central obesity in the Chinese children. PLoS ONE 2013, 8, e56472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salim, S.; Kartawidjajaputra, F.; Suwanto, A. Association of FTO rs9939609 and CD36 rs1761667 with Visceral Obesity. J. Nutr. Sci. Vitam. 2020, 66, S329–S335. [Google Scholar] [CrossRef] [PubMed]
- Kring, S.I.; Holst, C.; Zimmermann, E.; Jess, T.; Berentzen, T.; Toubro, S.; Hansen, T.; Astrup, A.; Pedersen, O.; Sørensen, T.I. FTO gene associated fatness in relation to body fat distribution and metabolic traits throughout a broad range of fatness. PLoS ONE 2008, 3, e2958. [Google Scholar] [CrossRef] [PubMed]
- Ağagündüz, D.; Gezmen-Karadağ, M. Association of FTO common variant (rs9939609) with body fat in Turkish individuals. Lipids Health Dis. 2019, 18, 212. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.; Xing, X.; Zhang, X.; Hong, J.; Yang, W. Sexual Dimorphism of a Genetic Risk Score for Obesity and Related Traits among Chinese Patients with Type 2 Diabetes. Obes. Facts 2019, 12, 328–343. [Google Scholar] [CrossRef]
- Moore, S.C.; Gunter, M.J.; Daniel, C.R.; Reddy, K.S.; George, P.S.; Yurgalevitch, S.; Devasenapathy, N.; Ramakrishnan, L.; Chatterjee, N.; Chanock, S.J.; et al. Common genetic variants and central adiposity among Asian-Indians. Obesity 2012, 20, 1902–1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monnereau, C.; Santos, S.; van der Lugt, A.; Jaddoe, V.W.V.; Felix, J.F. Associations of adult genetic risk scores for adiposity with childhood abdominal, liver and pericardial fat assessed by magnetic resonance imaging. Int. J. Obes. 2018, 42, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Monnereau, C.; Vogelezang, S.; Kruithof, C.J.; Jaddoe, V.W.; Felix, J.F. Associations of genetic risk scores based on adult adiposity pathways with childhood growth and adiposity measures. BMC Genet. 2016, 17, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkler, T.W.; Justice, A.E.; Graff, M.; Barata, L.; Feitosa, M.F.; Chu, S.; Czajkowski, J.; Esko, T.; Fall, T.; Kilpeläinen, T.O.; et al. The Influence of Age and Sex on Genetic Associations with Adult Body Size and Shape: A Large-Scale Genome-Wide Interaction Study. PLoS Genet. 2015, 11, e1005378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GCG Glucagon [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/2641 (accessed on 14 November 2022).
- Torekov, S.S.; Ma, L.; Grarup, N.; Hartmann, B.; Hainerová, I.A.; Kielgast, U.; Kissow, H.; Rosenkilde, M.; Lebl, J.; Witte, D.R.; et al. Homozygous Carriers of the G Allele of Rs4664447 of the Glucagon Gene (GCG) Are Characterised by Decreased Fasting and Stimulated Levels of Insulin, Glucagon and Glucagon-like Peptide (GLP)-1. Diabetologia 2011, 54, 2820–2831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GLP1R Glucagon Like Peptide 1 Receptor [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/2740 (accessed on 14 November 2022).
- Tokuyama, Y.; Matsui, K.; Egashira, T.; Nozaki, O.; Ishizuka, T.; Kanatsuka, A. Five Missense Mutations in Glucagon-like Peptide 1 Receptor Gene in Japanese Population. Diabetes Res. Clin. Pract. 2004, 66, 63–69. [Google Scholar] [CrossRef]
- Li, P.; Tiwari, H.K.; Lin, W.Y.; Allison, D.B.; Chung, W.K.; Leibel, R.L.; Yi, N.; Liu, N. Genetic Association Analysis of 30 Genes Related to Obesity in a European American Population. Int. J. Obes. 2014, 38, 724–729. [Google Scholar] [CrossRef] [Green Version]
- de Luis, D.A.; Aller, R.; de la Fuente, B.; Primo, D.; Conde, R.; Izaola, O.; Sagrado, M.G. Relation of the Rs6923761 Gene Variant in Glucagon-like Peptide 1 Receptor with Weight, Cardiovascular Risk Factor, and Serum Adipokine Levels in Obese Female Subjects. J. Clin. Lab. Anal. 2015, 29, 100–105. [Google Scholar] [CrossRef]
- Michałowska, J.; Miller-Kasprzak, E.; Seraszek-Jaros, A.; Mostowska, A.; Bogdański, P. Association of GLP1R Variants Rs2268641 and Rs6923761 with Obesity and Other Metabolic Parameters in a Polish Cohort. Front. Endocrinol. 2022, 13, 1000185. [Google Scholar] [CrossRef]
- Wessel, J.; Chu, A.Y.; Willems, S.M.; Wang, S.; Yaghootkar, H.; Brody, J.A.; Dauriz, M.; Hivert, M.F.; Raghavan, S.; Lipovich, L.; et al. Low-Frequency and Rare Exome Chip Variants Associate with Fasting Glucose and Type 2 Diabetes Susceptibility. Nat. Commun. 2015, 6, 5897. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Li, P.; Li, R.; Yu, Z.; Sun, X.; Ji, G.; Yang, X.; Zhu, L.; Zhu, S. GLP1R Single-Nucleotide Polymorphisms Rs3765467 and Rs10305492 Affect β Cell Insulin Secretory Capacity and Apoptosis through GLP-1. DNA Cell Biol. 2020, 39, 1700–1710. [Google Scholar] [CrossRef] [PubMed]
- El Eid, L.; Reynolds, C.A.; Tomas, A.; Jones, B. Biased Agonism and Polymorphic Variation at the GLP-1 Receptor: Implications for the Development of Personalised Therapeutics. Pharmacol. Res. 2022, 184, 106411. [Google Scholar] [CrossRef] [PubMed]
- GHRL Ghrelin and Obestatin Prepropeptide [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/51738 (accessed on 14 November 2022).
