


Within-Plant and Within-Field Distribution Patterns of Asian Bean Thrips and Melon Thrips in Snap Bean





Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Crop Management
2.2. Sampling for Within-Plant Distribution of Thrips
2.3. Sampling Thrips for Within-Field Distribution
2.4. Statistical Analysis
2.4.1. Within-Plant Distribution
2.4.2. Within-Field Distribution
- s2 = sample variance,
- = sample mean,
- x1 = number of thrips found in the sample units,
- n = number of sample units.
3. Results
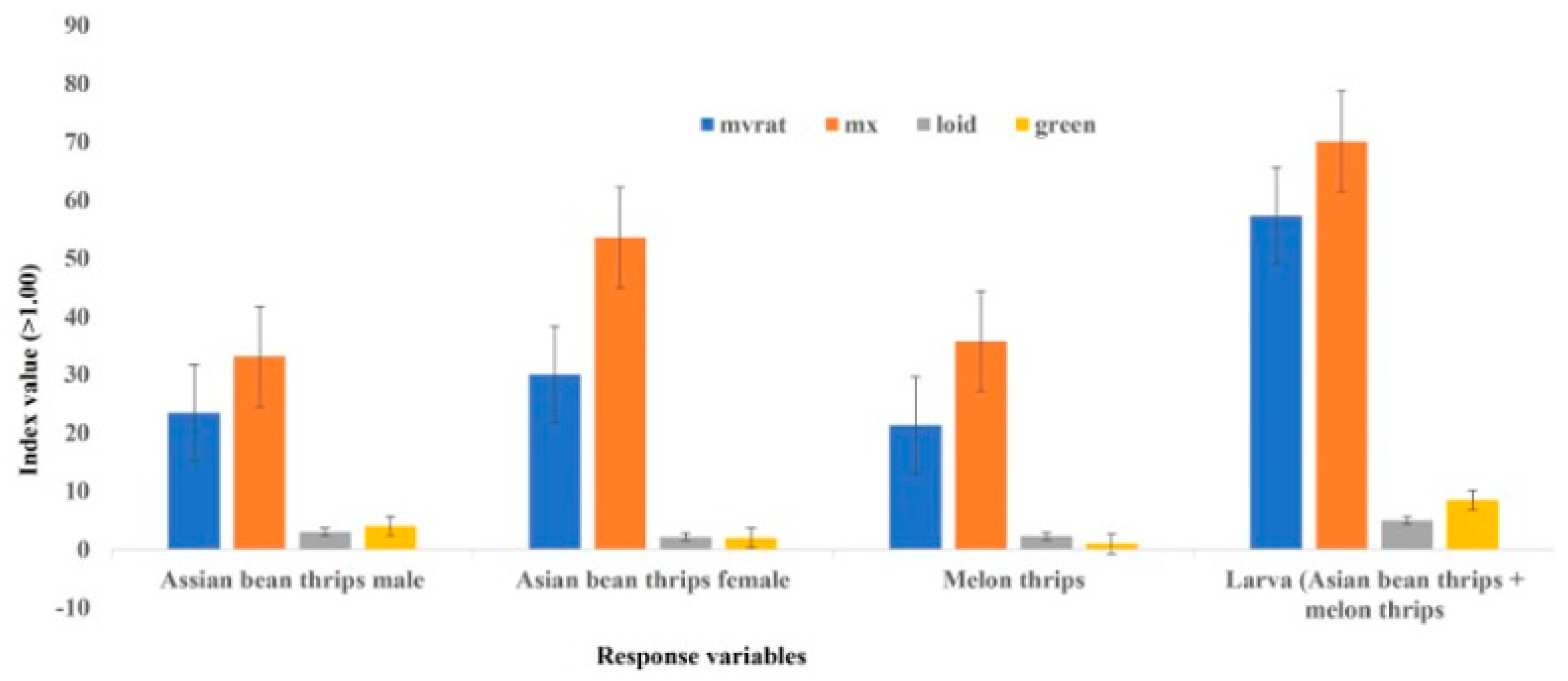
3.1. Within-Plant Distribution
3.2. Within-Field Distribution Based on Flower and Leaf Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, S.; Seal, D.; Ozores-Hampton, M.; Lambert, M.; Li, Y.C.; Klassen, W.; Olczyk, T. Bush Snapbean Production in Miami-Dade County, Florida. 2014. Available online: https://edis.ifas.ufl.edu/publication/TR005 (accessed on 2 February 2020).
