


Competitive Plant-Mediated and Intraguild Predation Interactions of the Invasive Spodoptera frugiperda and Resident Stemborers Busseola fusca and Chilo partellus in Maize Cropping Systems in Kenya

 , ,
, ,  , , ,
, , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Insects
2.3. Plants
2.4. Competitive Plant-Mediated Interactions
2.5. Predator–Prey Interactions
2.6. Field Occurrence of Spodoptera frugiperda and Stemborers
2.7. Data Analysis
3. Results
3.1. Competitive Plant-Mediated Effects
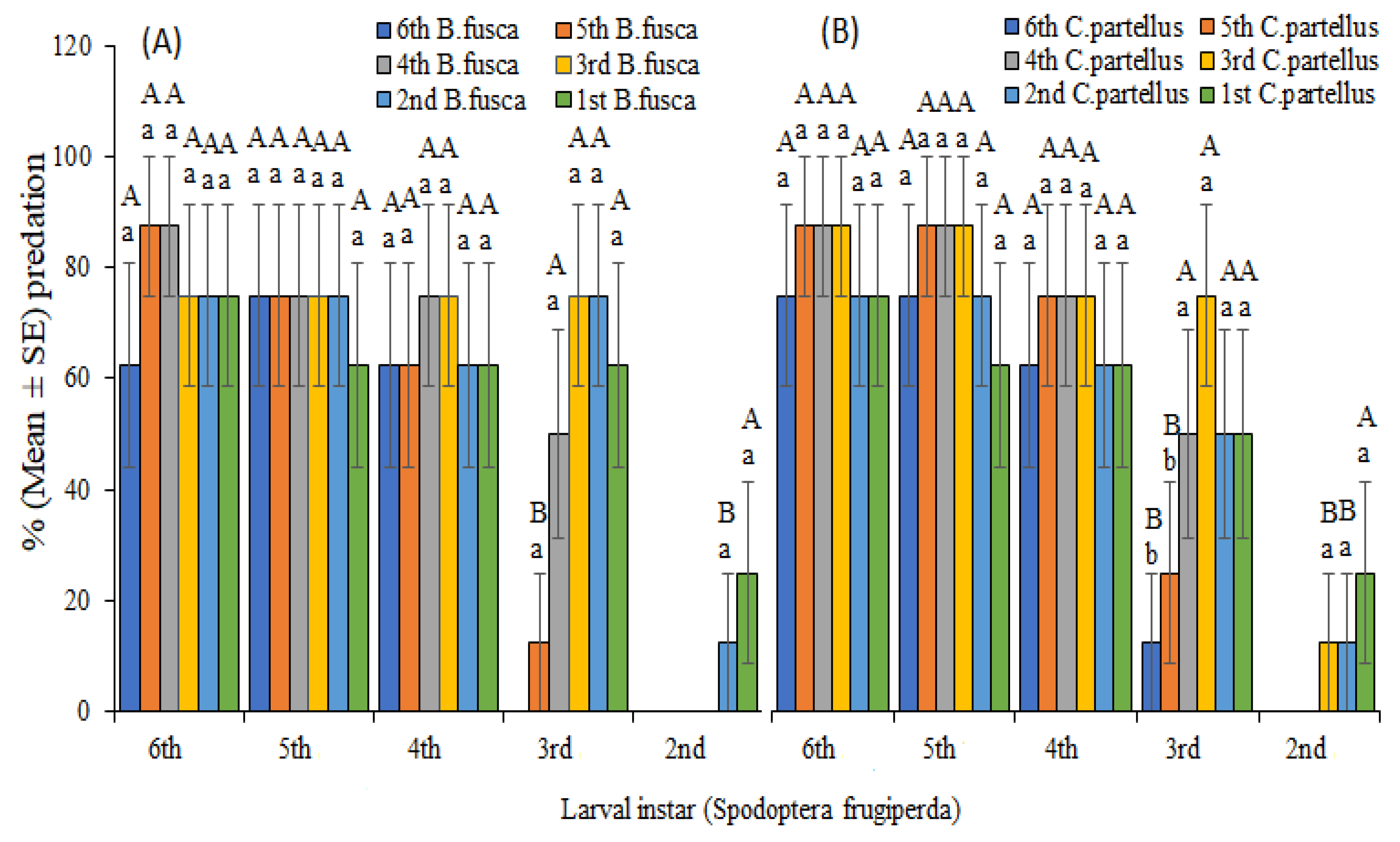
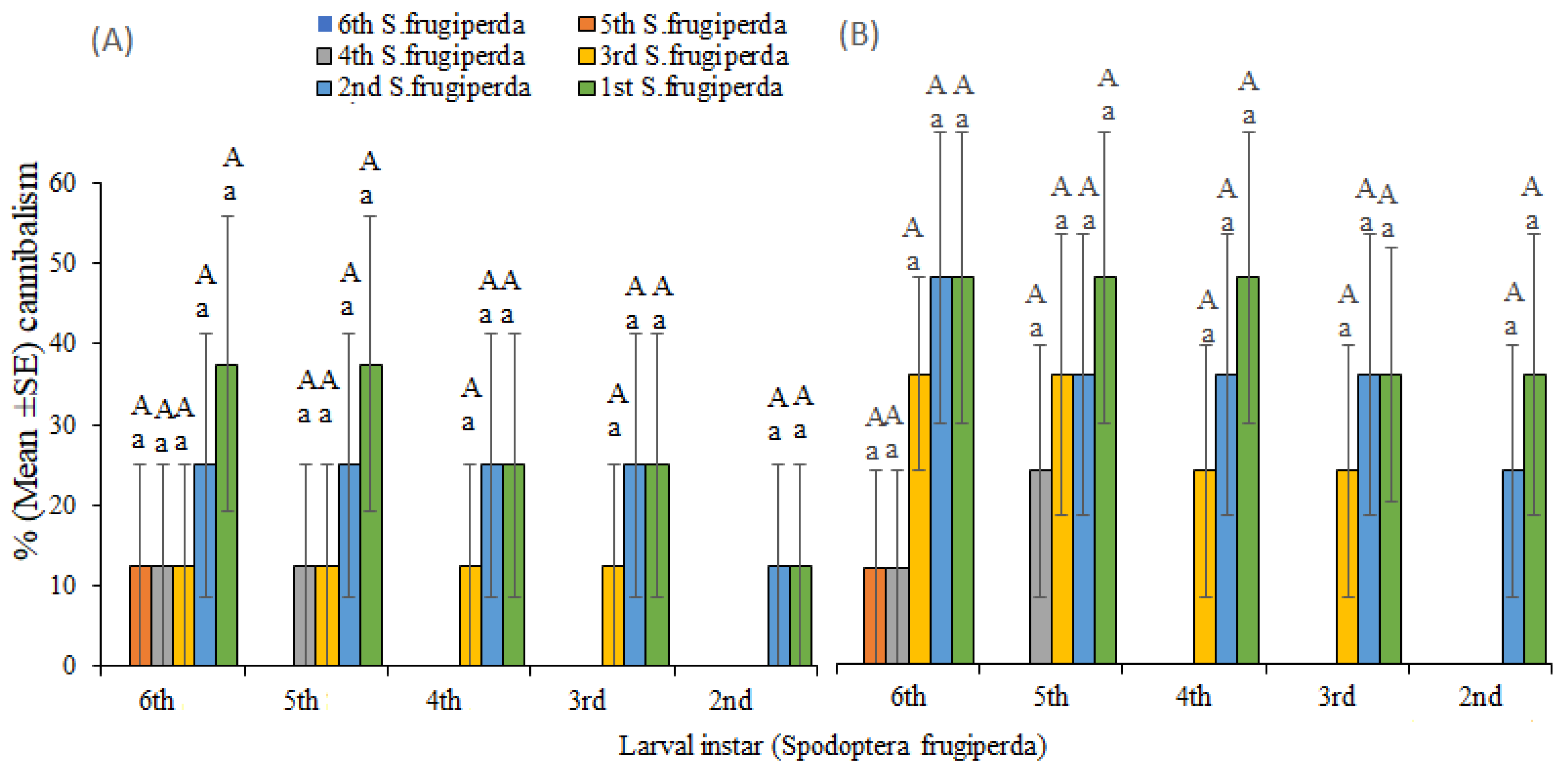
3.2. Predator–Prey Interactions
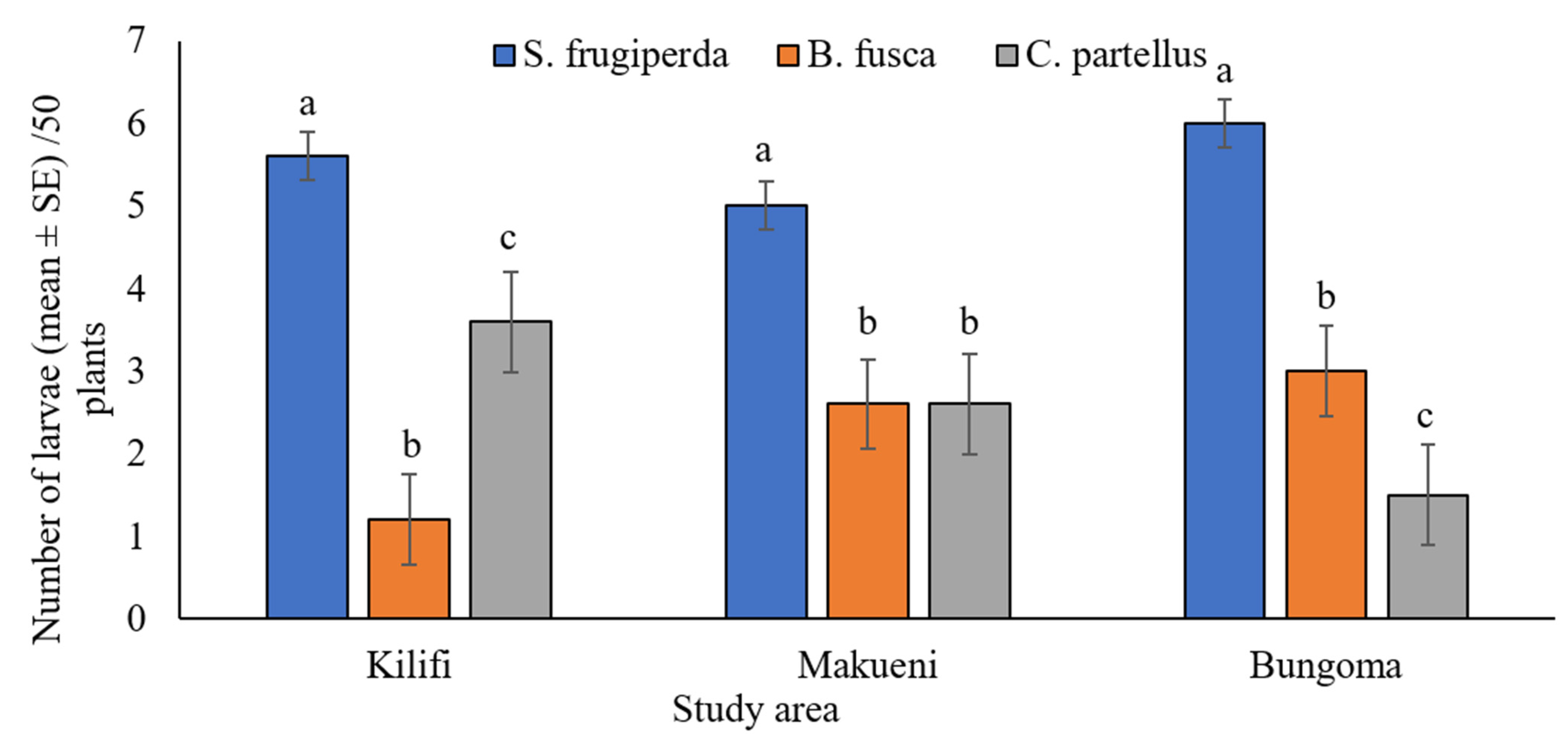
3.3. Co-Occurrence of Spodoptera frugiperda and Stemborers in Maize Fields
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [PubMed]
- De Groote, H.; Kimenju, S.C.; Munyua, B.; Palmas, S.; Kassie, M.; Bruce, A. Spread and impact of fall armyworm (Spodoptera frugiperda J.E. Smith) in maize production areas of Kenya. Agric. Ecosyst. Environ. 2020, 292, 106804. [Google Scholar] [CrossRef] [PubMed]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall armyworm: Impacts and implications for Africa. Outlooks on pest management. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Kfir, R. Natural control of the cereal stemborers Busseola fusca and Chilo partellus in South Africa. Insect Sci. Appl. 1997, 17, 61–67. [Google Scholar] [CrossRef]
- Sokame, B.M.; Obonyo, J.; Sammy, E.M.; Mohamed, S.A.; Subramanian, S.; Kilalo, D.C.; Juma, G.; Calatayud, P.-A. Impact of the exotic fall armyworm on larval parasitoids associated with the lepidopteran maize stemborers in Kenya. BioControl 2020, 66, 193–204. [Google Scholar] [CrossRef]
- Ntiri, E.S.; Calatayud, P.-A.; Berg, J.V.D.; Le Ru, B.P. Spatio-temporal interactions between maize lepidopteran stemborer communities and possible implications from the recent invasion of Spodoptera frugiperda (Lepidoptera: Noctuidae) in Sub-Saharan Africa. Environ. Entomol. 2019, 48, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Bentivenha, J.P.; Montezano, D.G.; Hunt, T.E.; Baldin, E.L.; Peterson, J.A.; Victor, V.S.; Pannuti, L.E.; Vélez, A.M.; Paula-Moraes, S.V. Intraguild interactions and behavior of Spodoptera frugiperda and Helicoverpa spp. on maize. Pest Manag. Sci. 2017, 73, 2244–2251. [Google Scholar] [CrossRef]