- Jiao, Z.T.; Luo, Q. Molecular Mechanisms and Health Benefits of Ghrelin: A Narrative Review. Nutrients 2022, 14, 4191. [Google Scholar] [CrossRef] [PubMed]
- Ukkola, O.; Ravussin, E.; Jacobson, P.; Snyder, E.E.; Chagnon, M.; SjÖstrÖm, L.; Bouchard, C. RAPID COMMUNICATIONS: Mutations in the Preproghrelin/Ghrelin Gene Associated with Obesity in Humans. J. Clin. Endocrinol. Metab. 2001, 86, 3996–3999. [Google Scholar] [CrossRef]
- Steinle, N.I.; Pollin, T.I.; O’Connell, J.R.; Mitchell, B.D.; Shuldiner, A.R. Variants in the Ghrelin Gene Are Associated with Metabolic Syndrome in the Old Order Amish. J. Clin. Endocrinol. Metab. 2005, 90, 6672–6677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gueorguiev, M.; Lecoeur, C.; Meyre, D.; Benzinou, M.; Mein, C.A.; Hinney, A.; Vatin, V.; Weill, J.; Heude, B.; Hebebrand, J.; et al. Association Studies on Ghrelin and Ghrelin Receptor Gene Polymorphisms with Obesity. Obesity 2009, 17, 745–754. [Google Scholar] [CrossRef]
- Chung, W.K.; Patki, A.; Matsuoka, N.; Boyer, B.B.; Liu, N.; Musani, S.K.; Goropashnaya, A.V.; Tan, P.L.; Katsanis, N.; Johnson, S.B.; et al. Analysis of 30 Genes (355 SNPS) Related to Energy Homeostasis for Association with Adiposity in European-American and Yup’ik Eskimo Populations. Hum. Hered. 2009, 67, 193–205. [Google Scholar] [CrossRef]
- Imaizumi, T.; Ando, M.; Nakatochi, M.; Yasuda, Y.; Honda, H.; Kuwatsuka, Y.; Kato, S.; Kondo, T.; Iwata, M.; Nakashima, T.; et al. Effect of Dietary Energy and Polymorphisms in BRAP and GHRL on Obesity and Metabolic Traits. Obes. Res. Clin. Pract. 2018, 12, 39–48. [Google Scholar] [CrossRef]
- Wang, X.; Qu, F.; Wang, C.; Wang, Y.; Wang, D.; Zhao, M.; Yun, X.; Zheng, Q.; Xu, L. Variation Analysis of Ghrelin Gene in Chinese Patients with Obesity, Having Polycystic Ovarian Syndrome. Gynecol. Endocrinol. 2020, 36, 594–598. [Google Scholar] [CrossRef]
- Ando, T.; Komaki, G.; Naruo, T.; Okabe, K.; Takii, M.; Kawai, K.; Konjiki, F.; Takei, M.; Oka, T.; Takeuchi, K.; et al. Possible Role of Preproghrelin Gene Polymorphisms in Susceptibility to Bulimia Nervosa. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2006, 141, 929–934. [Google Scholar] [CrossRef]
- GIP Gastric Inhibitory Polypeptide [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/2695 (accessed on 14 November 2022).
- Chang, C.L.; Cai, J.J.; Cheng, P.J.; Chueh, H.Y.; Hsu, S.Y.T. Identification of Metabolic Modifiers That Underlie Phenotypic Variations in Energy-Balance Regulation. Diabetes 2011, 60, 726–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, K.; Watanabe, K.; Boonvisut, S.; Makishima, S.; Miyashita, H.; Iwamoto, S. Common variants of GIP are associated with visceral fat accumulation in Japanese adults. Am. J. Physiol. Gastrointest. Liver. Physiol. 2014, 307, G1108–G1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- INS Insulin [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/3630 (accessed on 14 November 2022).
- Liu, M.; Weiss, M.A.; Arunagiri, A.; Yong, J.; Rege, N.; Sun, J.; Haataja, L.; Kaufman, R.J.; Arvan, P. Biosynthesis, Structure, and Folding of the Insulin Precursor Protein. Diabetes Obes. Metab. 2018, 20, 28–50. [Google Scholar] [CrossRef] [Green Version]
- Vakilian, M.; Tahamtani, Y.; Ghaedi, K. A Review on Insulin Trafficking and Exocytosis. Gene 2019, 706, 52–61. [Google Scholar] [CrossRef]
- Ghosh, S.; Mahalanobish, S.; Sil, P.C. Diabetes: Discovery of Insulin, Genetic, Epigenetic and Viral Infection Mediated Regulation. Nucleus 2021, 65, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Sun, J.; Cui, J.; Chen, W.; Guo, H.; Barbetti, F.; Arvan, P. INS-Gene Mutations: From Genetics and Beta Cell Biology to Clinical Disease. Mol. Aspects Med. 2015, 42, 3–18. [Google Scholar] [CrossRef] [Green Version]
- Arneth, B. Insulin Gene Mutations and Posttranslational and Translocation Defects: Associations with Diabetes. Endocrine 2020, 70, 488–497. [Google Scholar] [CrossRef]
- Edghill, E.L.; Flanagan, S.E.; Patch, A.M.; Boustred, C.; Parrish, A.; Shields, B.; Shepherd, M.H.; Hussain, K.; Kapoor, R.R.; Malecki, M.; et al. Insulin Mutation Screening in 1044 Patients with Diabetes Mutations in the INS Gene Are a Common Cause of Neonatal Diabetes but a Rare Cause of Diabetes Diagnosed in Childhood or Adulthood. Diabetes 2008, 57, 1034–1042. [Google Scholar] [CrossRef] [Green Version]
- Boesgaard, T.W.; Pruhova, S.; Andersson, E.A.; Cinek, O.; Obermannova, B.; Lauenborg, J.; Damm, P.; Bergholdt, R.; Pociot, F.; Pisinger, C.; et al. Further Evidence That Mutations in INS Can Be a Rare Cause of Maturity-Onset Diabetes of the Young (MODY). BMC Med. Genet. 2010, 11, 42. [Google Scholar] [CrossRef] [Green Version]
- LEP Leptin [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/3952 (accessed on 23 December 2022).
