
Fruit Fly in a Challenging Environment: Impact of Short-Term Temperature Stress on the Survival, Development, Reproduction, and Trehalose Metabolism of Bactrocera dorsalis (Diptera: Tephritidae)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Insects
2.2. Temperature Treatments and Materials
2.3. Short-Term Temperature Stress on Egg, Larval, and Pupal Developments
2.4. Short-Term Temperature Stress on the Survival, Longevity, and Fecundity of Adults
2.5. Short-Term Temperature Stress on the Glucose, Glycogen, and Trehalose Levels of Adults
2.6. Short-Term Temperature Stress on Adult TPS Activity
2.7. Statistical Analysis
3. Results
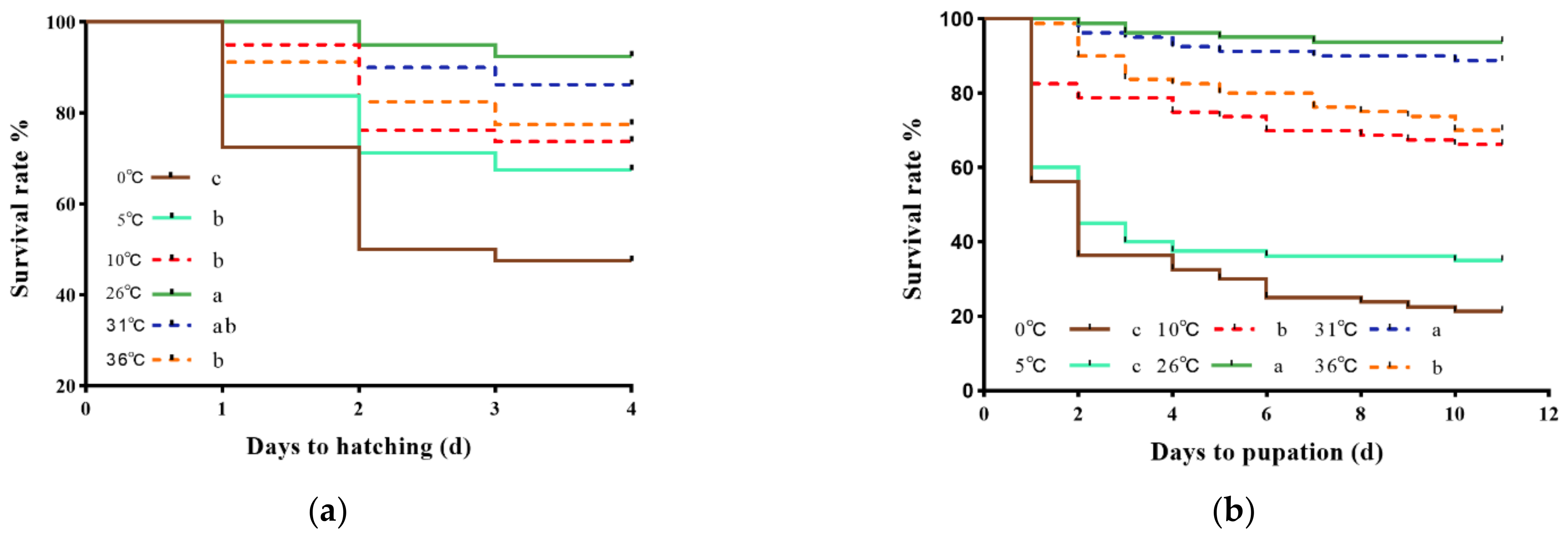
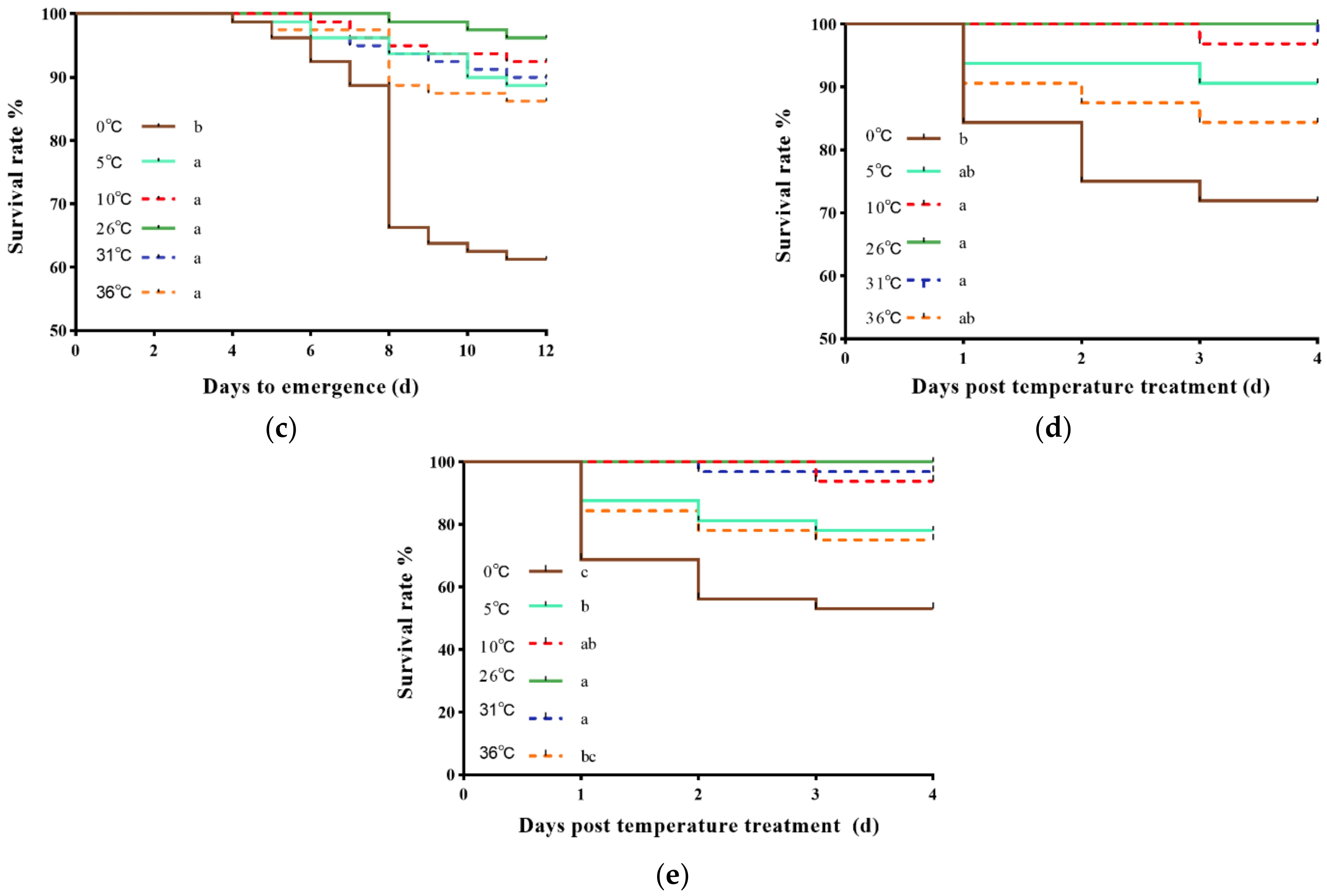
3.1. Effects of Short-Term Temperature Stress on Survival Rates
3.2. Effects of Short-Term Temperature Stress on Developmental Periods
3.3. Effects of Short-Term Temperature Stress on the Female Ratio in the Pupae Emerged and Preoviposition Period
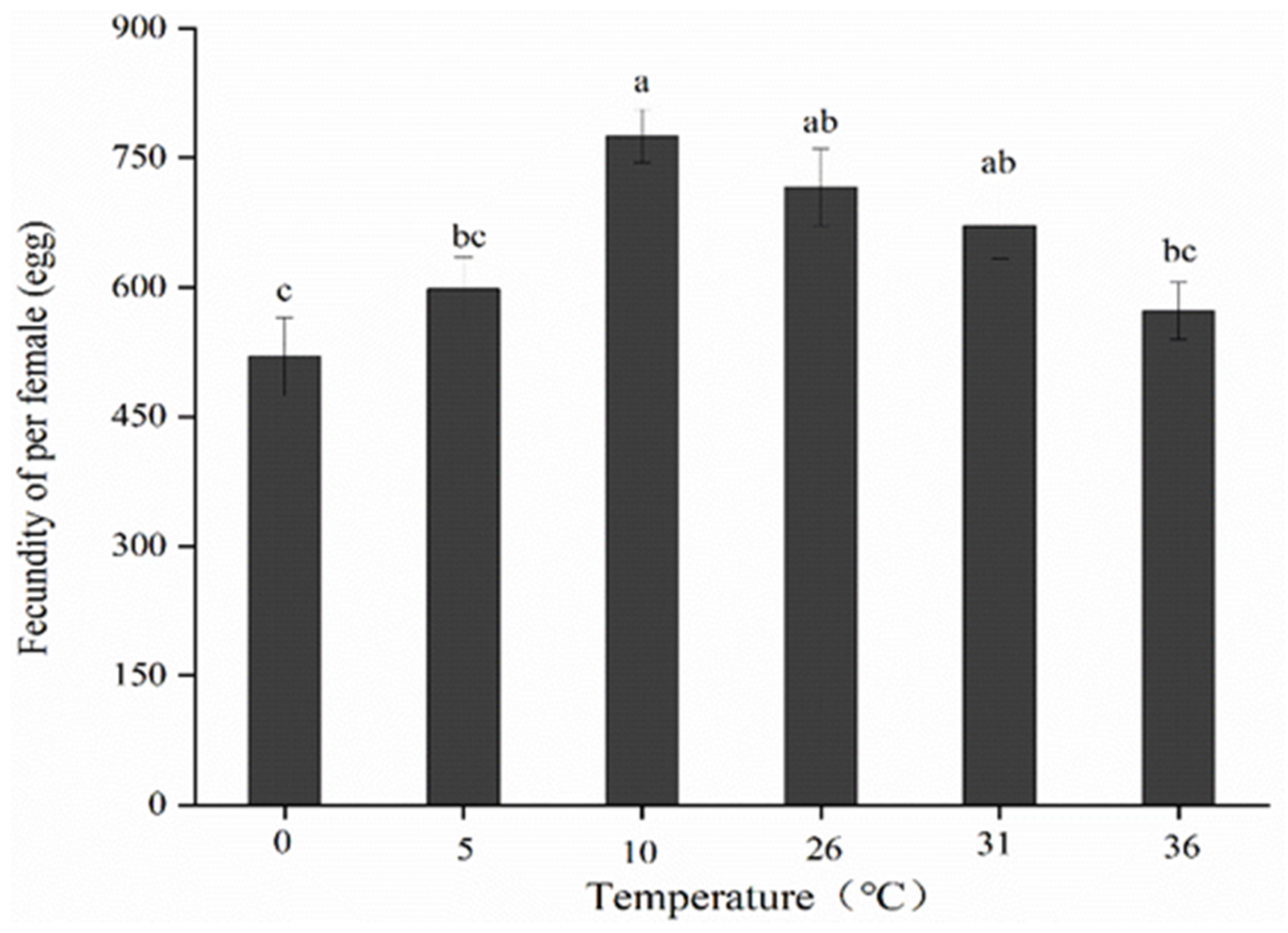
3.4. Effects of Short-Term Temperature Stress on Fecundity
3.5. Effects of Short-Term Temperature Stress on Longevity
3.6. Effects of Short-Term Temperature Stress on Low-Molecular-Weight Carbohydrate Levels
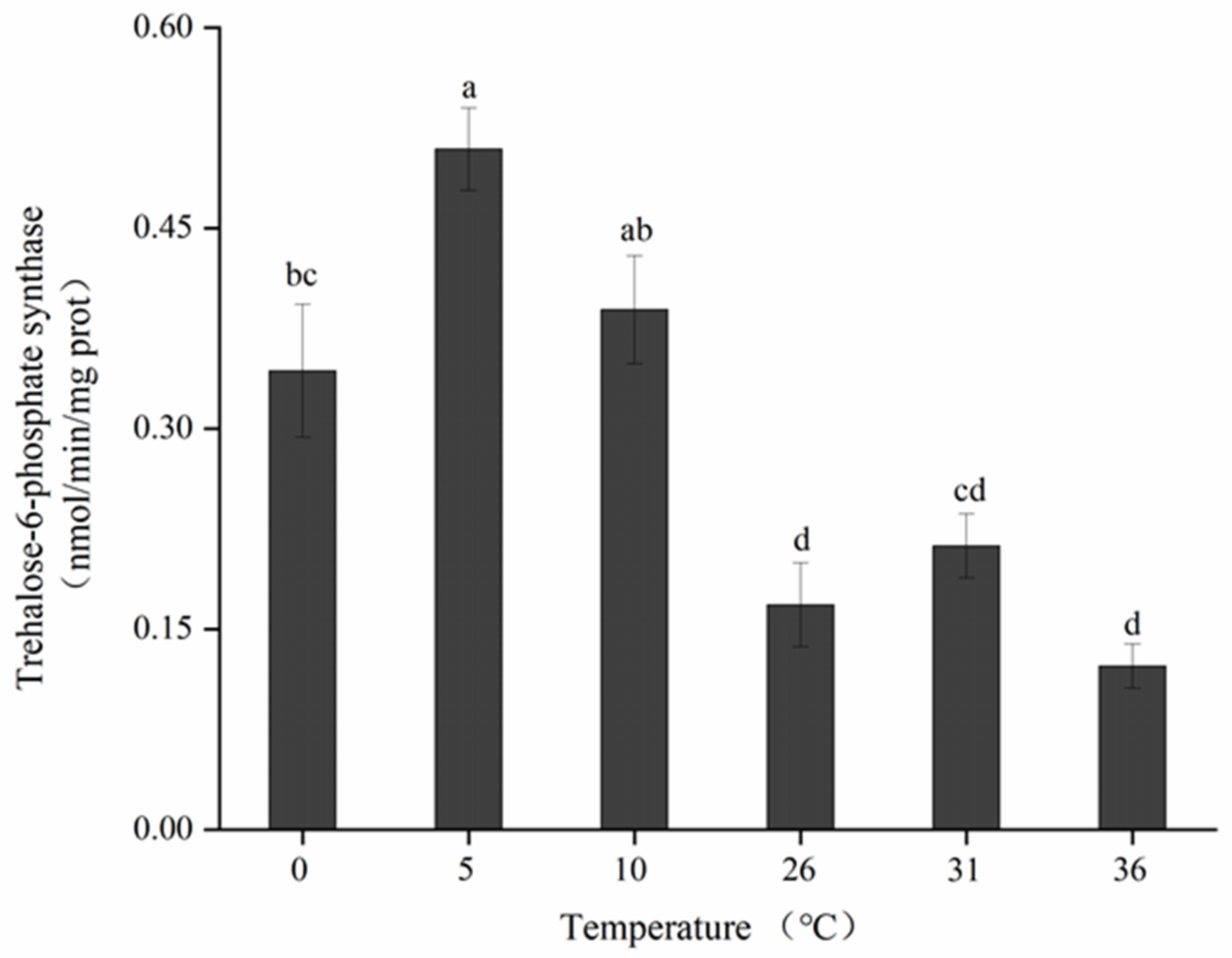
3.7. Effects of Short-Term Temperature Stress on TPS Activity of Adults
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nugnes, F.; Russo, E.; Viggiani, G.; Bernardo, U. First record of an invasive fruit fly belonging to Bactrocera dorsalis complex (Diptera: Tephritidae) in Europe. Insects 2018, 9, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, A.E.A.; Kriticos, D.J.; Leriche, A. The current and future potential geographical distribution of the oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae). Bull. Entomol. Res. 2007, 97, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.Y.; Kong, Q.; Xiao, C.; Chen, B.; Yang, S.S.; Zhang, J.B.; Sun, W.; Li, Z.Y. Influence of temperature on development, survival and fecundity of Bactrocera dorsalis Hendel. J. Huazhong Agric. Univ. 2005, 6, 588–591. [Google Scholar]
