
Combined Effects of Soil Silicon and Host Plant Resistance on Planthoppers, Blast and Bacterial Blight in Tropical Rice



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Herbivores
2.2. Disease Organisms
2.3. Plant Materials
2.4. Soil Preparation
2.5. Effects of Nitrogen and Silicon on Seedling Growth
2.6. Effects of Soil Silicon on Rice Resistance to Planthoppers and Leafhoppers
2.6.1. Antibiosis Tests
2.6.2. Antixenosis Tests
2.7. Effects of Soil Silicon on Rice Resistance to Blast
2.8. Effects of Soil Silicon on Rice Resistance to Two Strains of Bacterial Blight
2.9. Data Analyses
3. Results
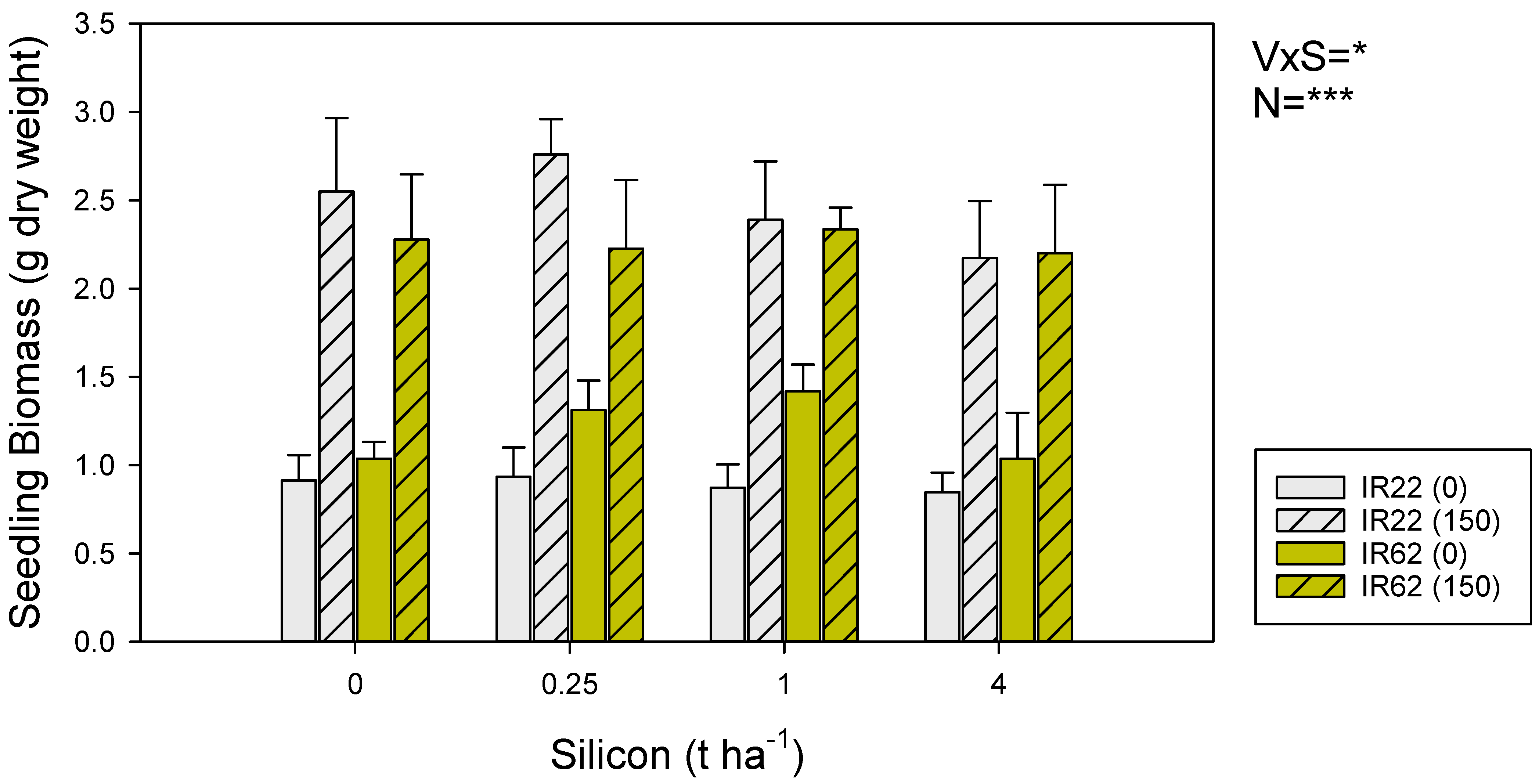
3.1. Seedling Growth under Varying Soil Silicon
3.2. Effects of Resistance, Nitrogen and Silicon on Antibiosis Defenses against Planthoppers and Leafhoppers (Philippines)
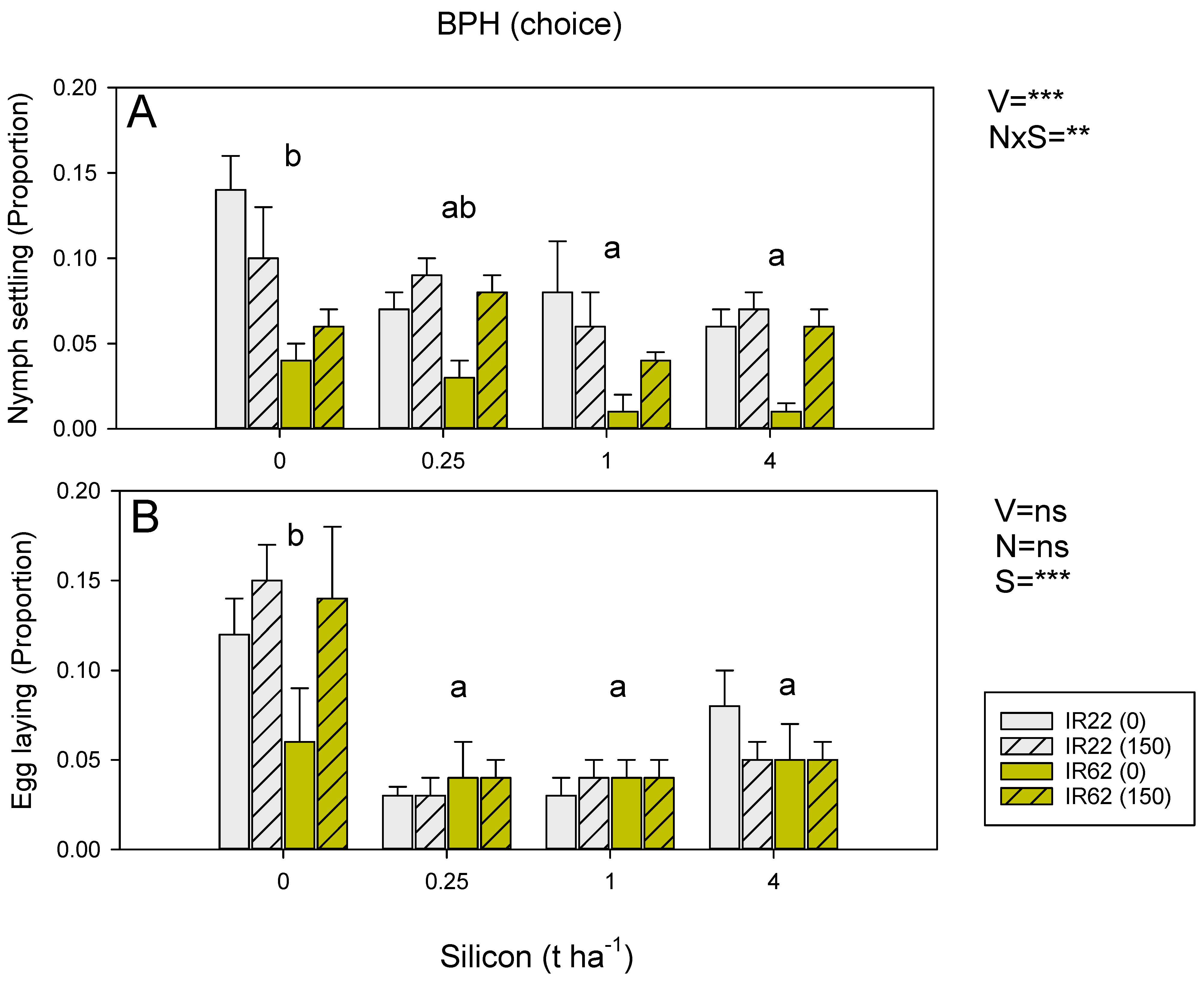
3.3. Effects of Resistance, Nitrogen and Silicon on Antixenotic Defenses against Planthoppers and Leafhoppers (Philippines)
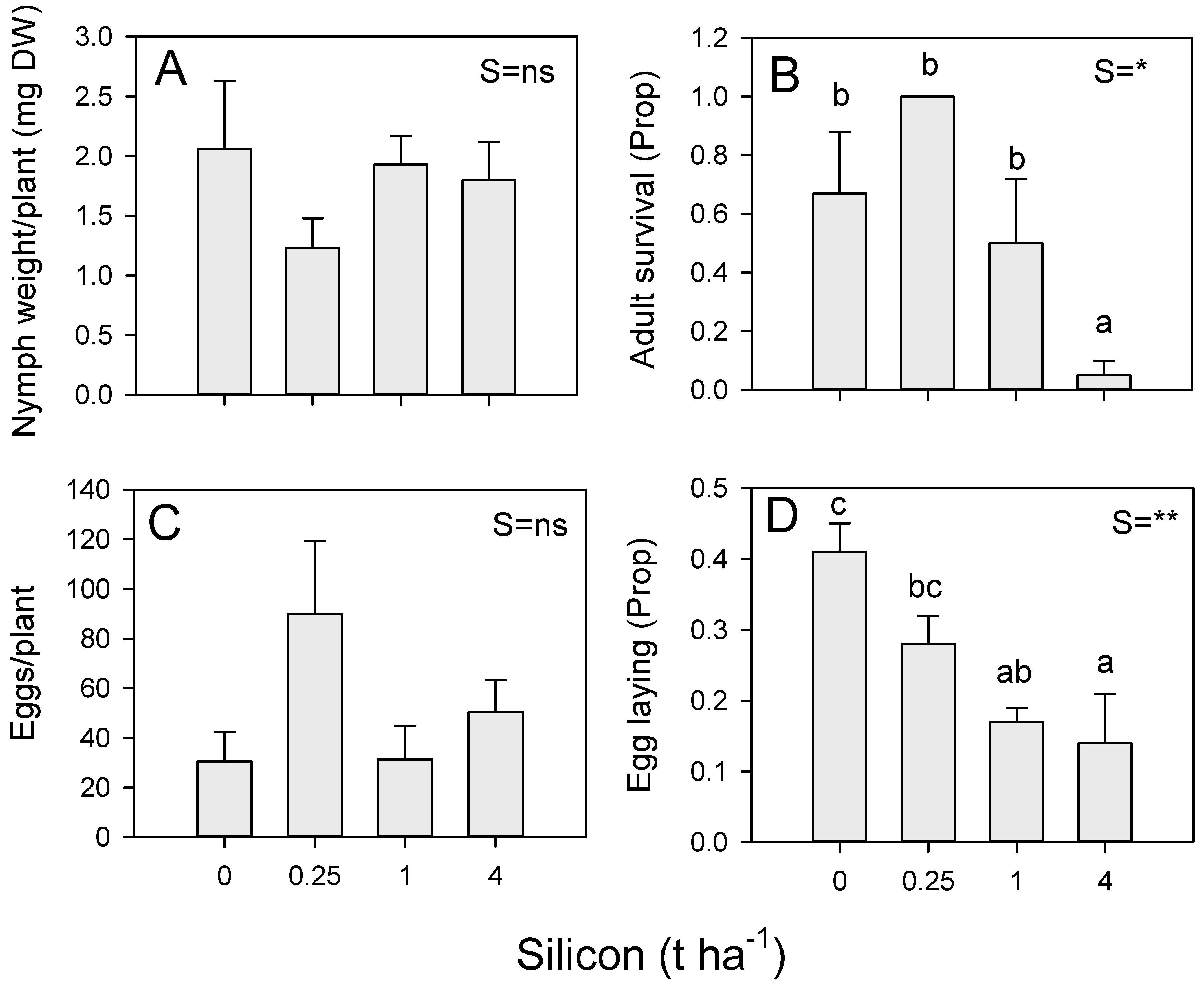
3.4. Effects of Silicon on Antibiosis and Antixenosis Defenses against BPH (Vietnam)
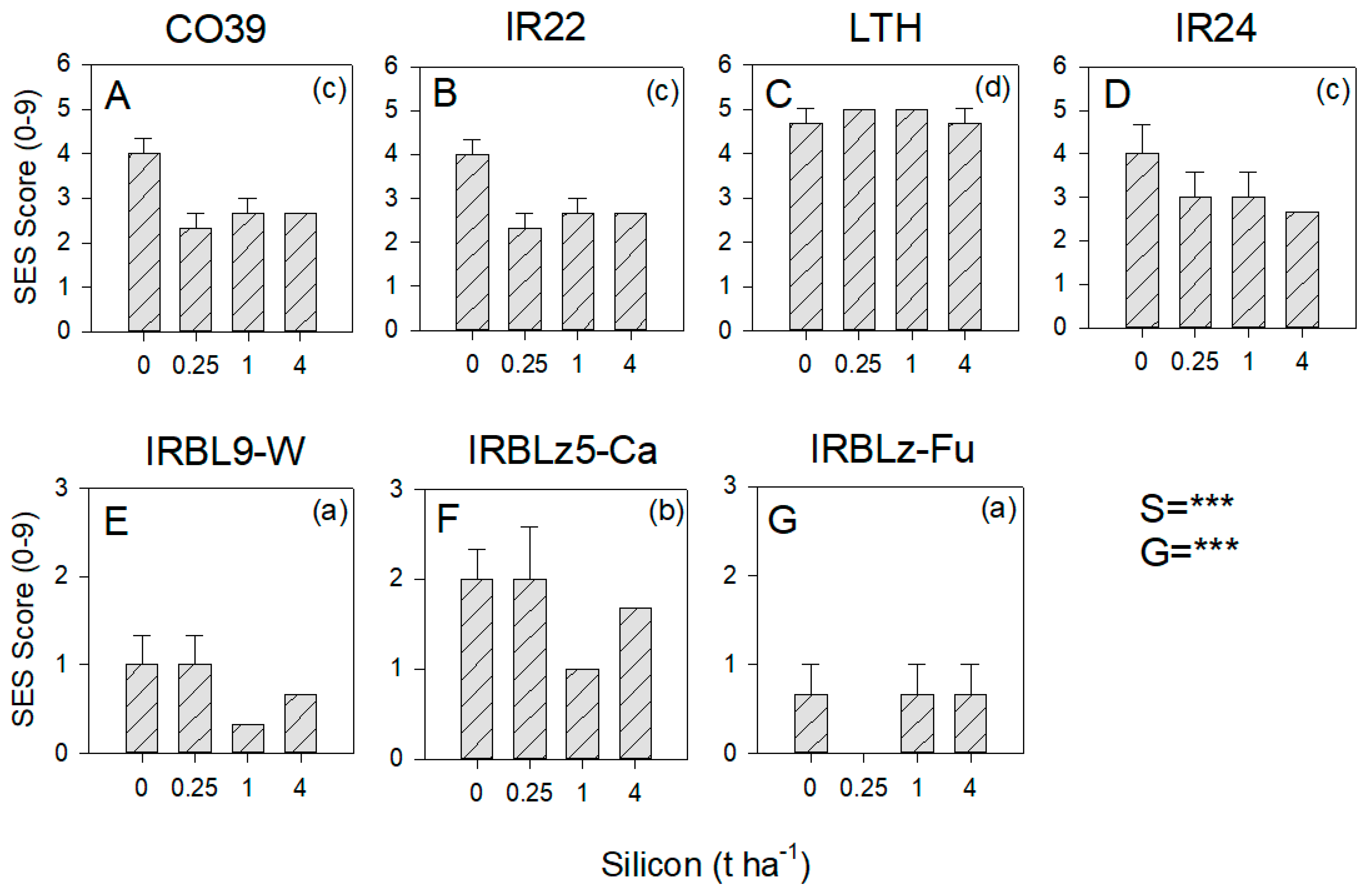
3.5. Effect of Resistance and Silicon on Blast Severity
3.6. Effect of Resistance and Silicon on Xoo Severity
4. Discussion
4.1. Direct Effects of Soil Silicon on Rice Growth
4.2. Silicon-Enhanced Resistance against Planthoppers and Leafhoppers
4.3. Combined Effects of Silicon and Nitrogen on Resistance to Planthoppers and Leafhoppers
4.4. Effects of Soil Silicon on Disease Resistance
4.5. Silicon Improves General Rice Plant Health
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, J.F. Functions of silicon in higher plants. In Silicon Biomineralization: Biology-Biochemistry-Molecular Biology-Biotechnology; Müller, W.E.G., Ed.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 127–147. [Google Scholar]
- Chandramani, P. Effect of fly ash on the feeding activity of brown planthopper and defence chemicals in rice plant. ORYZA 2014, 51, 150–154. [Google Scholar]
- Rodrigues, F.Á.; Datnoff, L.E.; Korndörfer, G.H.; Seebold, K.W.; Rush, M.C. Effect of silicon and host resistance on sheath blight development in rice. Plant Dis. 2001, 85, 827–832. [Google Scholar] [CrossRef] [Green Version]
- Hendawy, A.; Sherif, M.; El-Sayed, A.; Omar, A.; Taha, A. Role of the egg parasitoid, Trichogramma evanescens West., release and silica applications in controlling of the stem borer, Chilo agamemnon Bles. (Lepidoptera: Crambidae), in rice fields in Egypt. Egypt. J. Biol. Pest Control 2018, 28, 92. [Google Scholar] [CrossRef]
- Reynolds, O.L.; Keeping, M.G.; Meyer, J.H. Silicon-augmented resistance of plants to herbivorous insects: A review. Ann. Appl. Biol. 2009, 155, 171–186. [Google Scholar] [CrossRef]
- Reynolds, O.L.; Padula, M.P.; Zeng, R.; Gurr, G.M. Silicon: Potential to promote direct and indirect effects on plant defense against arthropod pests in agriculture. Front. Plant Sci. 2016, 7, 744. [Google Scholar] [CrossRef] [Green Version]
