


Oviposition Preference of the American Hoverfly, Eupeodes americanus, between Banker Plants and Target Crops

 , and
, and 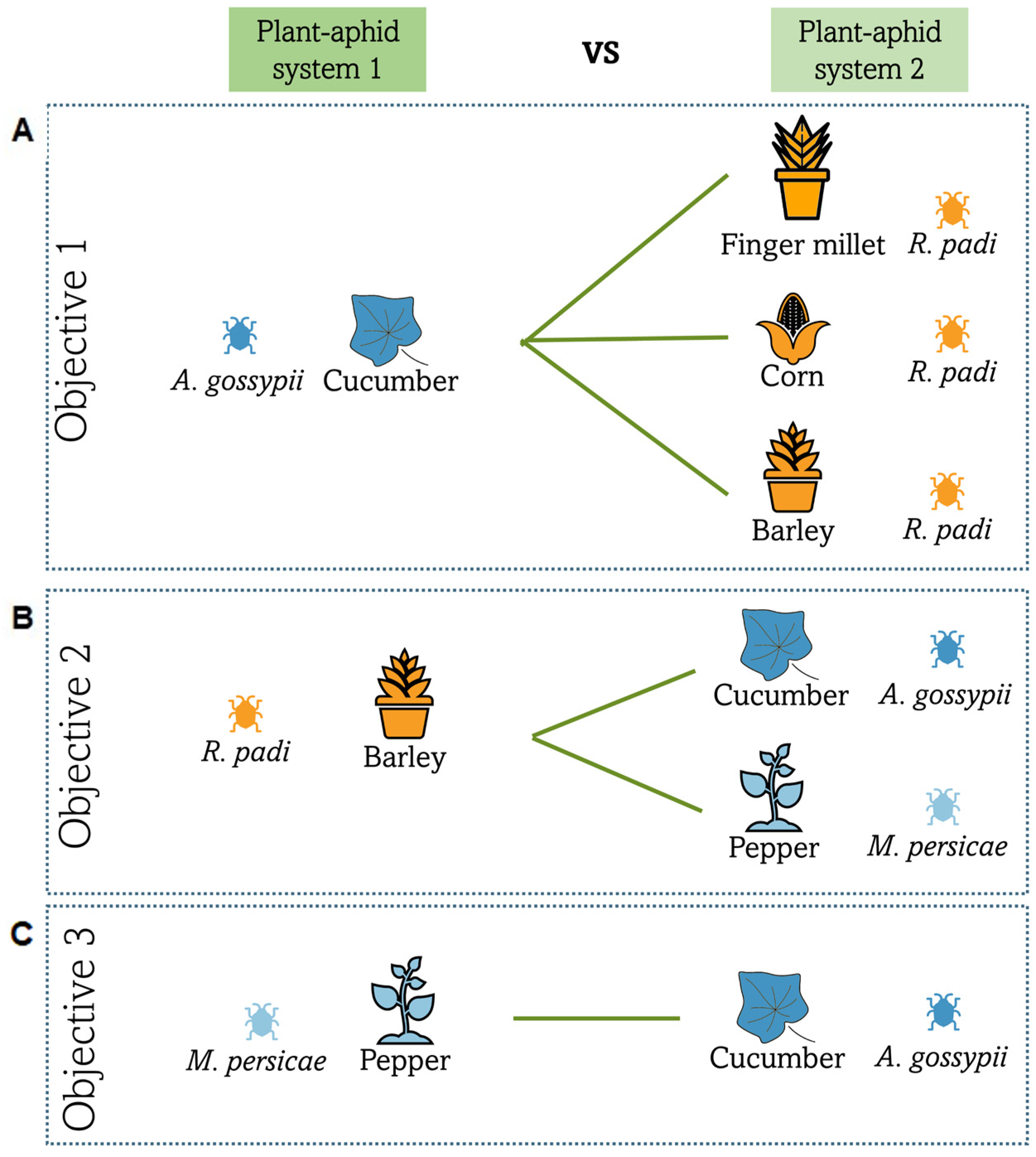{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
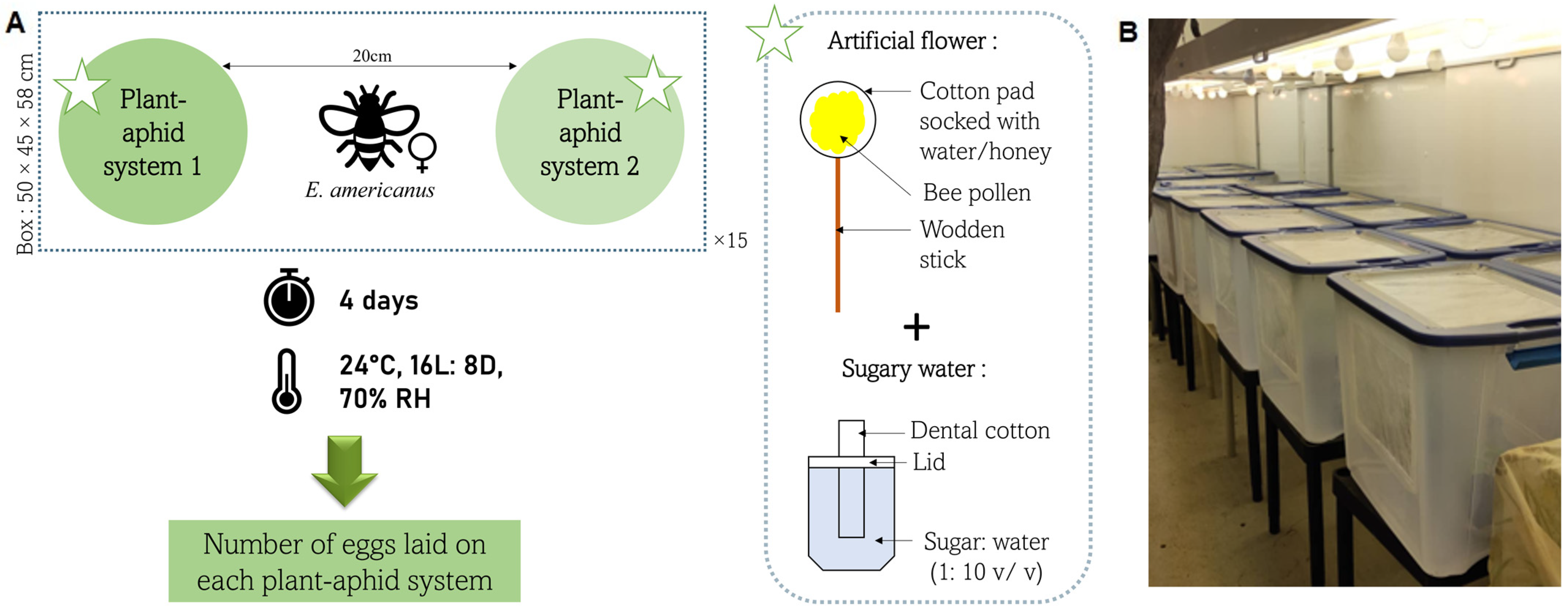
2. Materials and Methods
2.1. Plants
2.2. Insect Rearing
2.3. Objective 1: Suitability of Three Banker Plant Species to Control A. gossypii on Cucumber
2.4. Objective 2: Suitability of Barley Banker Plant Species to Control M. persicae on Pepper and Comparison with A. gossypii on Cucumber
2.5. Objective 3: Suitability of the American Hoverfly to Control Aphids in a Mixed Crop Greenhouse Context
2.6. Data Analysis
3. Results
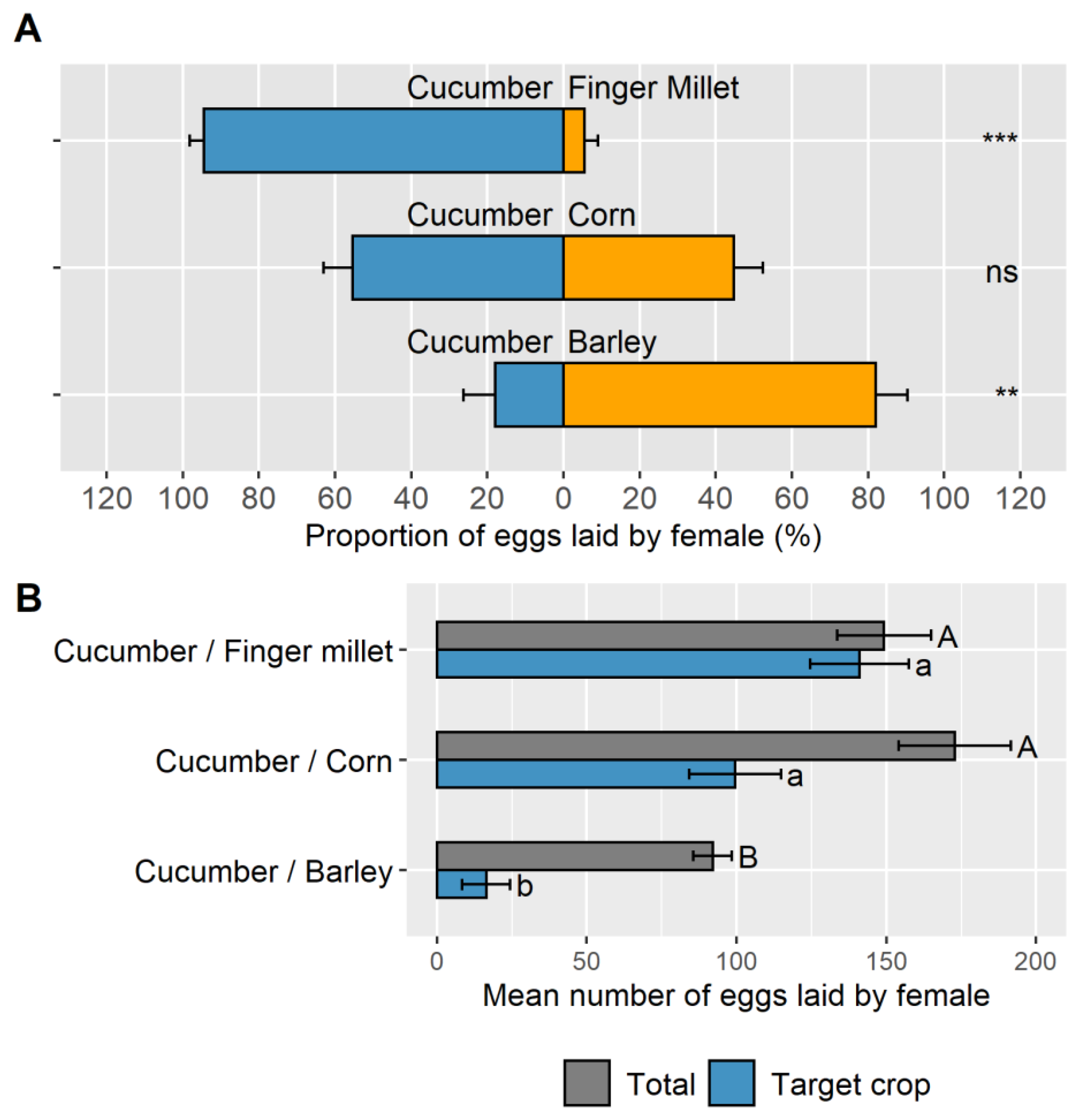
3.1. Objective 1: Suitability of Three Banker Plant Species to Control A. gossypii on Cucumber
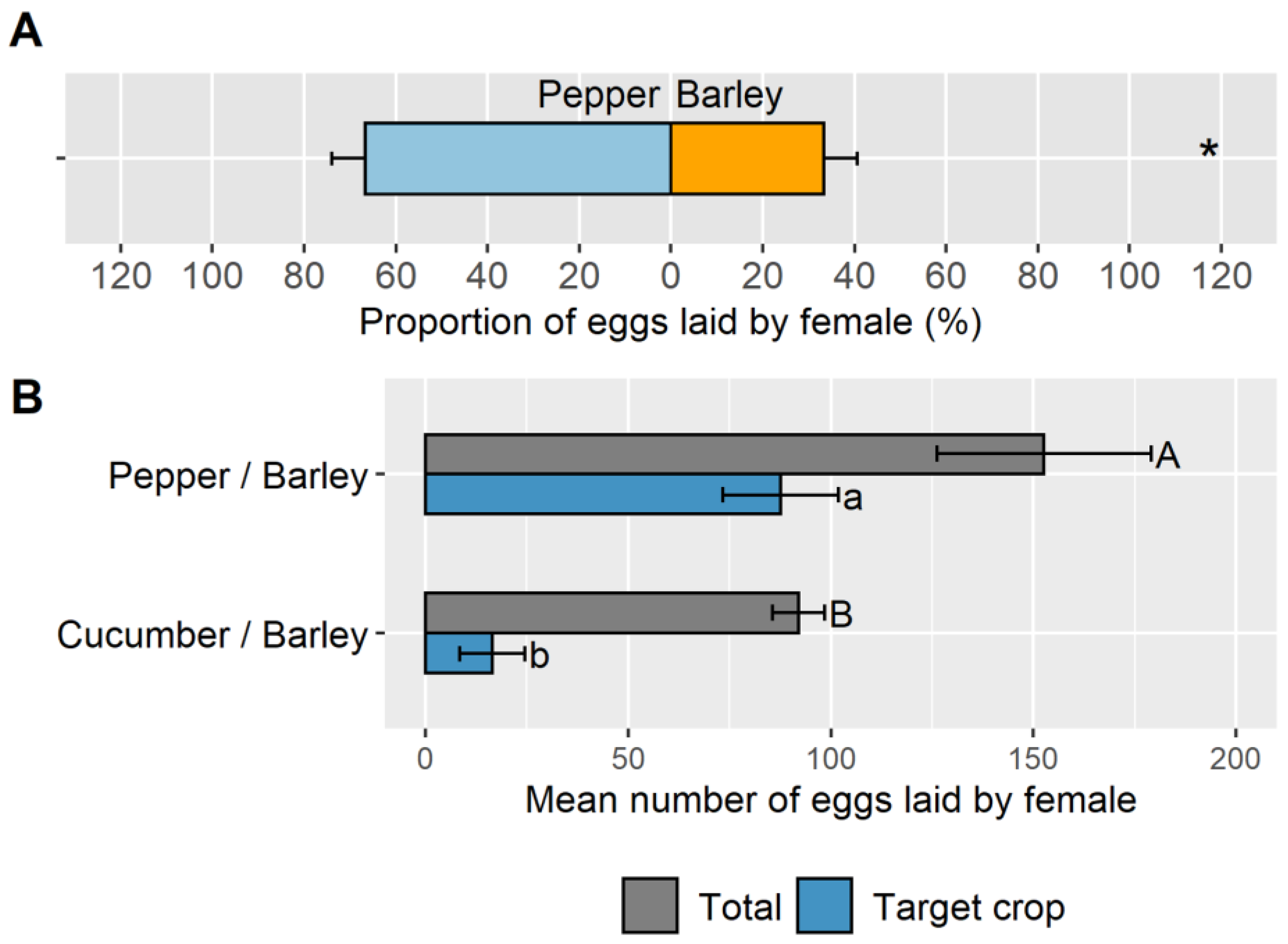
3.2. Objective 2: Suitability of Barley Banker Plant Species to Control M. persicae on Pepper and Comparison with A. gossypii on Cucumber
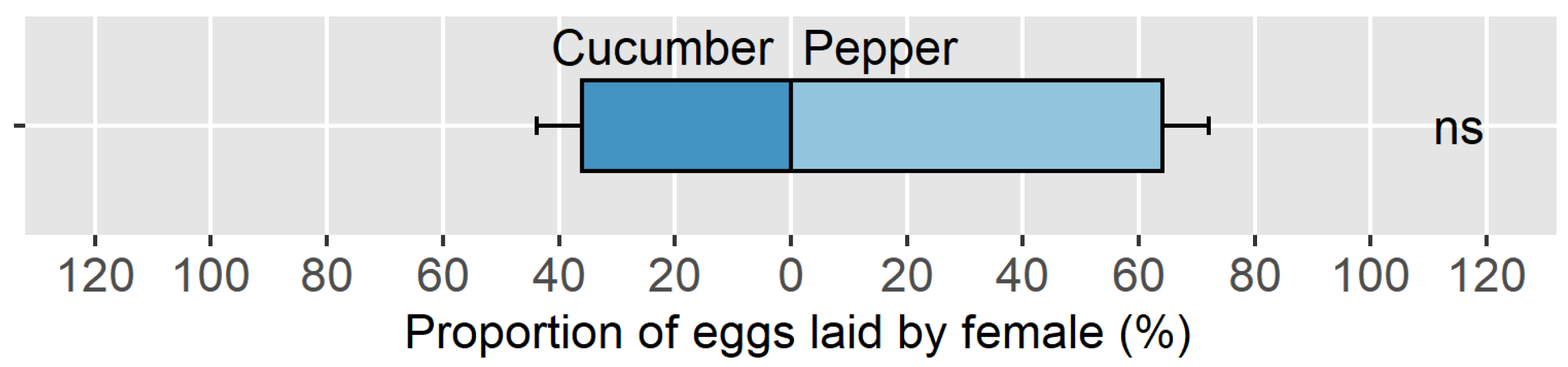
3.3. Objective 3: Suitability of the American Hoverfly to Control Aphids in a Mixed Crop Greenhouse Context
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rabasse, J.-M.; van Steenis, M.J. Biological Control of Aphids. In Integrated Pest and Disease Management in Greenhouse Crops; Albajes, R., Lodovica Gullino, M., van Lenteren, J.C., Elad, Y., Eds.; Developments in Plant Pathology; Springer: Dordrecht, The Netherlands, 1999; pp. 235–243. ISBN 978-0-306-47585-6. [Google Scholar]
- Knapp, M.; Palevsky, E.; Rapisarda, C. Insect and Mite Pests. In Integrated Pest and Disease Management in Greenhouse Crops; Gullino, M.L., Albajes, R., Nicot, P.C., Eds.; Plant Pathology in the 21st Century; Springer International Publishing: Cham, Switzerland, 2020; pp. 101–146. ISBN 978-3-030-22304-5. [Google Scholar]
- Capinera, J.L. Melon Aphid or Cotton Aphid, Aphis gossypii Glover (Insecta: Hemiptera: Aphididae): EENY-173/IN330, 11/2000. EDIS 2004, 2004, 1–5. [Google Scholar] [CrossRef]