- Mound, L.A. Biological diversity. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: Wallingford, UK, 1997; pp. 197–225. [Google Scholar]
- Soto-Adames, F.N. Megalurothrips usitatus (Bagnall), Asian bean thrips, Oriental bean flower thrips or bean flower thrips. Pest Alert FDACS-P-02137. In Bureau of Entomology, Nematology and Plant Pathology; Florida Department of Agriculture and Consumer Services Division of Plant Industry: Gainesville, FL, USA, 2020. [Google Scholar]
- Mound, L.A. Thysanoptera: Diversity and interactions. Annu. Rev. Entomol. 2005, 50, 247–269. [Google Scholar] [PubMed]
- Cho, K.; Kang, S.H.; Lee, G.S. Spatial distribution and sampling plans for Thrips palmi (Thysanoptera: Thripidae) infesting fall potato in Korea. J. Econ. Entomol. 2000, 93, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, S.; Funderburk, J.; Stavisky, J.; Olson, S. Population abundance and movement of Frankliniella species and Orius insidiosus in field pepper. Agric. For. Entomol. 2001, 3, 129–137. [Google Scholar]
- Reitz, S.R.; Yearby, E.L.; Funderburk, J.E.; Stavisky, J.; Momol, M.T.; Olson, S.M. Integrated management tactics for Frankliniella thrips (Thysanoptera: Thripidae) in field-grown pepper. J. Econ. Entomol. 2003, 96, 1201–1214. [Google Scholar] [CrossRef] [PubMed]
- Seal, D.R. Management, and biology of Thrips palmi Karny (Thysanoptera: Thripidae). In New Developments in Entomology; Bondari, K., Ed.; T.C. 36/248 (1), Tri-Vandrum-695008; Research Signpost: Trivandum, India, 1997; pp. 161–181. [Google Scholar]
- Kao, S.S. The spatial distribution of insects. In Phytopathologist and Entomologist; NTU: Taipei, Taiwan, 1984; Volume 11, pp. 18–32. [Google Scholar]
- Southwood, T.R.E. Ecological Methods; Springer: Dordrecht, The Netherlands, 1978. [Google Scholar]
- Seal, D.R.; Stansly, P.A. Seasonal abundance and within plant distribution of melon thrips (Thysanoptera: Thripidae) on beans in Southern Florida. In Proceedings of the Florida State Horticultural Society, Florida, FL, USA, 23–25 July 2000; Volume 113, pp. 201–205. [Google Scholar]
- Seal, D.R.; Ciomperlik, M.A.; Richards, M.L.; Klassen, W. Distribution of chilli thrips, Scirtothrips dorsalis (Thysanoptera: Thripidae), in pepper fields and pepper plants on St. Vincent. Fla. Entomol. 2006, 89, 311–320. [Google Scholar] [CrossRef]
- Weisz, R.; Fleischer, S.; Smilowitz, Z. Site-specific integrated pest management for high value crops: Sample units for map generation using the Colorado potato beetle (Coleoptera: Chrysomelidae) as a model system. J. Econ. Entomol. 1995, 88, 1069–1080. [Google Scholar] [CrossRef]
- Jacobson, R.J. Integrated pest management (IPM) in glasshouses. In Thrips as Crops Pests; Lewis, T., Ed.; CAB International: Wallingford, UK, 1997; pp. 639–666. [Google Scholar]
- Nobel, C.V.; Drew, R.W.; Slabaugh, V. Soil Survey of Dade County Area, Florida; U.S. Department of Agriculture Natural Resources Conservation Service: Washington, DC, USA, 1996.