- Van Bockhaven, J.; De Vleesschauwer, D.; Höfte, M. Towards establishing broad-spectrum disease resistance in plants: Silicon leads the way. J. Exp. Bot. 2013, 64, 1281–1293. [Google Scholar] [CrossRef]
- Cai, K.; Gao, D.; Chen, J.; Luo, S. Probing the mechanisms of silicon-mediated pathogen resistance. Plant Signal. Behav. 2009, 4, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Han, Y.; Li, P.; Li, F.; Ali, S.; Hou, M. Silicon amendment is involved in the induction of plant defense responses to a phloem feeder. Sci. Rep. 2017, 7, 4232. [Google Scholar] [CrossRef]
- Yang, L.; Han, Y.; Li, P.; Wen, L.; Hou, M. Silicon amendment to rice plants impairs sucking behaviors and population growth in the phloem feeder Nilaparvata lugens (Hemiptera: Delphacidae). Sci. Rep. 2017, 7, 1101. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhu, J.; Zhang, P.; Han, L.; Reynolds, O.L.; Zeng, R.; Wu, J.; Shao, Y.; You, M.; Gurr, G. Silicon supplementation alters the composition of herbivore induced plant volatiles and enhances attraction of parasitoids to infested rice plants. Front. Plant Sci. 2017, 8, 1265. [Google Scholar] [CrossRef] [Green Version]
- Leroy, N.; de Tombeur, F.; Walgraffe, Y.; Cornélis, J.-T.; Verheggen, F.J. Silicon and plant natural defenses against insect pests: Impact on plant volatile organic compounds and cascade effects on multitrophic interactions. Plants 2019, 8, 444. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Tamai, K.; Konishi, S.; Fujiwara, T.; Katsuhara, M.; Yano, M. An efflux transporter of silicon in rice. Nature 2007, 448, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Klotzbücher, T.; Leuther, F.; Marxen, A.; Vetterlein, D.; Horgan, F.G.; Jahn, R. Forms and fluxes of potential plant-available silicon in irrigated lowland rice production (Laguna, the Philippines). Plant Soil 2015, 393, 177–191. [Google Scholar] [CrossRef]
- Klotzbücher, A.; Klotzbücher, T.; Jahn, R.; Xuan, L.D.; Cuong, L.Q.; Van Chien, H.; Hinrichs, M.; Sann, C.; Vetterlein, D. Effects of Si fertilization on Si in soil solution, Si uptake by rice, and resistance of rice to biotic stresses in Southern Vietnam. Paddy Water Environ. 2018, 16, 243–252. [Google Scholar] [CrossRef]
- Winslow, M.D.; Okada, K.; Correa-Victoria, F. Silicon deficiency and the adaptation of tropical rice ecotypes. Plant Soil 1997, 188, 239–248. [Google Scholar] [CrossRef]
- Klotzbücher, T.; Marxen, A.; Vetterlein, D.; Schneiker, J.; Türke, M.; van Sinh, N.; Manh, N.H.; Van Chien, H.; Marquez, L.; Villareal, S. Plant-available silicon in paddy soils as a key factor for sustainable rice production in Southeast Asia. Basic Appl. Ecol. 2015, 16, 665–673. [Google Scholar] [CrossRef]
- Marxen, A.; Klotzbücher, T.; Jahn, R.; Kaiser, K.; Nguyen, V.S.; Schmidt, A.; Schädler, M.; Vetterlein, D. Interaction between silicon cycling and straw decomposition in a silicon deficient rice production system. Plant Soil 2016, 398, 153–163. [Google Scholar] [CrossRef]
- Jeer, M.; Suman, K.; Maheswari, T.U.; Voleti, S.R.; Padmakumari, A.P. Rice husk ash and imidazole application enhances silicon availability to rice plants and reduces yellow stem borer damage. Field Crops Res. 2018, 224, 60–66. [Google Scholar] [CrossRef]