- Pannuti, L.E.R.; Baldin, E.L.L.; Hunt, T.E.; Paula-Moraes, S.V. On-plant larval movement and feeding behavior of fall armyworm (lepidoptera: Noctuidae) on reproductive corn stages. Environ. Entomol. 2016, 45, 192–200. [Google Scholar] [CrossRef]
- Hailu, G.; Niassy, S.; Bässler, T.; Ochatum, N.; Studer, C.; Salifu, D.; Agbodzavu, M.K.; Khan, Z.R.; Midega, C.; Subramanian, S. Could fall armyworm, Spodoptera frugiperda (J. E. Smith) invasion in Africa contribute to the displacement of cereal stemborers in maize and sorghum cropping systems. Int. J. Trop. Insect Sci. 2021, 41, 1753–1762. [Google Scholar] [CrossRef]
- Kaplan, I.; Denno, R.F. Interspecific interactions in phytophagous insects revisited: A quantitative assessment of competition theory. Ecol. Lett. 2007, 10, 977–994. [Google Scholar] [CrossRef]
- Denno, R.F.; McClure, M.S.; Ott, J.R. Interactions in Resurrected. Annu. Rev. Entomol. 1995, 40, 297–331. [Google Scholar] [CrossRef]
- Liu, S.-S.; De Barro, P.J.; Xu, J.; Luan, J.-B.; Zang, L.-S.; Ruan, Y.-M.; Wan, F.-H. Asymmetric mating interactions drive widespread invasion and displacement in a whitefly. Science 2007, 318, 1769–1772. [Google Scholar] [CrossRef] [PubMed]
- Ekesi, S.; Billah, M.K.; Nderitu, P.W.; Lux, S.A.; Rwomushana, I. Evidence for competitive displacement of Ceratitis cosyra by the invasive fruit fly Bactrocera invadens (Diptera: Tephritidae) on mango and mechanisms contributing to the displacement. J. Econ. Entomol. 2009, 102, 981–991. [Google Scholar] [CrossRef] [PubMed]
- Ofomata, V.; Overholt, W.; Huis, A.; Egwuatu, R.; Ngi-Song, A. Niche overlap and interspecific association between Chilo partellus and Chilo orichalcociliellus on the Kenya coast. Entomol. Exp. Appl. 1999, 93, 141–148. [Google Scholar] [CrossRef]
- Overholt, W.A.; Ogedah, K.; Lammers, P. Distribution and sampling of Chilo partellus (Lepidoptera: Pyralidae) in maize and sorghum on the Kenya coast. Bull. Entomol. Res. 1994, 84, 367–378. [Google Scholar] [CrossRef]
