
Expression of MRJP3 and HSP70 mRNA Levels in Apis mellifera L. Workers after Dietary Supplementation with Proteins, Prebiotics, and Probiotics

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Experimental Design
2.2. Total RNA Extraction and cDNA Synthesis
2.3. Real-Time Polymerase Chain Reaction
2.4. Statistical Analysis
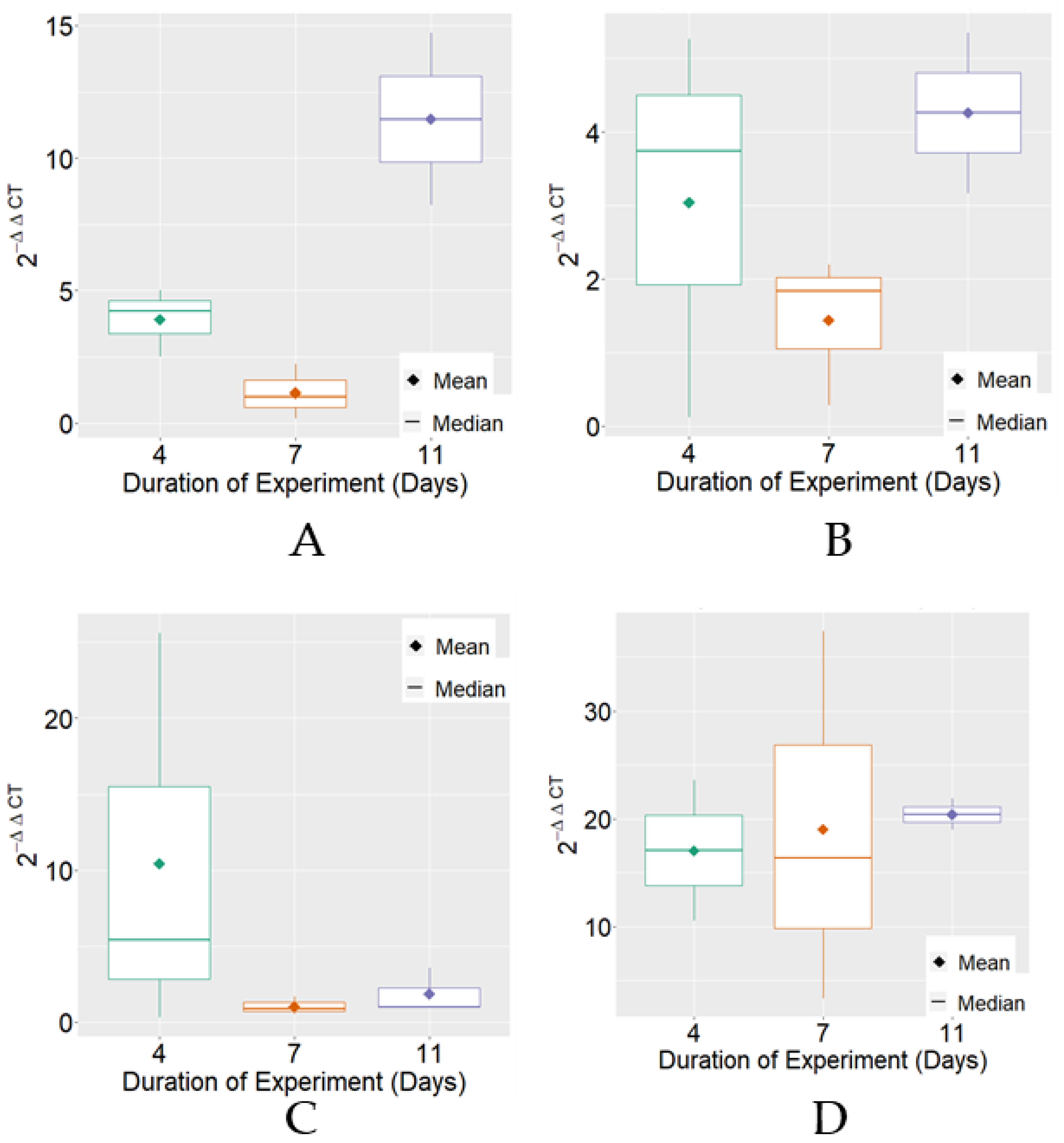
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Winston, M.L. The Biology of Honey Bee; Harvard University Press: London, UK, 1991. [Google Scholar]
- Carvalho, S.M.; Belzunces, L.P.; Carvalho, G.A.; Brunet, J.-L.; Badiou-Beneteau, A. Enzymatic biomarkers as tools to assess environmental quality: A case study of exposure of the honey bee Apis mellifera to insecticides. Environ. Toxicol. Chem. 2013, 32, 2117–2124. [Google Scholar] [CrossRef] [PubMed]
- Schmitzova, L.; Klaudiny, J.; Albert, S.; Schroeder, W.; Schreckengost, W.; Hanes, J.; Judova, J.; Simuth, J. A family of major Royal jelly proteins of the honey bee Apis mellifera L. Cell Mol. Life Sci. 1998, 54, 1020–1030. [Google Scholar] [CrossRef] [PubMed]
- Baitala, T.V.; Faquinello, P.; Toledo, V.A.A.; Mangolin, C.A.; Martins, E.N.; Ruvolo-Takasusuki, M.C.C. Potential use of major royal jelly proteins (MRJPs) as molecular markers for royal jelly production in Africanized honeybee colonies. Apidologie 2010, 41, 160–168. [Google Scholar] [CrossRef] [Green Version]
- Toledo, V.A.A.; Neves, C.A.; Alves, E.M.; Oliveira, J.R.; Ruvolo-Takasusuki, M.C.C.; Faquinello, P. Produção de geleia real em colônias de abelhas africanizadas considerando diferentes suplementos proteicos e a influência de fatores ambientais. Acta Sci.-Anim. Sci. 2010, 32, 101–108. [Google Scholar] [CrossRef]
- Buttstedt, A.; Moritz, R.F.A.; Erler, S. More than royal food—Major royal jelly protein genes in sexuals and workers of the honey bee Apis mellifera. Front. Zool. 2013, 10, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanes, J.; Simúth, J. Identification and partial characterization of the major royal jelly protein of the honey bee (Apis mellifera L.). J. Apic. Res. 1992, 31, 22–26. [Google Scholar] [CrossRef]
- Ostroverkhova, N.V.; Kucher, A.N.; Babushkina, N.P.; Konusova, O.L. Sequence of the mrjp3 microsatellite locus in honey bees of different origin. Russ. J. Genet. 2018, 54, 335–341. [Google Scholar] [CrossRef]
- The Honey bee Genome Sequencing Consortium Insights into social insects from the genome of the honey bee Apis mellifera. Nature 2006, 443, 931–949. [CrossRef]