- Voleti, S.R.; Padmakumari, A.P.; Raju, V.S.; Babu, S.M.; Ranganathan, S. Effect of silicon solubilizers on silica transportation, induced pest and disease resistance in rice (Oryza sativa L.). Crop Prot. 2008, 27, 1398–1402. [Google Scholar] [CrossRef]
- Seebold, K.W.; Datnoff, L.E.; Correa-Victoria, F.J.; Kucharek, T.A.; Snyder, G.H. Effects of silicon and fungicides on the control of leaf and neck blast in upland rice. Plant Dis. 2004, 88, 253–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, F.A.; Datnoff, L.E. Silicon and rice disease management. Fitopatol. Bras. 2005, 30, 457–469. [Google Scholar] [CrossRef]
- Alvarez, J.; Datnoff, L.E. The economic potential of silicon for integrated management and sustainable rice production. Crop Prot. 2001, 20, 43–48. [Google Scholar] [CrossRef]
- Heinrichs, E.A. Biology and Management of Rice Insects; Internation Rice Research Institute: Los Baños, Philippines, 1994. [Google Scholar]
- Ou, S.H. Rice Diseases; International Rice Research Institute: Los Baños, Philippines, 1985. [Google Scholar]
- Rajamani, K.; Gunti, B.R.; Vemuri, S.; Bellamkonda, R. Effect of silicon and nitrogen nutrition on pest and disease intensity in rice. J. Agric. Sci. Technol. 2013, A3, 568–574. [Google Scholar]
- Mishra, I.O.; Panda, S.; Dash, A. Effect of organic and inorganic silicon amendments against yellow stem borer (Scirpophaga incertulas Walker) and leaf folder (Cnaphalocrocis medinalis Guenee) of rice in Costal Odisha. J. Entomol. Zool. Stud. 2018, 6, 1495–1499. [Google Scholar]
- Villegas, J.M.; Way, M.O.; Pearson, R.A.; Stout, M.J. Integrating soil silicon amendment into management programs for insect pests of drill-seeded rice. Plants 2017, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Tripathy, S.; Rath, L. Effect of silica on yellow stem borer in rice. Indian J. Entomol. 2018, 80, 203–206. [Google Scholar] [CrossRef]
- Han, Y.; Wen, J.; Peng, Z.; Zhang, D.; Hou, M. Effects of silicon amendment on the occurrence of rice insect pests and diseases in a field test. J. Integr. Agric. 2018, 17, 2172–2181. [Google Scholar] [CrossRef]
- Chandramani, P.; Rajendran, R.; Muthiah, C.; Chinniah, C. Organic source induced silica on leaf folder, stem borer and gall midge population and rice yield. J. Biopestic. 2010, 3, 423–427. [Google Scholar]
- Seebold, K.W.; Datnoff, L.E.; Correa-Victoria, F.J.; Kucharek, T.A.; Snyder, G.H. Effect of silicon rate and host resistance on blast, scald, and yield of upland rice. Plant Dis. 2000, 84, 871–876. [Google Scholar] [CrossRef]
- Wang, J.; Xue, R.; Ju, X.; Yan, H.; Gao, Z.; Esmail, A.E.M.; Hu, L.; Zeng, R.; Song, Y. Silicon-mediated multiple interactions: Simultaneous induction of rice defense and inhibition of larval performance and insecticide tolerance of Chilo suppressalis by sodium silicate. Ecol. Evol. 2020, 10, 4816–4827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horgan, F.G. Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers. Crop Prot. 2018, 110, 21–33. [Google Scholar] [CrossRef]