- Dittmar, P.J.; Freeman, J.H.; Vallad, G.E. (Eds.) Commercial Vegetable Production in Florida. In Vegetable Production Handbook of Florida; UF, IFAS Extension, University of Florida: Gainesville, FL, USA, 2015; p. 248. [Google Scholar]
- Seal, D.R.; Baranowski, R.M. Effectiveness of different insecticides for the control of melon thrips, Thrips palmi Karny (Thysanoptera: Thripidae) affecting vegetables in south Florida. In Proceedings of the Florida State Horticultural Society, Gainesville, FL, USA, 3–5 November 1992. [Google Scholar]
- Adhikari, R. Distribution Pattern, Biology, and Management of Asian Bean Thrips (Megalurothrips usitatus Bangall) and Melon Thrips (Thrips palmi Karny) with an Emphasis on Beans in South Florida. Master’s Thesis, Entomology and Nematology Department, University of Florida, Gainesville, FL, USA, 2020. [Google Scholar]
- Mound, L.A. Australian Thysanoptera-biological diversity and a diversity of studies. Aust. J. Entomol. 2004, 43, 248–257. [Google Scholar] [CrossRef]
- Nakahara, S. Annotated list of the Frankliniella species of the world (Thysanoptera: Thripidae). Contributions on Entomology International. 1997, Volume 2, pp. 355–389. Available online: https://www.cabdirect.org/cabdirect/abstract/19981105670 (accessed on 20 November 2022).
- SAS Institute. SAS/STAT 9.3 User’s Guide; SAS Institute: Cary, NC, USA, 2013. [Google Scholar]
- Khan, R.A.; Seal, D.R.; Zhang, S.; Liburd, O.E.; Srinivasan, R.; Evans, E. Distribution pattern of thrips (Thysanoptera: Thripidae) and tomato chlorotic spot virus in South Florida Tomato Fields. Environ. Entomol. 2020, 49, 73–87. [Google Scholar] [CrossRef]
- Rozsa, L.; Reiczigel, J.; Majoros, G. Quantifying parasites in samples of hosts. J. Parasitol. 2000, 86, 228–232. [Google Scholar] [CrossRef]
- Rabinovich, J.E. Introduction a la Ecologia de Poblaciónes Animals; Compania Editorial Continental: Mexico City, Mexico, 1980. [Google Scholar]
- Green, R.H. Measurement of non-randomness in spatial distributions. Popul. Ecol. 1966, 8, 1–7. [Google Scholar]
- Taylor, L.R. Aggregation, variance, and the mean. Nature 1961, 189, 732–735. [Google Scholar] [CrossRef]
- Iwao, S. A new regression method for analyzing the aggregation pattern of animal populations. Popul. Ecol. 1968, 10, 1–20. [Google Scholar] [CrossRef]
- Cox, D.R.; Lewis, P.A.W. The Statistical Analysis of Series of Events; Methuen: London, UK, 1966; p. 72. [Google Scholar]
- Lloyd, M. Mean crowding. J. Anim. Ecol. 1967, 36, 1–30. [Google Scholar]
- Wilson, L.T.; Room, P.M. Relative efficiency and reliability of three methods for sampling arthropods in Australian cotton fields. Aust. J. Entomol. 1982, 21, 175–181. [Google Scholar] [CrossRef]
- Kawai, A. Life cycle and population dynamics of Thrips palmi Karny. JARQ 1990, 23, 282–288. [Google Scholar]
- Kakkar, G.; Seal, D.R.; Stansly, P.A.; Liburd, O.E.; Kumar, V. Abundance of Frankliniella schultzei (Thysanoptera: Thripidae) in flowers on major vegetable crops of south Florida. Fla. Entomol. 2012, 95, 468–475. [Google Scholar]