- Armstrong, J.W. Quarantine security of bananas at harvest maturity against Mediterranean and oriental fruit flies (Diptera: Tephritidae) in Hawaii. J. Econ. Entomol. 2001, 94, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Bentz, B.J.; Régnière, J.; Fettig, C.J.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Seybold, S.J. Climate change and bark beetles of the Western United States and Canada: Direct and indirect effects. BioScience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Gunderson, A.R.; Stillman, J.H. Plasticity in thermal tolerance has limited potential to buffer ectotherms from global warming. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150401. [Google Scholar] [CrossRef]
- Damien, M.; Tougeron, K. Prey–predator phenological mismatch under climate change. Curr. Opin. Insect Sci. 2019, 35, 60–68. [Google Scholar] [CrossRef]
- Abarca, M.; Spahn, R. Direct and indirect effects of altered temperature regimes and phenological mismatches on insect populations. Curr. Opin. Insect Sci. 2021, 47, 67–74. [Google Scholar] [CrossRef]
- Colinet, H.; Sinclair, B.J.; Vernon, P.; Renault, D. Insects in fluctuating thermal environments. Annu. Rev. Entomol. 2015, 60, 123–140. [Google Scholar] [CrossRef] [Green Version]
- Bale, J.S.; Hayward, S.A.L. Insect overwintering in a changing climate. J. Exp. Biol. 2010, 213, 980–994. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.J.; Bauer, L.S.; Van Driesche, R.V.; Schmude, J.M.; Petrice, T.; Chandler, J.L.; Elkinton, J. Effects of extreme low winter temperatures on the overwintering survival of the introduced larval parasitoids Spathius galinae and Tetrastichus planipennisi: Implications for biological control of emerald Ash borer in North America. J. Econ. Entomol. 2020, 113, 1145–1151. [Google Scholar] [CrossRef] [Green Version]
- Yocum, G.D.; Žďárek, J.; Joplin, K.H.; Lee, R.E.; Smith, D.C.; Manter, K.D.; Denlinger, D.L. Alteration of the eclosion rhythm and eclosion behavior in the flesh fly, Sarcophaga crassipalpis, by low and high temperature stress. J. Insect Physiol. 1994, 40, 13–21. [Google Scholar] [CrossRef]