- Warui, C.M.; Kuria, J.N. Population incidence and the control of maize stalk-borers Chilo partellus (Swinh.), C. orichalcociliellus Strand and Sesamia calamistis Hmps, in Coast Province, Kenya. Int. J. Trop. Insect Sci. 1983, 4, 11–18. [Google Scholar] [CrossRef]
- Song, Y.; Yang, X.; Zhang, H.; Zhang, D.; He, W.; Wyckhuys, K.A.G.; Wu, K. Interference competition and predation between invasive and native herbivores in maize. J. Pest Sci. 2021, 94, 1053–1063. [Google Scholar] [CrossRef]
- Zhao, J.; Hoffmann, A.; Jiang, Y.; Xiao, L.; Tan, Y.; Zhou, C.; Bai, L. Competitive interactions of a new invader (Spodoptera frugiperda) and indigenous species (Ostrinia furnacalis) on maize in China. J. Pest Sci. 2021, 95, 159–168. [Google Scholar] [CrossRef]
- Courchamp, F.; Fournier, A.; Bellard, C.; Bertelsmeier, C.; Bonnaud, E.; Jeschke, J.M.; Russell, J.C. Invasion Biology: Specific Problems and Possible Solutions. Trends Ecol. Evol. 2017, 32, 13–22. [Google Scholar] [CrossRef]
- Chapman, J.W.; Williams, T.; Escribano, A.; Caballero, P.; Cave, R.D.; Goulson, D. Fitness consequences of cannibalism in the fall armyworm, Spodoptera frugiperda. Behav. Ecol. 1999, 10, 298–303. [Google Scholar] [CrossRef] [Green Version]
- Njuguna, E.; Nethononda, P.; Maredia, K.; Mbabazi, R.; Kachapulula, P.; Rowe, A.; Ndolo, D. Experiences and Perspectives on Spodoptera frugiperda (Lepidoptera: Noctuidae) Management in Sub-Saharan Africa. J. Integr. Pest Manag. 2021, 12, 7. [Google Scholar] [CrossRef]
- Polis, G.A. The Evolution and Dynamics of Intraspecific Predation. Annu. Rev. Ecol. Syst. 1981, 12, 225–251. [Google Scholar] [CrossRef]
- Bonhof, M.; Overholt, W. Impact of solar radiation rainfall and cannibalism on disappearance of maize stemborers in Kenya. Insect Sci. Appl. 2001, 21, 403–407. [Google Scholar] [CrossRef]
- Schellhorn, N.A.; Andow, D.A. Cannibalism and Interspecific Predation: Role of Oviposition Behavior. Ecol. Appl. 1999, 9, 418–428. [Google Scholar] [CrossRef]