- Mahmood, I.; Imadi, S.R.; Shazadi, K.; Gul, A.; Hakeem, K.R. Effects of Pesticides on Environment. In Plant, Soil and Microbes: Volume 1: Implications in Crop Science; Hakeem, K.R., Akhtar, M.S., Abdullah, S.N.A., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 253–269. ISBN 978-3-319-27455-3. [Google Scholar]
- Amorós-Jiménez, R.; Pineda, A.; Fereres, A.; Marcos-García, M. Prey Availability and Abiotic Requirements of Immature Stages of the Aphid Predator Sphaerophoria rueppellii. Biol. Control 2012, 63, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Amorós-Jiménez, R.; Pineda, A.; Fereres, A.; Marcos-García, M.Á. Feeding Preferences of the Aphidophagous Hoverfly Sphaerophoria rueppellii Affect the Performance of Its Offspring. BioControl 2014, 59, 427–435. [Google Scholar] [CrossRef]
- Pekas, A.; De Craecker, I.; Boonen, S.; Wäckers, F.L.; Moerkens, R. One Stone; Two Birds: Concurrent Pest Control and Pollination Services Provided by Aphidophagous Hoverflies. Biol. Control 2020, 149, 104328. [Google Scholar] [CrossRef]
- Pineda, A.; Marcos-García, M.A. Evaluation of Several Strategies to Increase the Residence Time of Episyrphus Balteatus (Diptera, Syrphidae) Releases in Sweet Pepper Greenhouses. Ann. Appl. Biol. 2008, 152, 271–276. [Google Scholar] [CrossRef]
- Prado, S.G.; Jandricic, S.E.; Frank, S.D. Ecological Interactions Affecting the Efficacy of Aphidius colemani in Greenhouse Crops. Insects 2015, 6, 538–575. [Google Scholar] [CrossRef] [Green Version]
- Jandricic, S.E.; Wraight, S.P.; Gillespie, D.R.; Sanderson, J.P. Biological Control Outcomes Using the Generalist Aphid Predator Aphidoletes aphidimyza under Multi-Prey Conditions. Insects 2016, 7, 75. [Google Scholar] [CrossRef]
- La-Spina, M.; Jandricic, S.E.; Buitenhuis, R. Short-Term Increases in Aphid Dispersal from Defensive Dropping Do Not Necessarily Affect Long-Term Biological Control by Parasitoids. J. Econ. Entomol. 2019, 112, 1552–1559. [Google Scholar] [CrossRef]
- Fischer, S.; Léger, A. Lutte biologique contre les pucerons du concombre en serre au moyen de plantes banques. Rev. Suisse Vitic. Arboric. Hortic. 1997, 29, 119–126. [Google Scholar]
- Boll, R.; Geria, A.; Marconi, A.; Migliore, O.; Salles, M.; Fauvergue, X. Contre Les Pucerons En Serres de Concombre, Les Plantes-Relais: Une Solution de Lutte Biologique? Phytoma Déf. Végétaux 2001, 536, 40–44. [Google Scholar]