- Milne, M.; Walter, G.H.; Milne, J.R. Mating behavior and species status of host-associated populations of the polyphagous thrips, Frankliniella schultzei. J. Insect Behav. 2007, 20, 331–346. [Google Scholar] [CrossRef]
- Kakkar, G.; Seal, D.R.; Kumar, V. Assessing abundance and distribution of an invasive thrips Frankliniella schultzei (Thysanoptera: Thripidae) in south Florida. Bull. Entomol. Res. 2012, 102, 249–259. [Google Scholar]
- Sedaratian, A.A.; Fathipour, Y.; Talebi, A.A.; Farahani, S. Population density and spatial distribution pattern of Thrips tabaci (Thysanoptera: Thripidae) on different soybean varieties. J. Agric. Sci. Technol. 2010, 12, 275–288. [Google Scholar]
- Wang, K.; Shipp, J.L. Simulation model for population dynamics of Frankliniella occidentalis (Thysanoptera: Thripidae) on greenhouse cucumber. Environ. Entomol. 2001, 30, 1073–1081. [Google Scholar] [CrossRef]
- Parajulee, M.N.; Shrestha, R.B.; Leser, J.F. Sampling methods, dispersion patterns, and fixed precision sequential sampling plans for western flower thrips (Thysanoptera: Thripidae) and cotton fleahopper (Hemiptera: Miridae) in cotton. J. Econ. Entomol. 2006, 99, 568–577. [Google Scholar] [CrossRef]
- García-Mari, F.; González-Zamora, J.E.; Ribes, A.; Benages, E.; Meseguer, A. Méthodos de muestro bionomial y sequencial del trips de las floras Frankliniella occidentalis (Pergande) (Thysanoptera, Thripidae) y de antocóridos (Heteroptera, Anthocoridae) en fresón. Bol. San. Veg. Plagas. 1994, 20, 703–723. [Google Scholar]
- Deligeorgidis, P.N.; Anthanassiou, C.G.; Kavallieratos, N.G. Seasonal abundance, spatial distribution, and sampling indices of thrips populations on cotton; a 4-year survey from central Greece. J. Appl. Entomol. 2002, 126, 343–348. [Google Scholar] [CrossRef]
- Larral, P.; Ripa, R.; Funderburk, J.; Lopez, E. Population abundance, phenology, spatial distribution and a binominal sampling plan for Heliothrips haemorrhoidalis (Thysanoptera: Thripidae) in avocado. Fla. Entomol. 2018, 101, 166–171. [Google Scholar] [CrossRef]
- Aliakbarpour, H.; Salmah, M.R.C. Seasonal abundance and spatial distribution of larval and adult thrips (Thysanoptera) on weed host plants in mango orchards in Penang, Malaysia. Appl. Entomol. Zool. 2011, 46, 185–194. [Google Scholar] [CrossRef]
- Navarro-Campos, C.; Aguilar, A.; Garcia-Mari, F. Aggregation pattern, sampling plan, and intervention threshold for Pezothrips kellyanus in citrus groves. Entomol. Exp. Appl. 2012, 142, 130–139. [Google Scholar]
- Salguero Navas, V.E.; Funderburk, J.E.; Olson, S.M.; Beshear, R.J. Damage to tomato fruit by the western flower thrips (Thysanoptera: Thripidae). Entomol. Sci. 1991, 26, 436–442. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Year | Field No. | Coordinates | Area (m2) | East | West | North | South |
|---|---|---|---|---|---|---|---|
| 2020 | 1 | 25°30′17.5″ N 80°32′01.3″ W | 20,234 | Squash, bean and fallow | Bean, housing apartments | Nursery, tomato and bean | Fallow, corn squash |
| 2020 | 2 | 25°29′39.1” N 80°30′08.5″ W | 28,328 | Tomato | Nursery | Bean | Bean |