- Raffa, K.F. Influence of Host Plant on Deterrence by Azadirachtin of Feeding by Fall Armyworm Larvae (Lepidoptera: Noctuidae). J. Econ. Entomol. 1987, 80, 384–387. [Google Scholar] [CrossRef]
- Miura, K.; Ohgushi, T. A native predator affects the indirect interaction between exotic herbivorous insects on an invaded plant. J. Plant Interact. 2011, 6, 175–176. [Google Scholar] [CrossRef]
- Persson, L. Predator-Mediated Competition in Prey Refuges: The Importance of Habitat Dependent Prey Resources. Oikos 1993, 68, 12. [Google Scholar] [CrossRef]
- Karban, R. The ecology and evolution of induced resistance against herbivores. Funct. Ecol. 2011, 25, 339–347. [Google Scholar] [CrossRef]
- Gordy, J.W.; Leonard, B.R.; Blouin, D.; Davis, J.A.; Stout, M.J. Comparative effectiveness of potential elicitors of plant resistance against Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) in four crop plants. PLoS ONE 2015, 10, e0136689. [Google Scholar] [CrossRef]
- Chuang, W.-P.; Ray, S.; Acevedo, F.E.; Peiffer, M.; Felton, G.W.; Luthe, D.S. Herbivore cues from the fall armyworm (Spodoptera frugiperda) larvae trigger direct defenses in maize. Mol. Plant-Microbe Interact. 2014, 27, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Peiffer, M.; Felton, G.W. The host plant as a factor in the synthesis and secretion of salivary glucose oxidase in larval Helicoverpa zea. Arch. Insect Biochem. Physiol. 2005, 58, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Dicke, M. Induced Responses to Herbivory by R. Karban and I.T. Baldwin. Trends Ecol. Evol. 1998, 13, 83. [Google Scholar] [CrossRef]
- Ray, S.; Alves, P.C.; Ahmad, I.; Gaffoor, I.; Acevedo, F.E.; Peiffer, M.; Jin, S.; Han, Y.; Shakeel, S.; Felton, G.W.; et al. Turnabout is fair play: Herbivory-induced plant chitinases excreted in fall armyworm frass suppress herbivore defenses in maize. Plant Physiol. 2016, 171, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Denno, R.F.; Kaplan, I. Plant-mediated interactions in herbivorous insects: Mechanisms, symmetry, and challenging the paradigms of competition past. Ecol. Communities Plant Mediat. Indirect. Interact. Webs 2007, 19–49. [Google Scholar] [CrossRef]