- Garzo, E.; Diaz, B.; Fereres, A. Settlement Rate of Aphis gossypii (Hemiptera, Aphididae) and Transmission Efficiency of Cucumber Mosaic Virus in Melons Protected with Kaolin-Particle Films. Span. J. Agric. Res. 2003, 1, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Polat Akköprü, E. The Effect of Some Cucumber Cultivars on the Biology of Aphis gossypii Glover (Hemiptera: Aphididae). Phytoparasitica 2018, 46, 511–520. [Google Scholar] [CrossRef]
- Chi, B.; Zhang, X.; Shi, Q.; Wang, N.; Liu, Y. Colored Plastic Films Affect Demographic Characteristics of Aphis gossypii on Cucumber Plants. Int. J. Pest Manag. 2019, 65, 338–347. [Google Scholar] [CrossRef]
- Alaserhat, İ.; Canbay, A.; Özdemir, İ. Aphid Species, Their Natural Enemies in Vegetables from Erzincan, Turkey: First Record of the Parasitoid Wasp Aphelinus mali (Haldeman) Parasitizing Lipaphis erysimi (Kaltenbach). J. Agric. Sci. 2021, 27, 16–25. [Google Scholar] [CrossRef]
- Kahia, M.; Nguyen, T.; McCune, F.; Naasz, R.; Antoun, H.; Fournier, V. Insecticidal Effect of Bacillus pumilus PTB180 and Bacillus subtilis PTB185 Used Alone and in Combination against the Foxglove Aphid and the Melon Aphid (Hemiptera: Aphididae). Can. Entomol. 2021, 153, 726–740. [Google Scholar] [CrossRef]
- Payton Miller, T.L.; Rebek, E.J. Banker Plants for Aphid Biological Control in Greenhouses. J. Integr. Pest Manag. 2018, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Frank, S.D. Biological Control of Arthropod Pests Using Banker Plant Systems: Past Progress and Future Directions. Biol. Control 2010, 52, 8–16. [Google Scholar] [CrossRef]
- Huang, N.; Enkegaard, A.; Osborne, L.S.; Ramakers, P.M.J.; Messelink, G.J.; Pijnakker, J.; Murphy, G. The Banker Plant Method in Biological Control. Crit. Rev. Plant Sci. 2011, 30, 259–278. [Google Scholar] [CrossRef]