| 2021 | 3 | 25°29′22.9″ N 80°30′13.6″ W | 32,375 | Squash, bean | Bean, fallow | Tomato, housing | Bean, nursery, and housing |
| 2021 | 4 | 25°30′17″ N 80°29′56″ W | 20,234 | Housing, squash and bean | Nursery, bean and squash | Bean, nursery | Tomato, squash |
| 2022 | 5 | 25°30′23″ N 80°28′50″ W | 20,234 | Squash, okra | Bean, weed, and housing | Squash | Squash |
| 2022 | 6 | 25°30′30″ N 80°28′30″ W | 24,281 | Palm grove | Bean | bean | Fallow |
| Field | Plant Parts | ABM | ABF | MT | LAR |
|---|---|---|---|---|---|
| Flowers | 41.70 ± 2.39 a | 74.85 ± 3.36 a | 40.65 ± 2.50 a | 72.48 ± 3.48 a | |
| Field 1 | Leaves | 2.62 ± 0.25 b | 2.00 ± 0.26 b | 4.60 ± 0.57 b | 40.08 ± 3.29 b |
| Pods | 0.08 ± 0.05 c | 0.65 ± 0.35 b | 0.37 ± 0.19 c | 0.29 ± 0.09 c | |
| Flowers | 10.29 ± 0.70 a | 35.29 ± 1.70 a | 33.60 ±1.88 a | 1.25 ± 0.22 a | |
| Field 2 | Leaves | 0.58 ± 0.14 b | 0.20 ± 0.07 b | 8.97 ± 0.82 b | 0.22 ± 0.08 b |
| Pods | 0.04 ± 0.04 b | 0.08 ± 0.05 b | 0.12 ± 0.05 c | 0 b | |
| Flowers | 5.25 ±0.58 a | 15.04 ± 1.13 a | 4.94 ± 0.48 a | 5.16 ± 0.62 a | |
| Field 3 | Leaves | 0.70 ± 0.12 b | 1.10 ± 0.19 b | 6.18 ± 0.75 a | 10.60 ±1.16 a |
| Pods | 0.10 ± 0.05 b | 0.39 ± 0.09 b | 0.39 ± 0.19 b | 1.00 ±0.42 b |
| 11.14 m2 | 44.59 m2 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Taylor’s Power Law | Iwao’s Patchiness | Taylor’s Power Law | Iwao’s Patchiness | |||||||||
| Fld1 | 0.27 | −2.75 | 2.88 ag 1 | 0.91 | −2.37 | 1.12 ag | 0.72 | −2.05 | 2.72 ag | 0.97 | −3.47 | 1.20 ag |
| Fld2 | 0.11 | −0.19 | 1.24 ag | 0.76 | 1.17 | 0.98 rn | 0.28 | 0.52 | 0.78 re | 0.88 | 1.77 | 0.94 re |
| Fld3 | 0.05 | −0.19 | 0.87 re | 0.78 | −0.65 | 1.13 ag | 0.83 | −2.32 | 4.22 ag | 0.93 | −3.13 | 1.63 ag |
| Fld4 | 0.02 | −0.03 | 0.21 re | 0.14 | 0.86 | 0.48 re | 0.04 | 0.05 | 0.30 re | 0.54 | 0.37 | 0.68 re |
| Fld5 1 | 0.02 | −0.03 | 0.21 re | 0.14 | 0.86 | 0.48 re | 0.04 | 0.05 | 0.29 re | 0.53 | 0.37 | 0.68 re |
| Fld6 | 0.03 | 0.04 | −0.26 re | 0.74 | 0.33 | 0.73 re | 0.52 | −0.74 | 2.24 ag | 0.87 | −1.69 | 1.50 ag |
| All fields | 0.78 | −1.06 | 3.33 ag | 0.57 | −41.39 | 3.94 ag | 0.9 | −1.05 | 3.33 ag | 0.9 | −34.40 | 6.33 ag |
| 11.14 m2 | 44.59 m2 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Taylor’s Power Law | Iwao’s Patchiness | Taylor’s Power Law | Iwao’s Patchiness | |||||||||
| r2 | a | b | r2 | a | β | r2 | a | b | r2 | a | β | |
| Fld1 | 0.01 | 1.46 | 0.27 re 1 | 0.98 | 2.19 | 0.99 rn | 0.82 | 12.62 | 8.07 ag | 0.98 | −44.80 | 1.67 ag |
| Fld2 | 0.17 | −2.96 | 2.90 ag | 0.93 | −4.03 | 1.15 ag | 0.61 | −5.55 | 4.84 ag | 0.97 | −11.53 | 1.39 ag |
| Fld3 | 0.08 | −0.26 | 1.18 ag | 0.81 | −1.04 | 1.15 ag | 0.19 | −0.38 | 1.54 ag | 0.84 | 0.7 | 1.04 ag |
| Fld4 | 0.03 | −0.49 | 0.70 re | 0.94 | −0.49 | 0.95 rn | 0.09 | −0.52 | 0.93 re | 0.98 | −0.75 | 1.01 rn |
| Fld5 | 0.003 | 0.1 | 0.26 re | 0.93 | −0.19 | 0.96 rn | 0.2 | −0.96 | 1.73 ag | 0.97 | −0.37 | 0.99 rn |
| Fld6 | 0.006 | 0.36 | −0.05 re | 0.92 | −0.01 | 0.95 rn | 0.2 | −0.96 | 1.74 ag | 0.97 | −0.37 | 1.00 rn |
| All fields | 0.63 | −1.58 | 3.18 ag | 0.87 | −19.47 | 3.69 ag | 0.62 | −1.69 | 3.26 ag | 0.66 | −55.48 | 4.41 ag |