- Ruvolo-Takasusuki, M.C.C.; Pozza, A.P.B.C.; Oliveira, A.P.N.Z.; Parpinelli, R.S.; Costa-Maia, F.M.; Faquinello, P.; Toledo, V.A.A. Improvement and Selection of Honey bees Assisted by Molecular Markers Beekeeping and Bee Conservation. In Beekiping and Bee Conservation Advances in Research; Chambó, E.D., Ed.; IntechOpen: Rijeka, Croatia, 2016; pp. 63–75. [Google Scholar] [CrossRef] [Green Version]
- Even, N.; Devaud, J.-M.; Barron, A.B. General stress responses in the honey bee. Insects 2012, 3, 1271–1298. [Google Scholar] [CrossRef] [Green Version]
- Lindquist, S.; Craig, E.A. The heat-shock proteins. Annu. Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef]
- Meyer, T.N.; Silva, A.L. Resposta celular ao estresse. Rev. Assoc. Médica Bras. 1999, 45, 181–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, T.E.; Fofah, O.; Dinges, C.W.; Ortiz-Alvarado, C.A.; Avalos, A.; Ortiz-Alvarado, Y.; Abramson, C.I. Effects of aversive conditioning on expression of physiological stress in honey bees (Apis mellifera). Neurobiol. Learn. Mem. 2021, 178, 1074–7427. [Google Scholar] [CrossRef] [PubMed]
- Sereia, M.J.; Toledo, V.A.A.; Furlan, A.C.; Faquinello, P.; Maia, F.M.C.; Wielewski, P. Alternative sources of supplements for Africanized honeybees submitted to royal jelly production. Acta Sci.-Anim. Sci. 2013, 35, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Pereira, F.M.; Freitas, B.M.; Neto, J.M.V.; Lopes, M.T.R.; Barbosa, A.L.; Camargo, R.C.R. Desenvolvimento de colônias de abelhas com diferentes alimentos proteicos. Pesq. Agropec. Bras. 2006, 41, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Turcatto, A.P. Desenvolvimento e análise do efeito de dietas protéicas como suplementação nutricional para abelhas Apis mellifera. Master’s Thesis, Universidade de São Paulo-USP, Ribeirão Preto, Brazil, 2011. [Google Scholar]
- Pachla, A.; Ptaszynska, A.A.; Wicha, M.; Kunat, M.; Wydrych, J.; Olenska, E.; Małek, W. Insight into probiotic properties of lactic acid bacterial endosymbionts of Apis mellifera L. derived from the Polish apiary. Saudi J. Biol. Sci. 2021, 28, 1890–1899. [Google Scholar] [CrossRef] [PubMed]
- Stefe, C.A.; Ribeiro-Alves, M.A.; Ribeiro, R.L. Probióticos, prebióticos e simbióticos. Rev. Saúde Ambiente 2008, 3, 16–33. [Google Scholar]
- Williams, G.R.; Alaux, C.; Costa, C.; Csáki, T.; Doublet, V.; Eisenhardt, D.; Fries, I.; Kuhn, R.; Mcmahon, D.P.; Medrzycki, P.; et al. Standard methods for maintaining adult Apis mellifera in cages under in vitro laboratory conditions. J. Apic. Res. 2013, 52, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Lima, E.G.; Parpinelli, R.S.; Santos, P.R.; Azevedo, A.S.B.; Sereia, M.J.; Toledo, V.A.A. Optimization of protein feed fermentation process for supplementation of Apis mellifera honeybees. J. Apic. Sci. 2020, 64, 15–27. [Google Scholar] [CrossRef]