- Yano, E. Functions of Banker Plants for Biological Control of Arthropod Pests in Protected Culture. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2019, 2019, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Rezende, B.L.A.; Cecílio Filho, A.B.; Barros Júnior, A.P.; Porto, D.R.Q.; Martins, M.I.E.G. Economic Analysis of Cucumber and Lettuce Intercropping under Greenhouse in the Winter-Spring. An. Acad. Bras. Cienc. 2011, 83, 705–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cecílio Filho, A.B.; Rezende, B.L.A.; Barbosa, J.C.; Grangeiro, L.C. Agronomic Efficiency of Intercropping Tomato and Lettuce. An. Acad. Bras. Cienc. 2011, 83, 1109–1119. [Google Scholar] [CrossRef]
- Cecílio Filho, A.B.; Neto, F.B.; Rezende, B.L.A.; Barros Júnior, A.P.; de Lima, J.S.S. Indices of Bio-Agroeconomic Efficiency in Intercropping Systems of Cucumber and Lettuce in Greenhouse. Aust. J. Crop Sci. 2015, 9, 1154–1164. [Google Scholar]
- Almohamad, R.; Verheggen, F.; Haubruge, E. Searching and Oviposition Behavior of Aphidophagous Hoverflies (Diptera: Syrphidae): A Review. Biotechnol. Agron. Société Environ. 2009, 13, 467–481. [Google Scholar]
- Rodríguez-Gasol, N.; Alins, G.; Veronesi, E.R.; Wratten, S. The Ecology of Predatory Hoverflies as Ecosystem-Service Providers in Agricultural Systems. Biol. Control 2020, 151, 104405. [Google Scholar] [CrossRef]
- Bellefeuille, Y.; Fournier, M.; Lucas, E. Evaluation of Two Potential Biological Control Agents Against the Foxglove Aphid at Low Temperatures. J. Insect Sci. 2019, 19, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellefeuille, Y.; Fournier, M.; Lucas, E. Biological Control of the Foxglove Aphid Using a Banker Plant with Eupeodes americanus (Diptera: Syrphidae) in Experimental and Commercial Greenhouses. Biol. Control 2021, 155, 104541. [Google Scholar] [CrossRef]
- Ouattara, T.Y.; Fournier, M.; Rojo, S.; Lucas, E. Development Cycle of a Potential Biocontrol Agent: The American Hoverfly, Eupeodes americanus, and Comparison with the Commercial Biocontrol Agent Aphidoletes aphidimyza. Entomol. Exp. Appl. 2022, 170, 394–401. [Google Scholar] [CrossRef]
- Hemptinne, J.-L.; Doucet, J.-L.; Petersen, J.-E. Optimal Foraging by Hoverflies (Diptera: Syrphidae) and Ladybirds (Coleopteraz Coccinellidae): Mechanisms. Eur. J. Entomol. 1993, 903, 451–455. [Google Scholar]