- Onyango, F.O.; Ochieng‘-Odero, J.P.R. Continuous rearing of the maize stem borer Busseola fusca on an artificial diet. Entomol. Exp. Appl. 1994, 73, 139–144. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Tilman, D.; Knops, J.M. Herbivore Effects on Plant and Nitrogen Dynamics in Oak Savanna. Ecology 1998, 79, 165–177. [Google Scholar] [CrossRef]
- De Lange, E.S.; Laplanche, D.; Guo, H.; Xu, W.; Vlimant, M.; Erb, M.; Ton, J.; Turlings, T.C.J. Spodoptera frugiperda Caterpillars Suppress Herbivore-Induced Volatile Emissions in Maize. J. Chem. Ecol. 2020, 46, 344–360. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Ofomata, V.C.; Overholt, W.A.; Lux, S.A.; Van Huis, A.; Egwuatu, R.I. Comparative studies on the fecundity, egg survival, larval feeding, and development of Chilo partellus and Chilo orichalcociliellus (Lepidoptera: Crambidae) on five grasses. Ann. Entomol. Soc. Am. 2000, 93, 492–499. [Google Scholar] [CrossRef]
- Zhao, X.; Tong, C.; Pang, X.; Wang, Z.; Guo, Y.; Du, F.; Wu, R. Functional mapping of ontogeny in flowering plants. Brief. Bioinform. 2011, 13, 317–328. [Google Scholar] [CrossRef] [Green Version]
- FAO; CABI. Community-Based Fall Armyworm Monitoring, Early Warning and Management: Training of Trainers Manual; FAO: Rome, Italy, 2019. [Google Scholar]
- Luginbill, P. The fall army worm. Nature 1928, 121, 770–771. [Google Scholar]
- R Core Team. A Language and Environment for Statistical Computing. 2020. Available online: http://www.R-project.org (accessed on 20 May 2021).
- Acevedo, F.E.; Peiffer, M.; Ray, S.; Meagher, R.; Luthe, D.S.; Felton, G.W. Intraspecific differences in plant defense induction by fall armyworm strains. New Phytol. 2018, 218, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, F.E.; Peiffer, M.; Tan, C.-W.; Stanley, B.A.; Stanley, A.; Wang, J.; Jones, A.G.; Hoover, K.; Rosa, C.; Luthe, D.; et al. Fall armyworm-associated gut bacteria modulate plant defense responses. Mol. Plant-Microbe Interact. 2017, 30, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Mutyambai, D.M.; Bruce, T.; Berg, J.V.D.; Midega, C.A.O.; Pickett, J.A.; Khan, Z.R. An indirect defence trait mediated through egg-induced maize volatiles from neighbouring plants. PLoS ONE 2016, 11, e0158744. [Google Scholar] [CrossRef]