- Dossa, G.S.; Quibod, I.; Atienza-Grande, G.; Oliva, R.; Maiss, E.; Vera Cruz, C.; Wydra, K. Rice pyramided line IRBB67 (Xa4/Xa7) homeostasis under combined stress of high temperature and bacterial blight. Sci. Rep. 2020, 10, 683. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, N.; Telebanco-Yanoria, M.J.; Tsunematsu, H.; Kato, H.; Imbe, T.; Fukuta, Y. Development of new sets of international standard differential varieties for blast resistance in rice (Oryza sativa L.). JARQ 2007, 41, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Telebanco-Yanoria, M.J.; Imbe, T.; Kato, H.; Tsunematsu, H.; Ebron, L.A.; Vera Cruz, C.M.; Kobayashi, N.; Fukuta, Y. A set of standard differential blast isolates (Magnaporthe grisea (Hebert) Barr.) from the Philippines for rice (Oryza sativa L.) resistance. JARQ 2008, 42, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Telebanco-Yanoria, M.J.; Koide, Y.; Fukuta, Y.; Imbe, T.; Kato, H.; Tsunematsu, H.; Kobayashi, N. Development of near-isogenic lines of Japonica-type rice variety Lijiangxintuanheigu as differentials for blast resistance. Breed. Sci. 2010, 60, 629–638. [Google Scholar] [CrossRef] [Green Version]
- Hou, M.; Han, Y. Silicon-mediated rice plant resistance to the Asiatic rice borer (Lepidoptera: Crambidae): Effects of silicon amendment and rice varietal resistance. J. Econ. Entomol. 2010, 103, 1412–1419. [Google Scholar] [CrossRef]
- Sidhu, J.K.; Stout, M.J.; Blouin, D.C.; Datnoff, L.E. Effect of silicon soil amendment on performance of sugarcane borer, Diatraea saccharalis (Lepidoptera: Crambidae) on rice. Bull. Entomol. Res. 2013, 103, 656–664. [Google Scholar] [CrossRef]
- Horgan, F.G.; Romena, A.M.; Bernal, C.C.; Almazan, M.L.P.; Ramal, A.F. Stem borers revisited: Host resistance, tolerance, and vulnerability determine levels of field damage from a complex of Asian rice stemborers. Crop Prot. 2021, 142, 105513. [Google Scholar] [CrossRef]
- Fujita, D.; Kohli, A.; Horgan, F.G. Rice resistance to planthoppers and leafhoppers. Crit. Rev. Plant Sci. 2013, 32, 162–191. [Google Scholar] [CrossRef]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef]
- Horgan, F.G.; de Freitas, T.F.S.; Crisol-Martínez, E.; Mundaca, E.A.; Bernal, C.C. Nitrogenous fertilizer reduces resistance but enhances tolerance to the brown planthopper in fast-growing, moderately resistant rice. Insects 2021, 12, 989. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.M.; Jahan, M.; Islam, K.S. Impact of nitrogen, phosphorus and potassium on brown planthopper and tolerance to its host rice plants. Rice Sci. 2016, 23, 119–131. [Google Scholar] [CrossRef] [Green Version]
- Rebitanim, N.A.; Rebitanim, N.Z.; Tajudin, N.S. Impact of silicon in managing important rice diseases: Blast, sheath blight, brown spot and grain discoloration. Int. J. Agron. Agric. Res. 2015, 6, 71–85. [Google Scholar]