- Pineda, A.; Morales, I.; Marcos-García, M.A.; Fereres, A. Oviposition Avoidance of Parasitized Aphid Colonies by the Syrphid Predator Episyrphus balteatus Mediated by Different Cues. Biol. Control 2007, 42, 274–280. [Google Scholar] [CrossRef]
- Almohamad, R.; Verheggen, F.J.; Francis, F.; Haubruge, E. Intraguild Interactions between the Predatory Hoverfly Episyrphus balteatus (Diptera: Syrphidae) and the Asian Ladybird, Harmonia axyridis (Coleoptera: Coccinellidae): Effect of Larval Tracks. Eur. J. Entomol. 2010, 107, 41–45. [Google Scholar] [CrossRef] [Green Version]
- Amiri-Jami, A.R.; Sadeghi, H.; Gilbert, F.; Moravvej, G.; Asoodeh, A. Oviposition Preference of Aphidophagous Hoverflies toward Oviposition Site Quality: The Presence of Intra- and Interspecific Competitor, Glucosinolate Content, and Prey Species. J. Asia-Pac. Entomol. 2016, 19, 275–280. [Google Scholar] [CrossRef]
- Dunn, L.; Lequerica, M.; Reid, C.R.; Latty, T. Dual Ecosystem Services of Syrphid Flies (Diptera: Syrphidae): Pollinators and Biological Control Agents. Pest Manag. Sci. 2020, 76, 1973–1979. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, H.; Gilbert, F. Aphid Suitability and Its Relationship to Oviposition Preference in Predatory Hoverflies. J. Anim. Ecol. 2000, 69, 771–784. [Google Scholar] [CrossRef] [Green Version]
- Almohamad, R.; Verheggen, F.J.; Francis, F.; Haubruge, E. Predatory Hoverflies Select Their Oviposition Site According to Aphid Host Plant and Aphid Species. Entomol. Exp. Appl. 2007, 125, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Vanhaelen, N.; Haubruge, E.; Gaspar, C.; Francis, F. Oviposition Preferences of Episyrphus balteatus. Meded. Rijksuniv. Te Gent Fak. Van Landbouwkd. En Toegepaste Biol. Wet. 2001, 66, 269–275. [Google Scholar]
- Messelink, G.J.; Calvo, F.J.; Marín, F.; Janssen, D. Cucurbits. In Integrated Pest and Disease Management in Greenhouse Crops; Gullino, M.L., Albajes, R., Nicot, P.C., Eds.; Plant Pathology in the 21st Century; Springer International Publishing: Cham, Switzerland, 2020; pp. 537–566. ISBN 978-3-030-22304-5. [Google Scholar]
- Messelink, G.J.; Labbé, R.; Marchand, G.; Tavella, L. Sweet Pepper. In Integrated Pest and Disease Management in Greenhouse Crops; Gullino, M.L., Albajes, R., Nicot, P.C., Eds.; Plant Pathology in the 21st Century; Springer International Publishing: Cham, Switzerland, 2020; pp. 513–535. ISBN 978-3-030-22304-5. [Google Scholar]