- Kant, M.R.; Jonckheere, W.; Knegt, B.; Lemos, F.; Liu, J.; Schimmel, B.; Villarroel, C.A.; Ataíde, L.; Dermauw, W.; Glas, J.J.; et al. Mechanisms and ecological consequences of plant defence induction and suppression in herbivore communities. Ann. Bot. 2015, 115, 1015–1051. [Google Scholar] [CrossRef]
- Bentivenha, J.; Baldin, E.L.L.; Hunt, T.E.; Paula-Moraes, S.V.; Blankenship, E.E. Intraguild Competition of Three Noctuid Maize Pests. Environ. Entomol. 2016, 45, 999–1008. [Google Scholar] [CrossRef]
- Sokame, B.M.; Tonnang, H.E.Z.; Subramanian, S.; Bruce, A.Y.; Dubois, T.; Ekesi, S.; Calatayud, P.-A. A System Dynamics Model for Pests and Natural Enemies Interactions. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Ryuda, M.; Nakayama, H.; Hayakawa, Y. A novel gene associated with intraspecific predation in Spodoptera litura larvae. Appl. Entomol. Zool. 2008, 43, 563–568. [Google Scholar] [CrossRef]
- Spark, A.N. A Review of the Biology of the Fall Armyworm. Fla. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Ren, Q.; Haseeb, M.; Fan, J.; Wu, P.; Tian, T.; Zhang, R. Functional response and intraspecific competition in the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2020, 11, 806. [Google Scholar] [CrossRef]
- Kfir, R. Competitive displacement of Busseola fusca (Lepidoptera: Noctuidae) by Chilo partellus (Lepidoptera: Pyralidae). Ann. Entomol. Soc. Am. 1997, 90, 619–624. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Combinations | Survival (%) | p-Value | Weight (mg) | p-Value | Length (cm) | p-Value |
|---|---|---|---|---|---|---|
| S.f parameters co-inhabiting with: | ||||||
| B.f | 0.47 | 0.75 | 0.60 | |||
| S.f | ||||||
| S.f parameters co-inhabiting with: | ||||||
| C.p | 0.66 | 0.91 | 0.66 | |||
| S.f | ||||||
| B.f parameters co-inhabiting with: | ||||||
| S.f | <0.001 | 0.01 | <0.001 | |||
| B.f | ||||||
| B.f parameters co-inhabiting with: | ||||||
| C.p | 0.12 | 0.17 | 0.19 | |||
| B.f | ||||||