- Caldeira, R.S.; Panissa, V.L.G.; Inoue, D.S.; Campos, E.Z.; Monteiro, P.A.; Giglio, B.D.M.; Pimentel, G.D.; Hofmann, P.; Lira, F.S. Impact to short-term high intensity intermittent training on different storages of body fat, leptin and soluble leptin receptor levels in physically active non-obese men: A pilot investigation. Clin. Nutr. ESPEN 2018, 28, 186–192. [Google Scholar] [CrossRef]
- Hamilton, K.; Harvey, J. The neuronal actions of leptin and the implications for treating alzheimer’s disease. Pharmaceuticals 2021, 14, 52. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, A.; Hernández-Coronado, C.G.; Rosales-Torres, A.M.; Hernández-Medrano, J.H. Leptin regulates neuropeptides associated with food intake and GnRH secretion. Ann. Endocrinol. 2019, 80, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Marcos, P.; Coveñas, R. Neuropeptidergic control of feeding: Focus on the galanin family of peptides. Int. J. Mol. Sci. 2021, 22, 2544. [Google Scholar] [CrossRef] [PubMed]
- P41159 LEP_HUMAN. Available online: https://www.uniprot.org/uniprotkb/P41159/entry (accessed on 23 December 2022).
- Eikelis, N.; Lambert, G.; Wiesner, G.; Kaye, D.; Schlaich, M.; Morris, M.; Hastings, J.; Socratous, F.; Esler, M. Extra-adipocyte leptin release in human obesity and its relation to sympathoadrenal function. Am. J. Physiol. Endocrinol. Metab 2004, 286, E744–E752. [Google Scholar] [CrossRef]
- Funcke, J.B.; von Schnurbein, J.; Lennerz, B.; Lahr, G.; Debatin, K.M.; Fischer-Posovszky, P.; Wabitsch, M. Monogenic forms of childhood obesity due to mutations in the leptin gene. Mol. Cell. Pediatr. 2014, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, S.; Salman, M.; Siddalingaiah, L.B.; Lakshmi, G.L.; Xaviour, D.; Sreenath, J. Genetic variants in leptin: Determinants of obesity and leptin levels in South Indian population. Adipocyte 2014, 4, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Aljanabi, M.A.; Alfaqih, M.A.; Khanfar, M.; Amarin, Z.O.; Elsalem, L.; Saadeh, R.; Al-Mughales, F. Leptin and the GA genotype of rs2167270 of the LEP gene increase the risk of prediabetes. Biomed. Rep. 2021, 14, 44. [Google Scholar] [CrossRef]
- Lombard, Z.; Crowther, N.J.; van der Merwe, L.; Pitamber, P.; Norris, S.A.; Ramsay, M. Appetite regulation genes are associated with body mass index in black South African adolescents: A genetic association study. BMJ Open. 2012, 2, e000873. [Google Scholar] [CrossRef] [Green Version]
- Manju, S.K.; Anilkumar, T.R.; Vysakh, G.; Leena, B.K.; Lekshminarayan, V.; Kumar, P.G.; Shenoy, T.K. A Case-Control Study of the Association of Leptin Gene Polymorphisms with Plasma Leptin Levels and Obesity in the Kerala Population. J. Obes. 2022, 2022, 1040650. [Google Scholar] [CrossRef]
- Ashraf, R.; Khan, M.; Lone, S.; Bhat, M.; Rashid, S.; Majid, S.; Bashir, H. Implication of Leptin and Leptin Receptor Gene Variations in Type 2 Diabetes Mellitus: A Case-Control Study. J. Endocrinol. Metab. 2022, 12, 19–31. [Google Scholar] [CrossRef]
- Enns, J.E.; Taylor, C.G.; Zahradka, P. Variations in Adipokine Genes AdipoQ, Lep, and LepR are Associated with Risk for Obesity-Related Metabolic Disease: The Modulatory Role of Gene-Nutrient Interactions. J. Obes. 2011, 2011, 168659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaghootkar, H.; Zhang, Y.; Spracklen, C.N.; Karaderi, T.; Huang, L.O.; Bradfield, J.; Schurmann, C.; Fine, R.S.; Preuss, M.H.; Kutalik, Z.; et al. Genetic Studies of Leptin Concentrations Implicate Leptin in the Regulation of Early Adiposity. Diabetes 2020, 69, 2806–2818. [Google Scholar] [CrossRef] [PubMed]
- Haglund, E.; Nguyen, L.; Schafer, N.P.; Lammert, H.; Jennings, P.A.; Onuchic, J.N. Uncovering the molecular mechanisms behind disease-associated leptin variants. J. Biol. Chem. 2018, 293, 12919–12933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NAMPT Nicotinamide Phosphoribosyltransferase [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/10135 (accessed on 14 November 2022).