- Jacobson, R.J.; Croft, P. Strategies for the Control of Aphis gossypii Glover (Hom.: Aphididae) with Aphidius colemani Viereck (Hym.: Braconidae) in Protected Cucumbers. Biocontrol Sci. Technol. 1998, 8, 377–387. [Google Scholar] [CrossRef]
- Goh, H.G.; Kim, J.H.; Han, M.W. Application of Aphidius colemani Viereck for Control of the Aphid in Greenhouse. J. Asia-Pac. Entomol. 2001, 4, 171–174. [Google Scholar] [CrossRef]
- Branquart, E.; Hemptinne, J.-L. Development of Ovaries, Allometry of Reproductive Traits and Fecundity of Episyrphus balteatus (Diptera: Syrphidae). Eur. J. Entomol. 2000, 97, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Orengo-Green, J.J.; Casas, J.L.; Marcos-García, M.Á. Effect of Abiotic Climatic Factors on the Gonadal Maturation of the Biocontrol Agent Sphaerophoria rueppellii (Wiedemann, 1830) (Diptera: Syrphidae). Insects 2022, 13, 573. [Google Scholar] [CrossRef]
- Higashida, K.; Yano, E.; Nishikawa, S.; Ono, S.; Okuno, N.; Sakaguchi, T. Reproduction and Oviposition Selection by Aphidoletes aphidimyza (Diptera: Cecidomyiidae) on the Banker Plants with Alternative Prey Aphids or Crop Plants with Pest Aphids. Appl. Entomol. Zool. 2016, 51, 445–456. [Google Scholar] [CrossRef]
- Verheggen, F.J.; Capella, Q.; Schwartzberg, E.G.; Voigt, D.; Haubruge, E. Tomato-Aphid-Hoverfly: A Tritrophic Interaction Incompatible for Pest Management. Arthropod Plant Interact. 2009, 3, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Sobhani, M.; Madadi, H.; Gharali, B. Host Plant Effect on Functional Response and Consumption Rate of Episyrphus balteatus (Diptera: Syrphidae) Feeding on Different Densities of Aphis gossypii (Hemiptera: Aphididae). J. Crop Prot. 2013, 2, 375–385. [Google Scholar]
- Riddick, E.W.; Simmons, A.M. Do Plant Trichomes Cause More Harm than Good to Predatory Insects? Pest Manag. Sci. 2014, 70, 1655–1665. [Google Scholar] [CrossRef]
- Sadeghi, H. The Relationship Between Oviposition Preference and Larval Performance in an Aphidophagous Hover Fly, Syrphus ribesii L. (Diptera: Syrphidae). J. Agric. Sci. Technol. 2002, 4, 1–10. [Google Scholar]