- Al-Behadili, F.; Bilgi, V.; Li, J.; Wang, P.; Taniguchi, M.; Agarwal, M.; Ren, Y.L.; Xu, W. Cold response of the Mediterranean fruit fly (Ceratitis capitata) on a lab diet. Insects 2019, 10, 48. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Li, S.; Zhang, B.; Li, H.G.; Wang, F.H.; Zheng, C.Y. Effects of extreme high temperature on survival rate, reproduction, trehalose and sorbitol of Frankliniella occidentalis. Sci. Agric. Sin. 2016, 49, 2310–2321. [Google Scholar]
- Teets, N.M.; Denlinger, D.L. Physiological mechanisms of seasonal and rapid cold-hardening in insects. Physiol. Entomol. 2013, 38, 105–116. [Google Scholar] [CrossRef]
- Hu, J.; Chen, B.; Li, Z. Thermal plasticity is related to the hardening response of heat shock protein expression in two Bactrocera fruit flies. J. Insect Physiol. 2014, 67, 105–113. [Google Scholar] [CrossRef]
- Shukla, E.; Thorat, L.J.; Nath, B.B.; Gaikwad, S.M. Insect trehalase: Physiological significance and potential applications. Glycobiology 2014, 25, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Wharton, D.A. Cold tolerance of New Zealand alpine insects. J. Insect Physiol. 2011, 57, 1090–1095. [Google Scholar] [CrossRef]
- Hamedi, N.; Moharramipour, S.; Barzegar, M. Temperature-dependent chemical components accumulation in Hippodamia variegate (Coleoptera: Coccinellidae) during overwintering. Environ. Entomol. 2013, 42, 375–380. [Google Scholar] [CrossRef]
- Crowe, J.H.; Crowe, L.M.; Chapman, D. Preservation of membranes in anhydrobiotic organisms: The role of trehalose. Science 1984, 223, 701–703. [Google Scholar] [CrossRef]
- Feofilova, E.P.; Usov, A.I.; Mysyakina, I.S.; Kochkina, G.A. Trehalose: Chemical structure, biological functions, and practical application. Mikrobiologiia 2014, 83, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Thorat, L.J.; Gaikwad, S.M.; Nath, B.B. Trehalose as an indicator of desiccation stress in Drosophila melanogaster larvae: A potential marker of anhydrobiosis. Biochem. Biophys. Res. Commun. 2012, 419, 638–642. [Google Scholar] [CrossRef] [PubMed]
- Jagdale, G.B.; Grewal, P.S.; Salminen, S.O. Both heat-shock and cold-shock influence trehalose metabolism in an entomopathogenic nematode. J. Parasitol. 2005, 91, 988–994. [Google Scholar] [CrossRef] [PubMed]