- Fagerberg, L.; Hallstrom, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the Human Tissue-Specific Expression by Genome-Wide Integration of Transcriptomics and Antibody-Based Proteomics. Mol. Cell. Proteom. 2014, 13, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Böttcher, Y.; Teupser, D.; Enigk, B.; Berndt, J.; Klöting, N.; Schön, M.R.; Thiery, J.; Blüher, M.; Stumvoll, M.; Kovacs, P. Genetic Variation in the Visfatin Gene (PBEF1) and Its Relation to Glucose Metabolism and Fat-Depot-Specific Messenger Ribonucleic Acid Expression in Humans. J. Clin. Endocrinol. Metab. 2006, 91, 2725–2731. [Google Scholar] [CrossRef]
- Johansson, L.M.; Johansson, L.E.; Ridderstråle, M. The Visfatin (PBEF1) G-948T Gene Polymorphism Is Associated with Increased High-Density Lipoprotein Cholesterol in Obese Subjects. Metabolism 2008, 57, 1558–1562. [Google Scholar] [CrossRef]
- Blakemore, A.I.F.; Meyre, D.; Delplanque, J.; Vatin, V.; Lecoeur, C.; Marre, M.; Tichet, J.; Balkau, B.; Froguel, P.; Walley, A.J. A Rare Variant in the Visfatin Gene (Nampt/Pbef1) Is Associated with Protection from Obesity. Obesity 2009, 17, 1549–1553. [Google Scholar] [CrossRef]
- Tabassum, R.; Mahendran, Y.; Dwivedi, O.P.; Chauhan, G.; Ghosh, S.; Marwaha, R.K.; Tandon, N.; Bharadwaj, D. Common Variants of IL6, LEPR, and PBEF1 Are Associated with Obesity in Indian Children. Diabetes 2012, 61, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Rong, J.; Chu, M.; Xing, B.; Zhu, L.; Wang, S.; Tao, T.; Zhao, Y.; Jiang, L. Variations in the PBEF1 Gene Are Associated with Body Mass Index: A Population-Based Study in Northern China. Meta Gene 2015, 6, 65–68. [Google Scholar] [CrossRef]
- Zhou, Q.; Chen, B.; Ji, T.; Luo, M.; Luo, J. Association of Genetic Variants in RETN, NAMPT and ADIPOQ Gene with Glycemic, Metabolic Traits and Diabetes Risk in a Chinese Population. Gene 2018, 642, 439–446. [Google Scholar] [CrossRef]
- Kim, J.-E.; Kim, J.-S.; Jo, M.-J.; Cho, E.; Ahn, S.-Y.; Kwon, Y.-J.; Ko, G.-J.T.; Roles, A.; Kim, J.-E.; Kim, J.-S.; et al. The Roles and Associated Mechanisms of Adipokines in Development of Metabolic Syndrome. Molecules 2022, 27, 334. [Google Scholar] [CrossRef]
- PPY Pancreatic Polypeptide [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/5539 (accessed on 14 November 2022).
- PYY Peptide YY [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/5697 (accessed on 14 November 2022).
- Simpson, K.; Parker, J.; Plumer, J.; Bloom, S. CCK, PYY and PP: The Control of Energy Balance. Handb. Exp. Pharmacol. 2012, 209, 209–230. [Google Scholar] [CrossRef]
- Friedlander, Y.; Li, G.; Fornage, M.; Williams, O.D.; Lewis, C.E.; Schreiner, P.; Pletcher, M.J.; Enquobahrie, D.; Williams, M.; Siscovick, D.S. Candidate Molecular Pathway Genes Related to Appetite Regulatory Neural Network, Adipocyte Homeostasis and Obesity: Results from the CARDIA Study. Ann. Hum. Genet. 2010, 74, 387–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiq, A.; Gueorguiev, M.; Samson, C.; Hercberg, S.; Heude, B.; Levy-Marchal, C.; Jouret, B.; Weill, J.; Meyre, D.; Walley, A.; et al. Single Nucleotide Polymorphisms in the Neuropeptide Y2 Receptor (NPY2R) Gene and Association with Severe Obesity in French White Subjects. Diabetologia 2007, 50, 574–584. [Google Scholar] [CrossRef] [Green Version]
- Ahituv, N.; Kavaslar, N.; Schackwitz, W.; Ustaszewska, A.; Martin, J.; Hébert, S.; Doelle, H.; Ersoy, B.; Kryukov, G.; Schmidt, S.; et al. Medical Sequencing at the Extremes of Human Body Mass. Am. J. Hum. Genet. 2007, 80, 779–791. [Google Scholar] [CrossRef] [Green Version]
- Perez-Frances, M.; van Gurp, L.; Abate, M.V.; Cigliola, V.; Furuyama, K.; Bru-Tari, E.; Oropeza, D.; Carreaux, T.; Fujitani, Y.; Thorel, F.; et al. Pancreatic Ppy-Expressing γ-Cells Display Mixed Phenotypic Traits and the Adaptive Plasticity to Engage Insulin Production. Nat. Commun. 2021, 12, 4458. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Lee, S.Y.; Kim, C.M. Association between Gene Polymorphisms and Obesity and Physical Fitness in Korean Children. Biol. Sport 2018, 35, 21–27. [Google Scholar] [CrossRef]
- RBP4 Retinol Binding Protein 4 [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/5950 (accessed on 14 November 2022).