- Jeon, J.H.; Moon, K.; Kim, Y.; Kim, Y.H. Reference gene selection for qRT-PCR analysis of season- and tissue-specific gene expression profiles in the honey bee Apis mellifera. Sci. Rep. 2020, 10, 13935. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer: Houston, TA, USA, 2016. [Google Scholar]
- Feng, M.; Fang, Y.; Li, J. Proteomic analysis of honey bee worker (Apis mellifera) hypopharyngeal gland development. BMC Genom. 2009, 10, 645. [Google Scholar] [CrossRef] [Green Version]
- Jianke, L.; Feng, M.; Begna, D.; Fang, Y.; Zheng, A. Proteome comparison of hypopharyngeal gland development between Italian and royal jelly producing worker honey bees (Apis mellifera L.). J. Proteome Res. 2010, 9, 6578–6594. [Google Scholar] [PubMed]
- Deseyn, J.; Billen, J. Age-dependent morphology and ultrastructure of the hypopharyngeal gland of Apis mellifera workers (Hymenoptera, Apidae). Apidologie 2005, 36, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Dobritzsch, D.; Aumer, D.; Fuszard, M.; Erler, S.; Buttstedt, A. The rise and fall of major royal jelly proteins during a honey bee (Apis mellifera) workers’ life. Ecol. Evol. 2019, 9, 8771–8782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.K.; Lee, M.L.; Lee, M.; Yong-Soo, C.; Han, S.M.; Kang, A.R.; Lee, K.Y. Evaluation of Royal Jelly Productivity and Characteristics in Apis mellifera Inbred Lines. Korean J. Apic. 2017, 32, 155–162. [Google Scholar]
- Fernández-Fernández, M.R.; Valpuesta, J.M. Hsp70 chaperone: A master player in protein homeostasis. F1000Research 2018, 7, 1497. [Google Scholar] [CrossRef] [Green Version]
- Genova, J.L.; Rodrigues, R.B.; Martins, J.S.; Uczay, M.; Henriques, J.K.S. Propolis and bee pollen in the nutrition of non ruminant animals. Arch. Zootc. 2020, 69, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Hranitz, J.M.; Barthell, J.F.; Abramson, C.I.; Brubaker, K.D.; Wells, H. Stress protein responses in honey bees: Is it useful to measure stress responses of individual bees in the hive? Uludag Bee J. 2009, 9, 60–71. [Google Scholar]
- Beere, H.M. ‘The stress of dying’: The role of heat shock proteins in the regulation of apoptosis. J. Cell Sci. 2004, 117, 2641–2651. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene | Amplicon Size (bp) | Annealing Temperature (°C) | Primer Sequence 5′-3′ |
|---|---|---|---|
| Actin Forward | 134 | 60 | CCATGTATCCTGGAATCGCAG |