- Ghaedi, B.; Andrew, N.R. The physiological consequences of varied heat exposure events in adult Myzus persicae: A single prolonged exposure compared to repeated shorter exposures. PeerJ 2016, 4, e2290. [Google Scholar] [CrossRef] [Green Version]
- Almeida, V.M.; Marana, S.R. Optimum temperature may be a misleading parameter in enzyme characterization and application. PLoS ONE 2019, 14, e0212977. [Google Scholar] [CrossRef] [Green Version]
- González-Tokman, D.; Córdoba-Aguilar, A.; Dáttilo, W.; Lira-Noriega, A.; Sánchez-Guillén, R.A.; Villalobos, F. Insect responses to heat: Physiological mechanisms, evolution and ecological implications in a warming world. Biol. Rev. 2020, 95, 802–821. [Google Scholar] [CrossRef]
- Lum, P.Y.; Chino, H. Trehalose, the insect blood sugar, inhibits loading of diacylglycerol by lipophorin from the fat body in locusts. Biochem. Biophys. Res. Commun. 1990, 172, 588–594. [Google Scholar] [CrossRef]
- Tang, B.; Wang, S.; Wang, S.G.; Wang, H.J.; Zhang, J.Y.; Cui, S.Y. Invertebrate Trehalose-6-Phosphate synthase gene: Genetic architecture, biochemistry, physiological function, and potential applications. Front. Physiol. 2018, 9, 30. [Google Scholar] [CrossRef] [Green Version]
- Mitsumasu, K.; Kanamori, Y.; Fujita, M.; Iwata, K.; Tanaka, D.; Kikuta, S.; Watanabe, M.; Cornette, R.; Okuda, T.; Kikawada, T. Enzymatic control of anhydrobiosis-related accumulation of trehalose in the sleeping chironomid, Polypedilum vanderplanki. FEBS J. 2010, 277, 4215–4228. [Google Scholar] [CrossRef] [Green Version]
- Murphy, T.A.; Wyatt, G.R. The enzymes of glycogen and trehalose synthesis in silk moth fat body. J. Biol. Chem. 1965, 240, 1500–1508. [Google Scholar] [CrossRef]
- Mendes, V.; Acebrón-García-de-Eulate, M.; Verma, N.; Blaszczyk, M.; Dias, M.V.B.; Blundell, T.L. Mycobacterial OtsA structures unveil substrate preference mechanism and allosteric regulation by 2-Oxoglutarate and 2-Phosphoglycerate. mBio 2019, 10, e02272-19. [Google Scholar] [CrossRef] [Green Version]
- Kern, C.; Wolf, C.; Bender, F.; Berger, M.; Noack, S.; Schmalz, S.; Ilg, T. Trehalose-6-phosphate synthase from the cat flea Ctenocephalides felis and Drosophila melanogaster: Gene identification, cloning, heterologous functional expression and identification of inhibitors by high throughput screening. Insect Mol. Biol. 2012, 21, 456–471. [Google Scholar] [CrossRef]