- Yang, Q.; Graham, T.E.; Mody, N.; Preitner, F.; Peroni, O.D.; Zabolotny, J.M.; Kotani, K.; Quadro, L.; Kahn, B.B. Serum Retinol Binding Protein 4 Contributes to Insulin Resistance in Obesity and Type 2 Diabetes. Nature 2005, 436, 356–362. [Google Scholar] [CrossRef]
- Craig, R.L.; Chu, W.S.; Elbein, S.C. Retinol Binding Protein 4 as a Candidate Gene for Type 2 Diabetes and Prediabetic Intermediate Traits. Mol. Genet. Metab. 2007, 90, 338–344. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Li, H.; Loos, R.J.F.; Qi, Q.; Hu, F.B.; Liu, Y.; Lin, X. RBP4 Variants Are Significantly Associated with Plasma RBP4 Levels and Hypertriglyceridemia Risk in Chinese Hans. J. Lipid. Res. 2009, 50, 1479–1486. [Google Scholar] [CrossRef] [Green Version]
- Codoñer-Franch, P.; Carrasco-Luna, J.; Allepuz, P.; Codoñer-Alejos, A.; Guillem, V. Association of RBP4 Genetic Variants with Childhood Obesity and Cardiovascular Risk Factors. Pediatr. Diabetes 2016, 17, 576–583. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Ma, S.; Li, X.; Tian, Z.; Liang, H.; Yan, J.; Chen, M.; Tan, H. Relationships of SLC2A4, RBP4, PCK1, and PI3K Gene Polymorphisms with Gestational Diabetes Mellitus in a Chinese Population. Biomed. Res. Int. 2019, 2019, 7398063. [Google Scholar] [CrossRef] [Green Version]
- RETN Resistin [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/56729 (accessed on 14 November 2022).
- Tripathi, D.; Kant, S.; Pandey, S.; Ehtesham, N.Z. Resistin in Metabolism, Inflammation, and Disease. FEBS J. 2020, 287, 3141–3149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hishida, A.; Wakai, K.; Okada, R.; Morita, E.; Hamajima, N.; Hosono, S.; Higaki, Y.; Turin, T.C.; Suzuki, S.; Motahareh, K.; et al. Significant Interaction between RETN -420 G/G Genotype and Lower BMI on Decreased Risk of Type 2 Diabetes Mellitus (T2DM) in Japanese--the J-MICC Study. Endocr. J. 2013, 60, 237–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zayani, N.; Omezzine, A.; Boumaiza, I.; Achour, O.; Rebhi, L.; Rejeb, J.; ben Rejeb, N.; ben Abdelaziz, A.; Bouslama, A. Association of ADIPOQ, Leptin, LEPR, and Resistin Polymorphisms with Obesity Parameters in Hammam Sousse Sahloul Heart Study. J. Clin. Lab. Anal. 2017, 31, e22148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckers, S.; Zegers, D.; van Camp, J.K.; Boudin, E.; Nielsen, T.L.; Brixen, K.; Andersen, M.; van Hul, W. Resistin Polymorphisms Show Associations with Obesity, but Not with Bone Parameters in Men: Results from the Odense Androgen Study. Mol. Biol. Rep. 2013, 40, 2467–2472. [Google Scholar] [CrossRef]
- Chung, C.M.; Lin, T.H.; Chen, J.W.; Leu, H.B.; Yin, W.H.; Ho, H.Y.; Sheu, S.H.; Tsai, W.C.; Chen, J.H.; Lin, S.J.; et al. Common Quantitative Trait Locus Downstream of RETN Gene Identified by Genome-Wide Association Study Is Associated with Risk of Type 2 Diabetes Mellitus in Han Chinese: A Mendelian Randomization Effect. Diabetes Metab. Res. Rev. 2014, 30, 232–240. [Google Scholar] [CrossRef]
- Ortega, L.; Navarro, P.; Riestra, P.; Gavela-Pérez, T.; Soriano-Guillén, L.; Garcés, C. Association of Resistin Polymorphisms with Resistin Levels and Lipid Profile in Children. Mol. Biol. Rep. 2014, 41, 7659–7664. [Google Scholar] [CrossRef]
- Nakatochi, M.; Ichihara, S.; Yamamoto, K.; Ohnaka, K.; Kato, Y.; Yokota, S.; Hirashiki, A.; Naruse, K.; Asano, H.; Izawa, H.; et al. Epigenome-Wide Association Study Suggests That SNPs in the Promoter Region of RETN Influence Plasma Resistin Level via Effects on DNA Methylation at Neighbouring Sites. Diabetologia 2015, 58, 2781–2790. [Google Scholar] [CrossRef]
- Elkhattabi, L.; Morjane, I.; Charoute, H.; Amghar, S.; Bouafi, H.; Elkarhat, Z.; Saile, R.; Rouba, H.; Barakat, A. In Silico Analysis of Coding/Noncoding SNPs of Human RETN Gene and Characterization of Their Impact on Resistin Stability and Structure. J. Diabetes Res. 2019, 2019, 4951627. [Google Scholar] [CrossRef] [Green Version]
- SCT Secretin [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/6343 (accessed on 14 November 2022).