| Actin Reverse | AGAAGCAAGAATTGACCCACC | ||
| mrjp3 Forward | 143 | 60 | TTGACAGTC GCTGGA GAA AG |
| mrjp3 Reverse | GTGGATTGCTGAATTGTTCCG | ||
| hsp70 Forward | 74 | 60 | CAAGAGAGGAACACGACCATACC |
| hsp70 Reverse | AGACGCCAGGTTGATTATCG |
| Variation Sources | DF | Mean Square |
|---|---|---|
| −ΔΔCT | ||
| PE | 2 | 5.10 |
| T | 3 | 320.50 *** |
| P | 1 | 172.50 *** |
| PE × T | 6 | 121.40 *** |
| PE × P | 2 | 142.70 *** |
| T × P | 3 | 510.50 *** |
| PE × T × P | 6 | 120.18 *** |
| Residual | 48 | 42.60 |
| CV (%) | - | 102.37 |
| Overall Mean | - | 6.37 |
| Periods of the Experiment | Treatments | 2−∆∆CT | |
|---|---|---|---|
| MRJP3 | HSP70 | ||
| 4 | Negative Control | 3.91 aC | 2.00 aB |
| 4 | Positive Control | 3.04 aC | 1.13 aB |
| 4 | BPF | 10.43 aB | 12.84 aA |
| 4 | FPP | 17.06 aA | 1.04 bB |
| 7 | Negative Control | 1.13 aB | 1.16 aB |
| 7 | Positive Control | 1.44 aB | 1.59 aB |
| 7 | BPF | 1.02 bB | 28.56 aA |
| 7 | FPP | 19.03 aA | 0.48 bB |
| 11 | Negative Control | 11.47 aB | 0.95 bB |
| 11 | Positive Control | 4.26 BC | 1.32 aB |
| 11 | BPF | 1.84 BC | 0.81 aB |
| 11 | FPP | 20.41 aA | 6.03 bA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, M.C.P.V.; Pereira, E.M.; Sereia, M.J.; Lima, É.G.; Silva, B.G.; Toledo, V.A.A.; Ruvolo-Takasusuki, M.C.C. Expression of MRJP3 and HSP70 mRNA Levels in Apis mellifera L. Workers after Dietary Supplementation with Proteins, Prebiotics, and Probiotics. Insects 2022, 13, 571. https://doi.org/10.3390/insects13070571
Oliveira MCPV, Pereira EM, Sereia MJ, Lima ÉG, Silva BG, Toledo VAA, Ruvolo-Takasusuki MCC. Expression of MRJP3 and HSP70 mRNA Levels in Apis mellifera L. Workers after Dietary Supplementation with Proteins, Prebiotics, and Probiotics. Insects. 2022; 13(7):571. https://doi.org/10.3390/insects13070571
Chicago/Turabian StyleOliveira, Maria Carolina Paleari Varjão, Eloisa Magalhaes Pereira, Maria Josiane Sereia, Érica Gomes Lima, Breno Gabriel Silva, Vagner Alencar Arnaut Toledo, and Maria Claudia Colla Ruvolo-Takasusuki. 2022. "Expression of MRJP3 and HSP70 mRNA Levels in Apis mellifera L. Workers after Dietary Supplementation with Proteins, Prebiotics, and Probiotics" Insects 13, no. 7: 571. https://doi.org/10.3390/insects13070571
APA StyleOliveira, M. C. P. V., Pereira, E. M., Sereia, M. J., Lima, É. G., Silva, B. G., Toledo, V. A. A., & Ruvolo-Takasusuki, M. C. C. (2022). Expression of MRJP3 and HSP70 mRNA Levels in Apis mellifera L. Workers after Dietary Supplementation with Proteins, Prebiotics, and Probiotics. Insects, 13(7), 571. https://doi.org/10.3390/insects13070571