- Li, Y.; Hao, Y.J.; Zhang, Y.J.; Si, F.L.; Chen, B. Cloning, bioinformatic analysis and diapause-related expression of trehalose-6- phosphate synthase gene from the onion maggot, Delia antiqua (Diptera: Anthomyiidae). Acta Entomol. Sin. 2013, 56, 329–338. [Google Scholar]
- Kim, Y.; Lee, D.W.; Jung, J.K. Rapid cold-hardening of a subtropical species, Maruca vitrata (Lepidoptera: Crambidae), accompanies hypertrehalosemia by upregulating trehalose-6-phosphate synthase. Environ. Entomol. 2017, 46, 1432–1438. [Google Scholar] [CrossRef]
- Clark, M.S.; Thorne, M.A.; Purać, J.; Burns, G.; Hillyard, G.; Popović, Ž.D.; Grubor-Lajšić, G.; Worland, M.R. Surviving the cold: Molecular analyses of insect cryoprotective dehydration in the Arctic springtail Megaphorura arctica (Tullberg). BMC Genom. 2009, 10, 328. [Google Scholar] [CrossRef] [Green Version]
- Samayoa, A.C.; Choi, K.S.; Wang, Y.S.; Hwang, S.Y.; Huang, Y.B.; Ahn, J.J. Thermal effects on the development of Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) and model validation in Taiwan. Phytoparasitica 2018, 46, 365–376. [Google Scholar] [CrossRef]
- Duyck, P.F.; Sterlin, J.F.; Quilici, S. Survival and development of different life stages of Bactrocera zonata (Diptera: Tephritidae) reared at five constant temperatures compared to other fruit fly species. Bull. Entomol. Res. 2004, 94, 89–93. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.L. The Influence of Temperature Stress, Soil Moisture and Different Hosts on Population of Bactrocera dorsalis (Hendel). Master’s Thesis, Southwest University, Chongqing, China, 2009. [Google Scholar]
- Dongmo, M.A.K.; Fiaboe, K.K.M.; Kekeunou, S.; Nanga, S.N.; Kuate, A.F.; Tonnang, H.E.Z.; Gnanvossou, D.; Hanna, R. Temperature-based phenology model to predict the development, survival, and reproduction of the oriental fruit fly Bactrocera dorsalis. J. Therm. Biol. 2021, 97, 102877. [Google Scholar]
- Yuan, S.Y.; Xiao, C.; Li, Z.Y.; Zhu, J.Y. A study on laboratory rearing techniques for Bactrocera dorsalis (Hendal). Acta Agric. Univ. Jiangxiensis 2003, 4, 577–580. [Google Scholar]