- Laurila, S.; Rebelos, E.; Honka, M.J.; Nuutila, P. Pleiotropic Effects of Secretin: A Potential Drug Candidate in the Treatment of Obesity? Front Endocrinol. 2021, 12, 1259. [Google Scholar] [CrossRef]
- Laurila, S.; Sun, L.; Lahesmaa, M.; Schnabl, K.; Laitinen, K.; Klén, R.; Li, Y.; Balaz, M.; Wolfrum, C.; Steiger, K.; et al. Secretin Activates Brown Fat and Induces Satiation. Nat. Metab. 2021, 3, 798–809. [Google Scholar] [CrossRef] [PubMed]
- UCP2 Uncoupling Protein 2 [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/7351 (accessed on 14 November 2022).
- Demine, S.; Renard, P.; Arnould, T. Mitochondrial Uncoupling: A Key Controller of Biological Processes in Physiology and Diseases. Cells 2019, 8, 795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esterbauer, H.; Schneitler, C.; Oberkofler, H.; Ebenbichler, C.; Paulweber, B.; Sandhofer, F.; Ladurner, G.; Hell, E.; Strosberg, A.D.; Patsch, J.R.; et al. A Common Polymorphism in the Promoter of UCP2 Is Associated with Decreased Risk of Obesity in Middle-Aged Humans. Nat. Genet. 2001, 28, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Bulotta, A.; Ludovico, O.; Coco, A.; di Paola, R.; Quattrone, A.; Carella, M.; Pellegrini, F.; Prudente, S.; Trischitta, V. The Common -866G/A Polymorphism in the Promoter Region of the UCP-2 Gene Is Associated with Reduced Risk of Type 2 Diabetes in Caucasians from Italy. J. Clin. Endocrinol. Metab. 2005, 90, 1176–1180. [Google Scholar] [CrossRef] [Green Version]
- Andersen, G.; Dalgaard, L.T.; Justesen, J.M.; Anthonsen, S.; Nielsen, T.; Thørner, L.W.; Witte, D.; Jørgensen, T.; Clausen, J.O.; Lauritzen, T.; et al. The Frequent UCP2-866G>A Polymorphism Protects against Insulin Resistance and Is Associated with Obesity: A Study of Obesity and Related Metabolic Traits among 17 636 Danes. Int. J. Obes. 2013, 37, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Salopuro, T.; Pulkkinen, L.; Lindström, J.; Kolehmainen, M.; Tolppanen, A.M.; Eriksson, J.G.; Valle, T.T.; Aunola, S.; Ilanne-Parikka, P.; Keinänen-Kiukaanniemi, S. Variation in the UCP2 and UCP3 genes associates with abdominal obesity and serum lipids: The Finnish Diabetes Prevention Study. BMC Med. Genet. 2009, 10, 94. [Google Scholar] [CrossRef] [Green Version]
- Oktavianthi, S.; Trimarsanto, H.; Febinia, C.A.; Suastika, K.; Saraswati, M.R.; Dwipayana, P.; Arindrarto, W.; Sudoyo, H.; Malik, S.G. Uncoupling Protein 2 Gene Polymorphisms Are Associated with Obesity. Cardiovasc. Diabetol. 2012, 11, 41. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Hervas, S.; Mansego, M.L.; de Marco, G.; Martinez, F.; Alonso, M.P.; Morcillo, S.; Rojo-Martinez, G.; Real, J.T.; Ascaso, J.F.; Redon, J.; et al. Polymorphisms of the UCP2 Gene Are Associated with Body Fat Distribution and Risk of Abdominal Obesity in Spanish Population. Eur. J. Clin. Invest. 2012, 42, 171–178. [Google Scholar] [CrossRef]
- Xu, L.; Chen, S.; Zhan, L. Association of Uncoupling Protein-2 -866G/A and Ala55Val Polymorphisms with Susceptibility to Type 2 Diabetes Mellitus: A Meta-Analysis of Case-Control Studies. Medicine 2021, 100, e24464. [Google Scholar] [CrossRef]
- Fall, T.; Mendelson, M.; Speliotes, E.K. Recent Advances in Human Genetics and Epigenetics of Adiposity: Pathway to Precision Medicine? Gastroenterology 2017, 152, 1695–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christakoudi, S.; Tsilidis, K.K.; Evangelou, E.; Riboli, E. A Body Shape Index (ABSI), hip index, and risk of cancer in the UK Biobank cohort. Cancer Med. 2021, 10, 5614–5628. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Hsu, L.-A.; Teng, M.-S.; Chou, H.-H.; Ko, Y.-L. Differential Genetic and Epigenetic Effects of the KLF14 Gene on Body Shape Indices and Metabolic Traits. Int. J. Mol. Sci. 2022, 23, 4165. [Google Scholar] [CrossRef] [PubMed]
- Nagayama, D.; Sugiura, T.; Choi, S.Y.; Shirai, K. Various Obesity Indices and Arterial Function Evaluated with CAVI—Is Waist Circumference Adequate to Define Metabolic Syndrome? Vasc. Health Risk. Manag. 2022, 18, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Nagayama, D.; Fujishiro, K.; Watanabe, Y.; Yamaguchi, T.; Suzuki, K.; Saiki, A.; Shirai, K. A Body Shape Index (ABSI) as a Variant of Conicity Index Not Affected by the Obesity Paradox: A Cross-Sectional Study Using Arterial Stiffness Parameter. J. Pers. Med. 2022, 12, 2014. [Google Scholar] [CrossRef]
- Young, K.L.; Graff, M.; Fernandez-Rhodes, L.; North, K.E. Genetics of Obesity in Diverse Populations. Curr. Diab. Rep. 2018, 18, 145. [Google Scholar] [CrossRef] [PubMed]
- Locke, A.E.; Kahali, B.; Berndt, S.I.; Justice, A.E.; Pers, T.H.; Day, F.R.; Powell, C.; Vedantam, S.; Buchkovich, M.L.; Yang, J.; et al. Genetic studies of body mass index yield new insights for obesity biology. Nature 2015, 518, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Pan, D.Z.; Miao, Z.; Comenho, C.; Rajkumar, S.; Koka, A.; Lee, S.H.T.; Alvarez, M.; Kaminska, D.; Ko, A.; Sinsheimer, J.S.; et al. Identification of TBX15 as an adipose master trans regulator of abdominal obesity genes. Genome Med. 2021, 13, 123. [Google Scholar] [CrossRef]