- Horgan, F.G.; Srinivasan, T.S.; Bentur, J.S.; Kumar, R.; Bhanu, K.V.; Sarao, P.S.; Van Chien, H.; Almazan, M.L.P.; Bernal, C.C.; Ramal, A.F.; et al. Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers: Implications for rice breeding and gene deployment. Agronomy 2017, 7, 62. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Gao, F.; Wu, X.; Lu, X.; Zeng, L.; Lv, J.; Su, X.; Luo, H.; Ren, G. Bph32, a novel gene encoding an unknown SCR domain-containing protein, confers resistance against the brown planthopper in rice. Sci. Rep. 2016, 6, 37645. [Google Scholar] [CrossRef]
- Nguyen, C.D.; Zheng, S.H.; Sanada-Morimura, S.; Matsumura, M.; Yasui, H.; Fujita, D. Substitution mapping and characterization of brown planthopper resistance genes from indica rice variety, ’PTB33’ (Oryza sativa L.). Breed. Sci. 2021, 71, 497–509. [Google Scholar] [CrossRef]
- Ferrater, J.B.; Naredo, A.I.; Almazan, M.L.P.; de Jong, P.W.; Dicke, M.; Horgan, F.G. Varied responses by yeast-like symbionts during virulence adaptation in a monophagous phloem-feeding insect. Arthropod-Plant Interact. 2015, 9, 215–224. [Google Scholar] [CrossRef]
- Mui, P.; Bong, B. Evaluation of rice varieties for resistance to brown planthopper in the Mekong Delta. Omonrice 1999, 7, e7. [Google Scholar]
- Zhang, Z.; Cui, B.; Yan, S.; Li, Y.; Xiao, H.; Li, Y.; Zhang, Y. Evaluation of tricin, a stylet probing stimulant of brown planthopper, in infested and non-infested rice plants. J. Appl. Entomol. 2017, 141, 393–401. [Google Scholar] [CrossRef]
- Chauhan, R.; Farman, M.; Zhang, H.B.; Leong, S. Genetic and physical mapping of a rice blast resistance locus, Pi-CO39(t), that corresponds to the avirulence gene AVR1-CO39 of Magnaporthe grisea. Mol. Genet. Genom. 2002, 267, 603–612. [Google Scholar] [CrossRef]
- Joshi, B.K.; Bimb, H.P.; Parajuli, G.; Chaudhary, B. Molecular tagging, allele mining and marker aided breeding for blast resistance in rice. BSN E-Bull. 2009, 1, 1–23. [Google Scholar]
- Snyder, G. Methods for silicon analysis in plants, soils, and fertilizers. In Studies in Plant Science; Dantoff, L.E., Snyder, G.H., Korndörfer, G.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2001; pp. 185–196. [Google Scholar]
- IRRI. Standard Evaluation System for Rice (SES); International Rice Research Institute (IRRI): Los Baños, Philippines, 2002. [Google Scholar]
- Han, Y.Q.; Li, P.; Gong, S.L.; Yang, L.; Wen, L.Z.; Hou, M.L. Defense responses in rice induced by silicon amendment against infestation by the leaf folder Cnaphalocrocis medinalis. PLoS ONE 2016, 11, e0153918. [Google Scholar] [CrossRef] [PubMed]
- Annamalai, M.; Guru-Pirasanna-Pandi, G.; Chellapandiyan, M.; Adak, T.; Basana-Gowda, G.; Patil, N.; Rath, P.C.; Sentil-Nathan, S. Silica amendment enhances resistance of rice to yellow stem borer Scirpophaga Incertulas (Walker) with no detrimental effect on non-target organism Eisenia fetida (Savigny). Silicon, 2022; online early. [Google Scholar] [CrossRef]
- Jia, L.; Han, Y.; Hou, M. Silicon amendment to rice plants reduces the transmission of southern rice black-streaked dwarf virus by Sogatella furcifera. Pest Manag. Sci. 2021, 77, 3233–3240. [Google Scholar] [CrossRef] [PubMed]