| C.p parameters co-inhabiting with: | ||||||
| S.f | <0.001 | <0.001 | <0.001 | |||
| C.p | ||||||
| C.p parameters co-inhabiting with: | ||||||
| B.f | 0.12 | 0.27 | 0.14 | |||
| C.p |
| Infestation | % Predation of S.f on B.f | % Predation of S.f on C.p on | |||
|---|---|---|---|---|---|
| S.f Instar | B.f and C.p Instars | Maize Plant | Petri Dish | Maize Plant | Petri Dish |
| 6th | 62.5 a | 87.50 12.50 a | 6.37 a | .00 a | |
| 5th | 2.50 a | 100.00 a | 87.52.50 a | .00 a | |
| 6th | 4th | 87.50 a | 100.00 a | 87.52.50 a | .00 a |
| 3rd | 6.37 a | .00 a | 87.52.50 a | .00 a | |
| 2nd | 6.37 a | .00 a | 6.37 a | .00 a | |
| 1st | 6.37 a | a | 6.37 a | a | |
| 6th | a | 87.52.50 a | 756.37 a | 87.52.50 a | |
| 5th | 756.37 a | 100.00 a | 87.52.50 a | 100.00 a | |
| 5th | 4th | 756.37 a | 100.00 a | 87.52.50 a | 100.00 a |
| 3rd | 87.52.50 a | 100.00 a | 87.5 12.50 a | 100.00 a | |
| 2nd | 756.37 a | 100.00 a | 756.37 a | 100.00 a | |
| 1st | 62.58.30 a | 100.00 b | 62.58.30 a | 100.00 b | |
| 6th | 62.58.30 a | 756.37 a | 62.58.30 a | 87.52.50 a | |
| 5th | 62.58.30 a | 87.52.50 a | 756.37 a | 87.52.50 a | |
| 4th | 4th | 756.37 a | 87.52.50 a | 756.37 a | 100.00 a |
| 3rd | 756.37 a | 100.00 a | 756.37 a | 100.00 a | |
| 2nd | 62.58.30 a | 100.00 b | 62.58.30 a | 100.00 b | |
| 1st | 62.58.30 a | 100.00 b | 62.58.30 a | 100.00 b | |
| 6th | 0.00 a | 37.5 b | 12.50 a | 50 a | |
| 5th | 12.50 a | 50 | 25 | 62.5 a | |
| 3rd | 4th | 50 a | 50 a | 50 a | 62.5 a |
| 3rd | 756.37 a | 87.52.50 a | 756.37 a | 87.52.50 a | |
| 2nd | 756.37 a | 87.52.50 a | 756.37 a | 87.52.50 a | |
| 1st | 62.58.30 a | 87.52.50 a | 50 | 100.00 b | |
| 6th | 0.00 a | 0.00 a | 0.00 | 0.00 a | |
| 5th | 0.00 a | 0.00 a | 0.00 a | 0.00 a | |
| 2nd | 4th | 0.00 a | 12.50 0 a | 0.00 a | 12.50 a |
| 3rd | 0.00 a | 37.5 | 12.50 | 50 a | |
| 2nd | 12.50 a | 62.58.30 b | 12.50 a | 756.37 b | |
| 1st | 25.00 16.37 a | 756.37 b | 25.00 16.37 a | 87.52.50 b | |
| Localities | Population Density | Predation of S.f on: | Cannibalism of S.f in Presence of: | Psb | Psc | Pbp | Pcp | |||
|---|---|---|---|---|---|---|---|---|---|---|