- Yuan, S.Y.; Kong, Q.; Xiao, C.; Yang, S.S.; Sun, W.; Zhang, J.B.; Li, Z.Y. lntroduction to two kinds of artificial diets for mass rearing of Adult Bactrocera dorsalis (Hendal). J. Huazhong Agric. Univ. 2006, 4, 371–374. [Google Scholar]
- Pieterse, W.; Manrakhan, A.; Terblanche, J.S.; Addison, P. Comparative demography of Bactrocera dorsalis (Hendel) and Ceratitis capitata (Wiedemann) (Diptera: Tephritidae) on deciduous fruit. Bull. Entomol. Res. 2020, 110, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Osnes, T.; Sandstad, O.; Skar, V.; Osnes, M.; Kierulf, P. Total protein in common duct bile measured by acetonitrile precipitation and a micro bicinchoninic acid (BCA) method. Scand. J. Clin. Lab. Investig. 1993, 53, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Geng, S.B.; Hou, H.L.; Zhang, J.; Qiao, L.; Jin, Y.L.; Yin, J.; Wang, G.J. Effects of temperature on longevity and fecundity of Scopula subpunctaria. Plant Prot. 2022, 48, 118–125. [Google Scholar]
- Maurizio, M.; Fronte1, B.; Sagona1, S.; Carrozza, M.L.; Forzan1, M.; Pizzurro1, F.; Bibbiani1, C.; Miragliotta, V.; Abramo1, F.; Millanta1, F.; et al. Effect of 1,3-1,6 β-Glucan on natural and experimental deformed wing virus infection in newly emerged honeybees (Apis mellifera ligustica). PLoS ONE 2016, 11, e166297. [Google Scholar]
- Colinet, H.; Rinehart, J.P.; Yocum, G.D.; Greenlee, K.J. Mechanisms underpinning the beneficial effects of fluctuating thermal regimes in insect cold tolerance. J. Exp. Biol. 2018, 221, jeb164806. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.Y.; Gu, X.P.; Peng, X.Q.; Tao, M.; Peng, L.; Chen, G.H.; Zhang, X.M. Effect of short-term low temperature on the growth, development, and reproduction of Bactrocera tau (Diptera: Tephritidae) and Bactrocera cucurbitae. J. Econ. Entomol. 2020, 113, 2141–2149. [Google Scholar] [CrossRef]
- Manrakhan, A.; Daneel, J.H.; Stephen, P.R.; Hattingh, V. Cold tolerance of immature stages of Ceratitis capitata and Bactrocera dorsalis (Diptera: Tephritidae). J. Econ. Entomol. 2022, 115, 482–492. [Google Scholar] [CrossRef]