- França, L.L.; Dierings, C.A.; Almeida, A.C.D.; Araujo, M.D.; Heinrichs, E.A.; da Silva, A.R.; Barrigossi, J.A.F.; de Jesus, F.G. Resistance in rice to Tibraca limbativentris (Hemiptera: Pentatomidae) influenced by plant silicon content. Fla. Entomol. 2018, 101, 587–591. [Google Scholar] [CrossRef] [Green Version]
- Cheng, B.; Chen, F.; Wang, C.; Liu, X.; Yue, L.; Cao, X.; Wang, Z.; Xing, B. The molecular mechanisms of silica nanomaterials enhancing the rice (Oryza sativa L.) resistance to planthoppers (Nilaparvata lugens Stål). Sci. Total Environ. 2021, 767, 144967. [Google Scholar] [CrossRef]
- Lin, Y.; Sun, Z.; Li, Z.; Xue, R.; Cui, W.; Sun, S.; Liu, T.; Zeng, R.; Song, Y. Deficiency in silicon transporter Lsi1 compromises inducibility of anti-herbivore defense in rice plants. Front. Plant Sci. 2019, 10, 652. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Yu, Y.; Baerson, S.R.; Song, Y.; Liang, G.; Ding, C.; Niu, J.; Pan, Z.; Zeng, R. Interactions between nitrogen and silicon in rice and their effects on resistance toward the brown planthopper Nilaparvata lugens. Front. Plant Sci. 2017, 8, 28. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, S.Z.; Jelodar, N.B.; Bagheri, N. Study of silicon effects on plant growth and resistance to stem borer in rice. Commun. Soil Sci. Plant Anal. 2012, 43, 2744–2751. [Google Scholar] [CrossRef]
- Wattanapayapkul, W.; Polthanee, A.; Siri, B.; Bhadalung, N.; Promkhambut, A. Effects of silicon in suppressing blast disease and increasing grain yield of organic rice in Northeast Thailand. Asian J. Plant Pathol. 2011, 5, 134–145. [Google Scholar] [CrossRef]
- Horgan, F.G.; Nogues Palenzuela, A.; Stuart, A.M.; Naredo, A.I.; Ramal, A.F.; Bernal, C.C.; Almazan, M.L.P. Effects of silicon soil amendments and nitrogen fertilizer on apple snail (Ampullariidae) damage to rice seedlings. Crop Prot. 2017, 91, 123–131. [Google Scholar] [CrossRef]
- Yang, L.; Li, P.; Li, F.; Ali, S.; Sun, X.; Hou, M. Silicon amendment to rice plants contributes to reduced feeding in a phloem-sucking insect through modulation of callose deposition. Ecol. Evol. 2018, 8, 631–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.; Yang, M.; Li, Z.; Qiu, J.; Liu, F.; Qu, X.; Qiu, Y.; Li, R. High levels of silicon provided as a nutrient in hydroponic culture enhances rice plant resistance to brown planthopper. Crop Prot. 2015, 67, 20–25. [Google Scholar] [CrossRef]
- Salim, M.; Saxena, R.C. Iron, silica, and aluminium stresses and varietal resistance in rice-effects on whitebacked planthopper. Crop Sci. 1992, 32, 212–219. [Google Scholar] [CrossRef]
- Waqas, M.; Shahzad, R.; Hamayun, M.; Asaf, S.; Khan, A.L.; Yun, S.; Kim, K.M.; Lee, I.J. Biochar amendment changes Jasmonic Acid levels in two rice varieties and alters their resistance to herbivory. PLoS ONE 2018, 13, e0191296. [Google Scholar] [CrossRef] [Green Version]
- Horgan, F.G.; Arida, A.; Ardestani, G.; Almazan, M.L.P. Elevated temperatures diminish the effects of a highly resistant rice variety on the brown planthopper. Sci. Rep. 2021, 11, 262. [Google Scholar] [CrossRef]