| B.f | C.p | S.f | C.p | B.f | ||||||
| Maize | 4 | 0.33 | 0.33 | <0.001 | <0.001 | |||||
| Maize | 8 | 0.04 | 0.01 | <0.001 | <0.001 | |||||
| Maize | 16 | <0.001 | <0.001 | <0.001 | <0.001 | |||||
| Petri dish | 4 | 4.09 | 0.14 | 0.14 | <0.001 | <0.001 | ||||
| Petri dish | 8 | 4.09 | 0.007 | 0.002 | <0.001 | <0.001 | ||||
| Petri dish | 16 | <0.001 | <0.001 | <0.001 | <0.001 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mutua, J.M.; Mutyambai, D.M.; Asudi, G.O.; Khamis, F.; Niassy, S.; Jalloh, A.A.; Salifu, D.; Magara, H.J.O.; Calatayud, P.-A.; Subramanian, S. Competitive Plant-Mediated and Intraguild Predation Interactions of the Invasive Spodoptera frugiperda and Resident Stemborers Busseola fusca and Chilo partellus in Maize Cropping Systems in Kenya. Insects 2022, 13, 790. https://doi.org/10.3390/insects13090790
Mutua JM, Mutyambai DM, Asudi GO, Khamis F, Niassy S, Jalloh AA, Salifu D, Magara HJO, Calatayud P-A, Subramanian S. Competitive Plant-Mediated and Intraguild Predation Interactions of the Invasive Spodoptera frugiperda and Resident Stemborers Busseola fusca and Chilo partellus in Maize Cropping Systems in Kenya. Insects. 2022; 13(9):790. https://doi.org/10.3390/insects13090790
Chicago/Turabian StyleMutua, Johnstone Mutiso, Daniel Munyao Mutyambai, George Ochieng’ Asudi, Fathiya Khamis, Saliou Niassy, Abdul A. Jalloh, Daisy Salifu, Henlay J. O. Magara, Paul-André Calatayud, and Sevgan Subramanian. 2022. "Competitive Plant-Mediated and Intraguild Predation Interactions of the Invasive Spodoptera frugiperda and Resident Stemborers Busseola fusca and Chilo partellus in Maize Cropping Systems in Kenya" Insects 13, no. 9: 790. https://doi.org/10.3390/insects13090790
APA StyleMutua, J. M., Mutyambai, D. M., Asudi, G. O., Khamis, F., Niassy, S., Jalloh, A. A., Salifu, D., Magara, H. J. O., Calatayud, P. -A., & Subramanian, S. (2022). Competitive Plant-Mediated and Intraguild Predation Interactions of the Invasive Spodoptera frugiperda and Resident Stemborers Busseola fusca and Chilo partellus in Maize Cropping Systems in Kenya. Insects, 13(9), 790. https://doi.org/10.3390/insects13090790