- Chandler, A.E.F. Locomotory Behaviour of First Instar Larvae of Aphidophagous Syrphidae (Diptera) after Contact with Aphids. Anim. Behav. 1969, 17, 673–678. [Google Scholar] [CrossRef]
- Scholz, D.; Poehling, H.-M. Oviposition Site Selection of Episyrphus balteatus. Entomol. Exp. Appl. 2000, 94, 149–158. [Google Scholar] [CrossRef]
- Ambrosino, M.D.; Jepson, P.C.; Luna, J.M. Hoverfly Oviposition Response to Aphids in Broccoli Fields. Entomol. Exp. Appl. 2007, 122, 99–107. [Google Scholar] [CrossRef]
- Widstrom, N.W.; Mcmillian, W.W.; Wiseman, B.R. Ovipositional Preference of the Corn Earworm and the Development of Trichomes on Two Exotic Corn Selections. Environ. Entomol. 1979, 8, 833–839. [Google Scholar] [CrossRef]
- Durbey, S.L.; Sarup, P. Morphological Characters—Development and Density of Trichomes on Varied Maize Germplasms in Relation to Preferential Oviposition by the Stalk Borer, Chilo partellus (Swinhoe). J. Entomol. Res. 1982, 6, 187–196. [Google Scholar]
- Kumar, H. Inhibition of Ovipositional Responses of Chilo partellus (Lepidoptera: Pyralidae) by the Trichomes on the Lower Leaf Surface of a Maize Cultivar. J. Econ. Entomol. 1992, 85, 1736–1739. [Google Scholar] [CrossRef]
- Cortesero, A.M.; Stapel, J.O.; Lewis, W.J. Understanding and Manipulating Plant Attributes to Enhance Biological Control. Biol. Control 2000, 17, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Budenberg, W.J.; Powell, W. The Role of Honeydew as an Ovipositional Stimulant for Two Species of Syrphids. Entomol. Exp. Appl. 1992, 64, 57–61. [Google Scholar] [CrossRef]
- Leroy, P.D.; Almohamad, R.; Attia, S.; Capella, Q.; Verheggen, F.J.; Haubruge, E.; Francis, F. Aphid Honeydew: An Arrestant and a Contact Kairomone for Episyrphus balteatus (Diptera: Syrphidae) Larvae and Adults. Eur. J. Entomol. 2014, 111, 237–242. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, J.P.; Sullivan, M.S.; Poppy, G.M. Oviposition Behaviour and Host Colony Size Discrimination in Episyrphus balteatus (Diptera: Syrphidae). Bull. Entomol. Res. 2001, 91, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Almohamad, R.; Verheggen, F.; Francis, F.; Haubruge, E. Evaluation of Hoverfly Episyrphus balteatus De Geer (Diptera: Syrphidae) Oviposition Behaviour toward Aphid-Infested Plants Using a Leaf Disc System. Commun. Agric. Appl. Biol. Sci. 2006, 71, 403–412. [Google Scholar]