| 11.14 m2 | 44.59 m2 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Taylor’s Power Law | Iwao’s Patchiness | Taylor’s Power Law | Iwao’s Patchiness | |||||||||
| Fields | r2 | a | b | r2 | a | β | r2 | a | b | r2 | a | β |
| Fld1 | 0.02 | 0.77 | 0.71 re 1 | 0.83 | 6.99 | 0.92 re | 0.68 | −1.47 | 2.43 ag | 0.94 | −6.09 | 1.29 ag |
| Fld2 | 0.14 | −2.52 | 2.49 ag | 0.95 | −1.46 | 1.07 ag | 0.13 | −1.19 | 2.15 ag | 0.85 | −2.22 | 1.17 ag |
| Fld3 | 0.13 | −0.33 | 0.88 re | 0.89 | −0.36 | 1.01 rn | 0.74 | −1.69 | 3.38 ag | 0.96 | −2.51 | 1.55 ag |
| Fld4 | 0.02 | −0.09 | 0.17 re | 0.59 | 0.29 | 0.68 re | 0.14 | −0.24 | 0.83 re | 0.81 | −0.61 | 1.10 ag |
| Fld5 | 0.008 | −0.29 | 0.37 re | 0.84 | −0.86 | 1.04 rn | 0.11 | −0.35 | 0.86 re | 0.95 | −0.63 | 1.00 rn |
| Fld6 | 0.04 | −0.34 | 0.74 re | 0.83 | −0.06 | 0.95 rn | 0.58 | −0.82 | 1.64 ag | 0.99 | −0.90 | 1.07 ag |
| All fields | 0.94 | −0.84 | 2.81 ag | 0.91 | −16.65 | 3.37 ag | 0.96 | −1.39 | 3.28 ag | 0.94 | −27.39 | 4.07 ag |
| 11.14 m2 | 44.59 m2 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Taylor’s Power Law | Iwao’s Patchiness | Taylor’s Power Law | Iwao’s Patchiness | |||||||||
| Fields | r2 | a | b | r2 | a | β | r2 | a | b | r2 | a | β |
| Fld1 | 0.03 | 2.86 | −0.4 re 1 | 0.67 | 12.35 | 0.92 re | 0.04 | 4.59 | 0.99 rn | 0.65 | 27.03 | 0.73 reg |
| Fld2 | 0.72 | 0.06 | 1.15 ag | 0.77 | −0.50 | 1.38 ag | 0.93 | −0.16 | 2.98 ag | 0.96 | −1.76 | 2.62 ag |
| Fld3 | 0.12 | −0.47 | 1.33 ag | 0.68 | −0.73 | 1.89 ag | 0.79 | −0.26 | 1.36 ag | 0.97 | −0.28 | 1.05 ag |
| Fld4 | 0.01 | −0.16 | 0.14 re | 0.36 | 0.17 | 0.78 re | 0.13 | −0.03 | 0.71 re | 0.61 | 0.01 | 0.93 re |
| Fld5 | 0.04 | −0.12 | 0.37 re | 0.6 | 0.23 | 0.77 re | 0.09 | 0.001 | 0.42 re | 0.89 | −0.06 | 0.88 re |
| Fld6 | 0.01 | −0.16 | 0.14 re | 0.37 | 0.17 | 0.78 re | 0.13 | −0.03 | 0.71 re | 0.61 | 0.01 | 0.93 re |
| All fields | 0.92 | 0.17 | 2.37 ag | 0.84 | −10.89 | 5.77 ag | 0.79 | 0.88 | 1.76 ag | 0.57 | 10.18 | 4.21 reg |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adhikari, R.; Seal, D.R.; Schaffer, B.; Liburd, O.E.; Khan, R.A. Within-Plant and Within-Field Distribution Patterns of Asian Bean Thrips and Melon Thrips in Snap Bean. Insects 2023, 14, 175. https://doi.org/10.3390/insects14020175
Adhikari R, Seal DR, Schaffer B, Liburd OE, Khan RA. Within-Plant and Within-Field Distribution Patterns of Asian Bean Thrips and Melon Thrips in Snap Bean. Insects. 2023; 14(2):175. https://doi.org/10.3390/insects14020175
Chicago/Turabian StyleAdhikari, Rosan, Dakshina R. Seal, Bruce Schaffer, Oscar E. Liburd, and Rafia A. Khan. 2023. "Within-Plant and Within-Field Distribution Patterns of Asian Bean Thrips and Melon Thrips in Snap Bean" Insects 14, no. 2: 175. https://doi.org/10.3390/insects14020175
APA StyleAdhikari, R., Seal, D. R., Schaffer, B., Liburd, O. E., & Khan, R. A. (2023). Within-Plant and Within-Field Distribution Patterns of Asian Bean Thrips and Melon Thrips in Snap Bean. Insects, 14(2), 175. https://doi.org/10.3390/insects14020175