- Han, Y.Q.; Lei, W.B.; Wen, L.Z.; Hou, M.L. Silicon-mediated resistance in a susceptible rice variety to the rice leaf folder, Cnaphalocrocis medinalis Guenee (Lepidoptera: Pyralidae). PLoS ONE 2015, 10, e0120557. [Google Scholar] [CrossRef]
- Horgan, F.G.; Crisol-Martínez, E.; Almazan, M.L.P.; Romena, A.; Ramal, A.F.; Ferrater, J.B.; Bernal, C.C. Susceptibility and tolerance in hybrid and pure-line rice varieties to herbivore attack: Biomass partitioning and resource-based compensation in response to damage. Ann. Appl. Biol. 2016, 169, 200–213. [Google Scholar] [CrossRef]
- Dallagnol, L.J.; Rodrigues, F.A.; DaMatta, F.M.; Mielli, M.V.B.; Pereira, S.C. Deficiency in silicon uptake affects cytological, physiological, and biochemical events in the rice—Bipolaris oryzae interaction. Phytopathology 2010, 101, 92–104. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, F.Á.; McNally, D.J.; Datnoff, L.E.; Jones, J.B.; Labbé, C.; Benhamou, N.; Menzies, J.G.; Bélanger, R.R. Silicon enhances the accumulation of diterpenoid phytoalexins in rice: A potential mechanism for blast resistance. Phytopathology 2004, 94, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Lemraski, M.G. Silicon and phosphorus effects on blast disease incidence of rice (Oryza sativa L.). Int. J. Farming Allied Sci. 2013, 2, 1369–1374. [Google Scholar]
- Hayasaka, T.; Fujii, H.; Namai, T. Silicon content in rice seedlings to protect rice blast fungus at the nursery stage. J. Gen. Plant Pathol. 2005, 71, 169–173. [Google Scholar] [CrossRef]
- Hayasaka, T.; Fujii, H.; Ishiguro, K. The role of silicon in preventing appressorial penetration by the rice blast fungus. Phytopathology 2008, 98, 1038–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datnoff, L.E.; Raid, R.N.; Snyder, G.H.; Jones, D.B. Effect of calcium silicate on blast and brown spot intensities and yields of rice. Plant Dis. 1991, 75, 729. [Google Scholar] [CrossRef]
- Cacique, I.S.; Domiciano, G.P.; Rodrigues, F.Á.; Ribeiro do Vale, F.X. Silicon and manganese on rice resistance to blast. Bragantia 2012, 71, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Liu, B.; Zhao, T.; Xu, X.; Lin, H.; Ji, Y.; Li, Y.; Li, Z.; Lu, C.; Li, P.; et al. Silica nanoparticles protect rice against biotic and abiotic stresses. J. Nanobiotechnol. 2022, 20, 197. [Google Scholar] [CrossRef]
- Kim, S.G.; Kim, K.W.; Park, E.W.; Choi, D. Silicon-induced cell wall fortification of rice leaves: A possible cellular mechanism of enhanced host resistance to blast. Phytopathology 2002, 92, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Ye, M.; Song, Y.Y.; Long, J.; Wang, R.L.; Baerson, S.R.; Pan, Z.Q.; Zhu-Salzman, K.; Xie, J.F.; Cai, K.Z.; Luo, S.M.; et al. Priming of jasmonate-mediated antiherbivore defense responses in rice by silicon. Proc. Natl. Acad. Sci. USA 2013, 110, E3631–E3639. [Google Scholar] [CrossRef] [Green Version]
- Rizwan, M.; Atta, B.; Rizwan, M.; Sabir, A.M.; Tahir, M.; Sabar, M.; Ali, M.; Ali, M.Y. Silicon plays an effective role in integrated pest management against rice leaffolder Cnaphalocrocis medinalis Guenée (Lepidoptera: Pyralidae). Pak. J. Zool. 2021, 54, 569–576. [Google Scholar] [CrossRef]