- Nelson, E.H.; Hogg, B.N.; Mills, N.J.; Daane, K.M. Syrphid Flies Suppress Lettuce Aphids. BioControl 2012, 57, 819–826. [Google Scholar] [CrossRef]
- Pu, D.; Zheng, Z.; Liu, H.; Wang, X.; Wu, X.; Chen, Y.; Deng, J.; Chen, X.; Li, Y. Development and Reproduction of the Hoverfly Eupeodes corollae (Diptera: Syrphidae). SDRP J. Earth Sci. Environ. Stud. 2019, 4, 654–660. [Google Scholar] [CrossRef]
- Parajulee, M.N. Influence of Constant Temperatures on Life History Parameters of the Cotton Aphid, Aphis gossypii, Infesting Cotton. Environ. Entomol. 2007, 36, 666–672. [Google Scholar] [CrossRef]
- Satar, S.; Kersting, U.; Uygun, N. Effect of Temperature on Population Parameters of Aphis gossypii Glover and Myzus persicae (Sulzer) (Homoptera: Aphididae) on Pepper. J. Plant Dis. Prot. 2008, 115, 69–74. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez, N.; Fauteux, A.; Louis, J.-C.; Buitenhuis, R.; Lucas, E. Oviposition Preference of the American Hoverfly, Eupeodes americanus, between Banker Plants and Target Crops. Insects 2023, 14, 295. https://doi.org/10.3390/insects14030295
Gonzalez N, Fauteux A, Louis J-C, Buitenhuis R, Lucas E. Oviposition Preference of the American Hoverfly, Eupeodes americanus, between Banker Plants and Target Crops. Insects. 2023; 14(3):295. https://doi.org/10.3390/insects14030295
Chicago/Turabian StyleGonzalez, Noémie, Arlette Fauteux, Jean-Christophe Louis, Rosemarije Buitenhuis, and Eric Lucas. 2023. "Oviposition Preference of the American Hoverfly, Eupeodes americanus, between Banker Plants and Target Crops" Insects 14, no. 3: 295. https://doi.org/10.3390/insects14030295
APA StyleGonzalez, N., Fauteux, A., Louis, J. -C., Buitenhuis, R., & Lucas, E. (2023). Oviposition Preference of the American Hoverfly, Eupeodes americanus, between Banker Plants and Target Crops. Insects, 14(3), 295. https://doi.org/10.3390/insects14030295