- Larsson, S.C.; Bäck, M.; Rees, J.M.B.; Mason, A.M.; Burgess, S. Body mass index and body composition in relation to 14 cardiovascular conditions in UK Biobank: A Mendelian randomization study. Eur. Heart J. 2020, 41, 221–226. [Google Scholar] [CrossRef] [Green Version]
- Beaney, K.E.; Cooper, J.A.; Shahid, S.U.; Ahmed, W.; Qamar, R.; Drenos, F.; Crockard, M.A.; Humphries, S.E. Clinical Utility of a Coronary Heart Disease Risk Prediction Gene Score in UK Healthy Middle-Aged Men and in the Pakistani Population. PLoS ONE 2015, 10, e0130754. [Google Scholar] [CrossRef] [Green Version]
- Iribarren, C.; Lu, M.; Jorgenson, E.; Martínez, M.; Lluis-Ganella, C.; Subirana, I.; Salas, E.; Elosua, R. Weighted Multi-marker Genetic Risk Scores for Incident Coronary Heart Disease among Individuals of African, Latino and East-Asian Ancestry. Sci. Rep. 2018, 8, 6853. [Google Scholar] [CrossRef] [Green Version]
- Semaev, S.; Shakhtshneider, E. Genetic Risk Score for Coronary Heart Disease: Review. J. Pers. Med. 2020, 10, 239. [Google Scholar] [CrossRef] [PubMed]
- Benbaibeche, H.; Bounihi, A.; Koceir, E.A. Leptin level as a biomarker of uncontrolled eating in obesity and overweight. Ir. J. Med. Sci. 2020, 190, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Poetsch, M.S.; Strano, A.; Guan, K. Role of Leptin in Cardiovascular Diseases. Front. Endocrinol. 2020, 11, 354. [Google Scholar] [CrossRef]
- Reyes-Barrera, J.; Sainz-Escárrega, V.H.; Medina-Urritia, A.X.; Jorge-Galarza, E.; Osorio-Alonso, H.; Torres-Tamayo, M.; Leal-Escobar, G.; Posadas-Romero, C.; Torre-Villalvazo, I.; Juárez-Rojas, J.G. Dysfunctional adiposity index as a marker of adipose tissue morpho-functional abnormalities and metabolic disorders in apparently healthy subjects. Adipocyte 2021, 10, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Trinh, T.; Broxmeyer, H.E. Role for Leptin and Leptin Receptors in Stem Cells During Health and Diseases. Stem. Cell Rev. Rep. 2021, 17, 511–522. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Gene | Genes Associated with the Secretory Activity of Visceral Adipocytes | References |
|---|---|---|
| ADIPOQ | This gene is expressed in adipose tissue exclusively. Mutations in this gene are associated with adiponectin deficiency. | [29] |
| ADRB3 | This receptor is located mainly in adipose tissue and is involved in the regulation of lipolysis and thermogenesis. Obesity-related and body weight–related disorders correlate with polymorphisms in this gene. | [30] |
| APLN, APLNR | Apelin is secreted by adipose tissue. Apelin and its receptor are widespread in the human body and take part in many physiological processes, such as glucose and lipid metabolism, homeostasis, endocrine responses to stress, and angiogenesis. | [31] |
| CCL2, CCL7 | Chemokines CCL2 and CL7 have been implicated in the pathogenesis of several disorders, including obesity. | [32] |
| FTO | This gene shows a strong association with the BMI, obesity risk, and T2DM. | [33] |
| GCG | The glucagon (GCG) family of peptide hormones plays a role in central control of feeding behavior. | [34] |
| GLP1R | The hormone called glucagon-like peptide 1 (GLP-1) plays an important part in the signaling cascades resulting in insulin secretion. | [35] |
| GHRL | This gene encodes ghrelin-obestatin preproprotein, which is cleaved thus yielding two peptides: ghrelin and obestatin. Ghrelin regulates multiple phenomena, including hunger and pancreatic glucose-stimulated insulin secretion. Obestatin has multiple metabolic functions, including regulation of adipocyte function and glucose metabolism. | [36] |
| GIP | This gene encodes an incretin hormone. This protein is important for glucose homeostasis because it is a potent stimulator of insulin secretion from pancreatic β -cells after food ingestion and nutrient absorption. | [37] |
| INS | This gene codes for insulin: a peptide hormone that plays a vital role in the regulation of carbohydrate and lipid metabolism. | [38] |
| LEP | This gene encodes a protein that is secreted by white adipocytes into the circulation and performs a major function in the modulation of energy homeostasis. Circulating leptin binds to leptin receptor in the brain, thereby triggering downstream signaling pathways that inhibit feeding and promote energy expenditure. Mutations in this gene and in its regulatory regions induce severe obesity. | [39] |
| NAMPT | This gene encodes a protein that participates in many important biological processes, including metabolism, stress responses, and aging. Levels of NAMPT in adipose tissue are rather high. | [40,41] |