- Ranganathan, S.; Suvarchala, V.; Rajesh, Y.; Prasad, M.S.; Padmakumari, A.P.; Voleti, S.R. Effects of silicon sources on its deposition, chlorophyll content, and disease and pest resistance in rice. Biol. Plant. 2006, 50, 713–716. [Google Scholar] [CrossRef]
- Jeer, M.; Telugu, U.M.; Voleti, S.R.; Padmakumari, A.P. Soil application of silicon reduces yellow stem borer, Scirpophaga incertulas (Walker) damage in rice. J. Appl. Entomol. 2017, 141, 189–201. [Google Scholar] [CrossRef]
- Chang, S.; Tzeng, D.; Li, C. Effects of nitrogen, calcium, and silicon nutrients on bacterial blight resistance in rice. In Rice Research for Food Security and Poverty Alleviation; Peng, S., Hardy, B., Eds.; International Rice Research Institute: Los Baños, Philippines, 2001; pp. 441–448. [Google Scholar]
- Rodrigues, F.Á.; Ribeiro do Vale, F.X.; Datnoff, L.E.; Prabhu, A.S.; Korndörfer, G.H. Effect of rice growth stages and silicon on sheath blight development. Phytopathology 2003, 93, 256–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallagnol, L.J.; Rodrigues, F.Á.; Mielli, M.V.B.; Ma, J.F.; Datnoff, L.E. Defective active silicon uptake affects some components of rice resistance to brown spot. Phytopathology 2008, 99, 116–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horgan, F.G.; Almazan, M.L.P.; Vu, Q.; Ramal, A.F.; Bernal, C.C.; Yasui, H.; Fujita, D. Unanticipated benefits and potential ecological costs associated with pyramiding leafhopper resistance loci in rice. Crop Prot. 2019, 115, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Vu, Q.; Quintana, R.; Fujita, D.; Bernal, C.C.; Yasui, H.; Medina, C.D.; Horgan, F.G. Responses and adaptation by Nephotettix virescens to monogenic and pyramided rice lines with Grh-resistance genes. Entomol. Exp. Appl. 2014, 150, 179–190. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vu, Q.; Dossa, G.S.; Mundaca, E.A.; Settele, J.; Crisol-Martínez, E.; Horgan, F.G. Combined Effects of Soil Silicon and Host Plant Resistance on Planthoppers, Blast and Bacterial Blight in Tropical Rice. Insects 2022, 13, 604. https://doi.org/10.3390/insects13070604
Vu Q, Dossa GS, Mundaca EA, Settele J, Crisol-Martínez E, Horgan FG. Combined Effects of Soil Silicon and Host Plant Resistance on Planthoppers, Blast and Bacterial Blight in Tropical Rice. Insects. 2022; 13(7):604. https://doi.org/10.3390/insects13070604
Chicago/Turabian StyleVu, Quynh, Gerbert Sylvestre Dossa, Enrique A. Mundaca, Josef Settele, Eduardo Crisol-Martínez, and Finbarr G. Horgan. 2022. "Combined Effects of Soil Silicon and Host Plant Resistance on Planthoppers, Blast and Bacterial Blight in Tropical Rice" Insects 13, no. 7: 604. https://doi.org/10.3390/insects13070604
APA StyleVu, Q., Dossa, G. S., Mundaca, E. A., Settele, J., Crisol-Martínez, E., & Horgan, F. G. (2022). Combined Effects of Soil Silicon and Host Plant Resistance on Planthoppers, Blast and Bacterial Blight in Tropical Rice. Insects, 13(7), 604. https://doi.org/10.3390/insects13070604