- Xiao, G.J.; Zeng, L.; Li, Q.; Lu, Y.Y. Cold hardiness of palm leaf beetle, Brontispa longissima. Chinese. Bull. Entomol. 2006, 43, 527–530. [Google Scholar]
- Hou, B.H.; Zhang, R.J. Supercooling capacity of the Oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Acta Entomol. Sin. 2007, 6, 638–643. [Google Scholar]
- Danjuma, S.; Thaochan, N.; Permkam, S.; Satasook, C. Effect of temperature on the development and survival of immature stages of the carambola fruit fly, Bactrocera carambolae, and the Asian papaya fruit fly, Bactrocera papayae, reared on guava diet. J. Insect Sci. 2014, 14, 126. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Chen, G.Q.; Meng, Y.Q. Survival ability of Bactrocera dorsalis under extreme temperature conditions in Taizhou. S. China Fruits 2012, 41, 50–52. [Google Scholar]
- Wei, L.; Ou, H.D.; Wang, X.Q.; Yu, X.F.; Yang, M.F. Effect of cold acclimation storage on adult longevity and reproduction of Habrobracon hebetor. J. Mt. Agric. Biol. 2022, 41, 76–81. [Google Scholar]
- Jervis, M.A.; Boggs, C.L.; Ferns, P.N. Egg maturation strategy and its associated trade-offs: A synthesis focusing on Lepidoptera. Ecol. Entomol. 2005, 30, 359–375. [Google Scholar] [CrossRef]
- Yu, L.P.; Xu, N.C. The relationship between the biological characteristics of citrus fruit flies and temperature. China Horticult. Abstr. 2012, 28, 19–21. [Google Scholar]
- Fleurat-Lessard, F.; Dupuis, V. Comparative analysis of upper thermal tolerance and CO2 production rate during heat shock in two different European strains of Sitophilus zeamais (Coleoptera: Curculionidae). J. Stored Prod. Res. 2010, 46, 20–27. [Google Scholar] [CrossRef]
- Bemani, M.; Izadi, H.; Mahdian, K.; Khani, A.; Amin Samih, M.A. Study on the physiology of diapause, cold hardiness and supercooling point of overwintering pupae of the pistachio fruit hull borer, Arimania comaroffi. J. Insect Physiol. 2012, 58, 897–902. [Google Scholar] [CrossRef]
- Wu, H.B.; Gong, Q.T.; Chen, Z.Z.; Jiang, L.L.; Gong, Y.; Xu, Y.Y.; Sun, R.H. Bioefficacy of the combined application of entomopathogenic nematodes and thiamethoxam and its effects on the protective and detoxification enzyme activities in Bradysia odoriphaga (Diptera: Sciaridae) larvae. Acta Entomol. Sin. 2018, 61, 851–859. [Google Scholar]