| PPY | This hormone acts as a regulator of pancreatic and gastrointestinal functions and may be important for the modulation of food intake. | [42] |
| PYY | Rare variations in this gene may increase susceptibility to obesity. | [42] |
| RBP4 | Vitamin A can affect obesity progression and the development of obesity-related diseases including insulin resistance, T2DM, hepatic steatosis, steatohepatitis, and cardiovascular diseases. | [43] |
| RETN | This gene codes for a protein called resistin. Resistin is secreted by adipocytes and may be the hormone potentially linking obesity to T2DM. | [44,45] |
| SCT | Secretin activates brown adipose tissue, reduces central responses to appetizing food, and delays the motivation to refeed after a meal. | [46] |
| UCP2 | This gene is expressed in many tissues, and the highest expression is seen in skeletal muscle. The product of this gene plays certain roles in unregulated thermogenesis, obesity, and diabetes mellitus. | [47] |
| Gene | dbSNP ID | Nucleotide Changes | Type of Variation/Amino Acid Changes | Minor Allele Frequency (GnomAD) | ClinVar Variation ID/LOVD Database ID | Associated Metabolic Disorder |
|---|---|---|---|---|---|---|
| ADIPOQ | rs62625753 | +268 G > A | Missense Variant G > S | 0.004625 | 708724 | allele A is associated with T2DM risk |
| ADIPOQ | rs266729 | C > G | 2KB Upstream Variant | G = 0.09786 | - | allele C is associated with T2DM risk |
| ADIPOQ | rs17366743 | +331 T > C | Missense Variant Y > H | 0.030445 | - | allele C is associated with T2DM risk |
| FTO | rs9939609 | 46-23525 T > A | Intron Variant | 0.41025 | - | allele A is associated with obesity |
| FTO | rs1421085 | 46-43098 T > C | Intron Variant | 0.419704 | 214481 | allele C is associated with obesity |
| GCG | rs4664447 | 254 + 672 A > T | Intron Variant | C = 0.00792 G = 0.00000 | - | GG genotype is associated with insulin level abnormalities |
| GHRL | rs34911341 | 152 G > A | Missense Variant R > Q | 0.007960 | 20100/ 00293181 | allele A is associated with metabolic syndrome |
| GHRL | rs696217 | 214 C > A | Missense Variant L > M | 0.080526 | 20101 | allele T is associated with metabolic syndrome and obesity |
| GHRL | rs4684677 | 269 A > T | Missense Variant Q > L | 0.06097 | 20102 | allele A is associated with obesity |
| GLP1R | rs2268641 | 1224 + 1751 C > T | Intron Variant | 0.39129 | - | Associated with BMI |
| GLP1R | rs6923761 | 502 G > A | Missense Variant G > S | 0.324795 | -/00208599 | GG genotype is associated with higher BMI and CV risks |
| GLP1R | rs10305492 | 946G > A | Missense Variant A > T | 0.015874 | - | allele A is associated with T2DM risk |
| NAMPT | rs9770242 | C > A | 2KB Upstream Variant | C = 0.23919 | - | allele C is associated with lower fasting plasma glucose and insulin levels |
| NAMPT | rs1319501 | T > C | 2KB Upstream Variant | C = 0.23662 A = 0.00001 | - | allele C is associated with lower fasting plasma glucose and insulin levels |
| NAMPT | rs10487818 | 448-303 T > A | Intron Variant | 0.01841 | -/00028958 | allele T is associated with protection against obesity |
| PYY | rs11684664 | C > T | A = 0.0000 T = 0.00038 | allele T is associated with obesity-related phenotypes in women only | ||
| PYY | rs162430 | 270-61 C > T | Intron Variant | 0.105072 | - | allele A is associated with childhood obesity |
| RBP4 | rs34571439 | A > C | 500B Downstream Variant | 0.186457 | - | allele C is associated with reduced insulin secretion |
| RBP4 | rs3758539 | C > T | 2KBUpstream Variant | 0.16303 | - | allele T is associated with reduced insulin secretion |
| RETN | rs3745367 | 118 + 181 G > A | Intron Variant | 0.248110 | - | allele A is associated with obesity |
| UCP2 | rs659366 | C > T | 2KB Upstream Variant | 0.370248 | 22904 | allele T is associated with higher BMI |
| UCP2 | rs660339 | 164 C > T | Missense Variant A > V | 0.402632 | 136143 | allele A is associated with obesity |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bairqdar, A.; Ivanoshchuk, D.; Shakhtshneider, E. Functionally Significant Variants in Genes Associated with Abdominal Obesity: A Review. J. Pers. Med. 2023, 13, 460. https://doi.org/10.3390/jpm13030460
Bairqdar A, Ivanoshchuk D, Shakhtshneider E. Functionally Significant Variants in Genes Associated with Abdominal Obesity: A Review. Journal of Personalized Medicine. 2023; 13(3):460. https://doi.org/10.3390/jpm13030460
Chicago/Turabian StyleBairqdar, Ahmad, Dinara Ivanoshchuk, and Elena Shakhtshneider. 2023. "Functionally Significant Variants in Genes Associated with Abdominal Obesity: A Review" Journal of Personalized Medicine 13, no. 3: 460. https://doi.org/10.3390/jpm13030460
APA StyleBairqdar, A., Ivanoshchuk, D., & Shakhtshneider, E. (2023). Functionally Significant Variants in Genes Associated with Abdominal Obesity: A Review. Journal of Personalized Medicine, 13(3), 460. https://doi.org/10.3390/jpm13030460