- Qian, L.Y.; Kou, X.Y.; Dong, Z.H.; Chang, Y.L.; Zhou, Z.J. Effects of different temperature regimes on the expression of the trehalose-6-phosphate synthase gene and haemolymph sugar content in Gampsocleis gratiosa. Chin. J. Appl. Entomol. 2017, 54, 56–67. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | Developmental Period (d) | ||
|---|---|---|---|
| Egg | Larva | Pupa | |
| 26 | 1.56 ± 0.07 d | 8.15 ± 0.16 d | 10.06 ± 0.03 b |
| 0 | 3.11 ± 0.17 a | 10.11 ± 0.34 a | 11.25 ± 0.16 a |
| 5 | 2.68 ± 0.05 b | 9.68 ± 0.06 ab | 11.08 ± 0.03 a |
| 10 | 2.35 ± 0.14 bc | 9.39 ± 0.13 ab | 10.28 ± 0.07 b |
| 31 | 1.82 ± 0.04 d | 8.43 ± 0.07 cd | 10.01 ± 0.14 b |
| 36 | 2.23 ± 0.07 c | 9.19 ± 0.08 bc | 10.25 ± 0.07 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, C.; Zhao, R.; Zhou, W.; Pan, Y.; Tian, H.; Yin, Z.; Chen, W. Fruit Fly in a Challenging Environment: Impact of Short-Term Temperature Stress on the Survival, Development, Reproduction, and Trehalose Metabolism of Bactrocera dorsalis (Diptera: Tephritidae). Insects 2022, 13, 753. https://doi.org/10.3390/insects13080753
Yu C, Zhao R, Zhou W, Pan Y, Tian H, Yin Z, Chen W. Fruit Fly in a Challenging Environment: Impact of Short-Term Temperature Stress on the Survival, Development, Reproduction, and Trehalose Metabolism of Bactrocera dorsalis (Diptera: Tephritidae). Insects. 2022; 13(8):753. https://doi.org/10.3390/insects13080753
Chicago/Turabian StyleYu, Chun, Runa Zhao, Wei Zhou, Yingna Pan, Hui Tian, Zhengyan Yin, and Wenlong Chen. 2022. "Fruit Fly in a Challenging Environment: Impact of Short-Term Temperature Stress on the Survival, Development, Reproduction, and Trehalose Metabolism of Bactrocera dorsalis (Diptera: Tephritidae)" Insects 13, no. 8: 753. https://doi.org/10.3390/insects13080753
APA StyleYu, C., Zhao, R., Zhou, W., Pan, Y., Tian, H., Yin, Z., & Chen, W. (2022). Fruit Fly in a Challenging Environment: Impact of Short-Term Temperature Stress on the Survival, Development, Reproduction, and Trehalose Metabolism of Bactrocera dorsalis (Diptera: Tephritidae). Insects, 13(8), 753. https://doi.org/10.3390/insects13080753