

Is Every Vein a Real Vein? Cross-Section of the Wing of Matsucoccus Pini (Insecta, Hemiptera, Coccoidea: Matsucoccidae)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
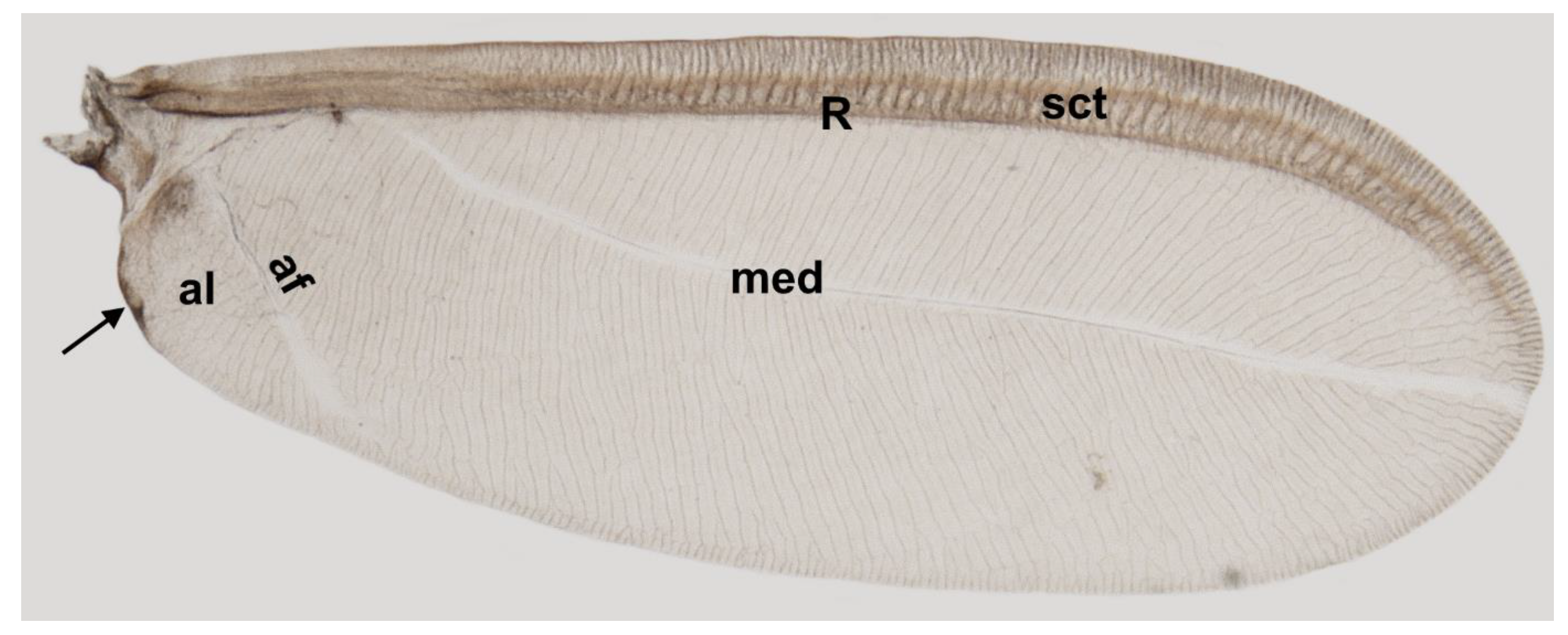
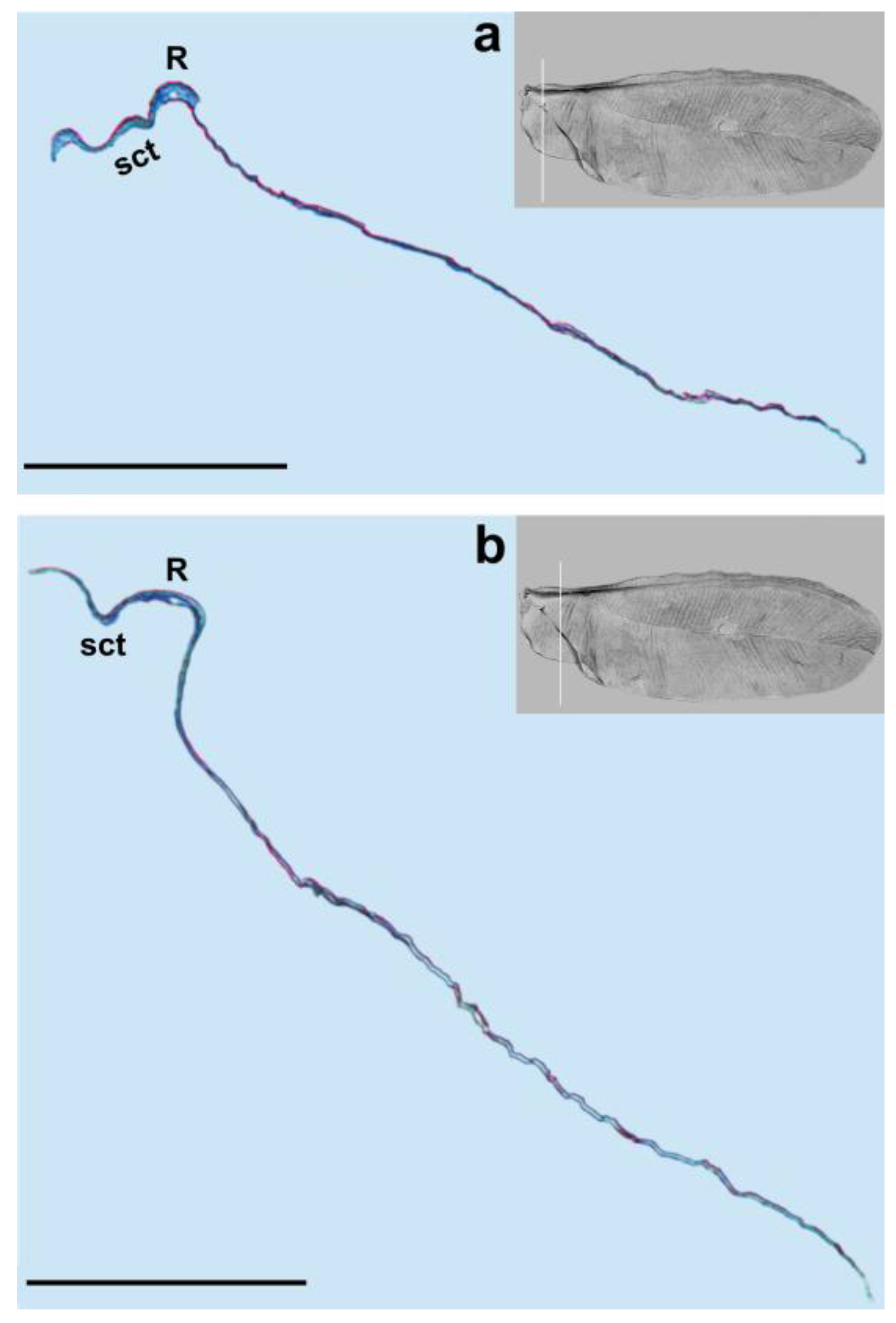
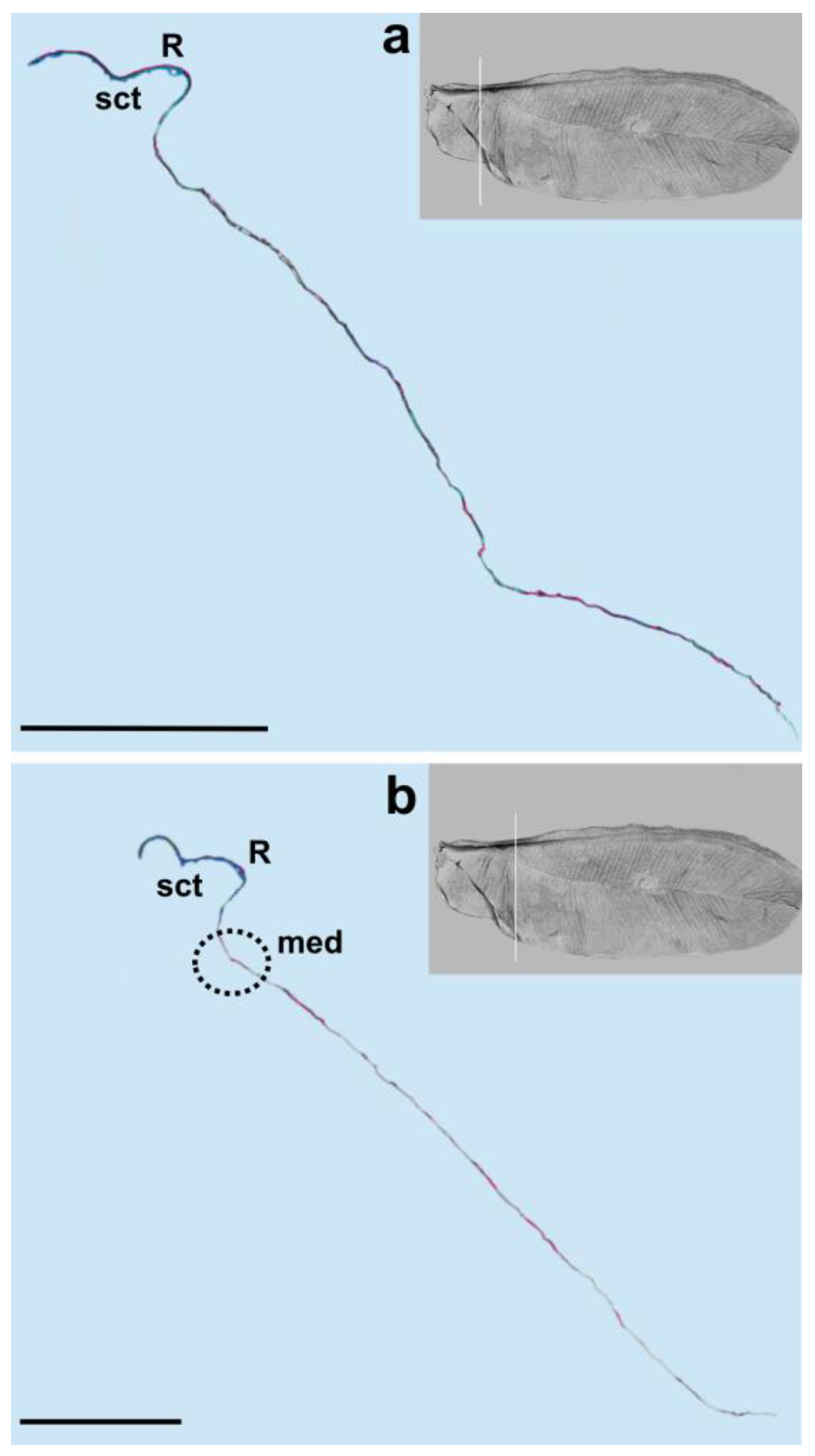
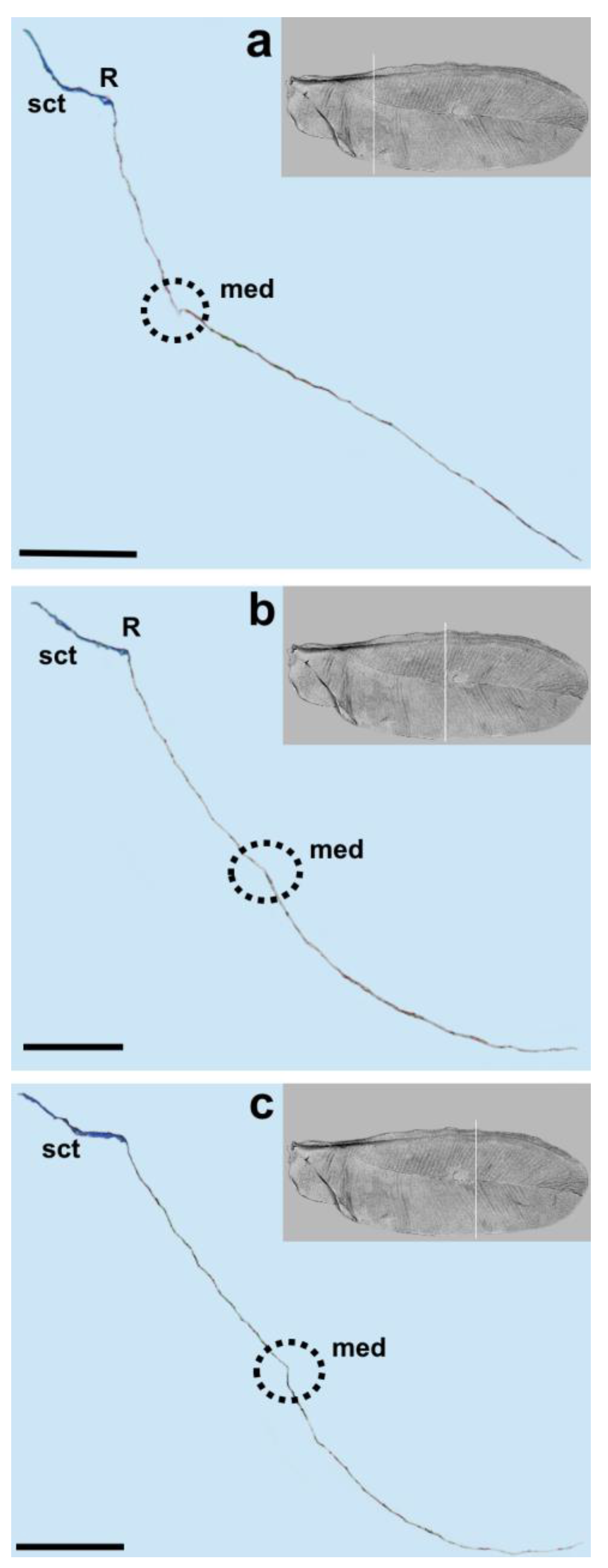
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- García Morales, M.; Denno, B.D.; Miller, D.R.; Miller, G.L.; Ben-Dov, Y.; Hardy, N.B. ScaleNet: A literature-based model of scale insect biology and systematics. Database 2016, 2016, bav118. [Google Scholar] [CrossRef] [PubMed]
- Gullan, P.J.; Cook, L.G. Phylogeny and higher classification of the scale insects (Hemiptera: Sternorrhyncha: Coccoidea). Zootaxa 2007, 1668, 413–425. [Google Scholar] [CrossRef]
- Gullan, P.J.; Martin, J.H. Sternorrhyncha (jumping plant-lice, whiteflies, aphids, and scale insects). In Encyclopedia of Insects, 2nd ed.; Resh, V.H., Cardé, R.T., Eds.; Elsevier: San Diego, CA, USA, 2009; pp. 1087–1089. [Google Scholar]
- Cook, L.G.; Gullan, P.J.; Trueman, H.E. A preliminary phylogeny of the scale insects (Hemiptera: Sternorrhyncha: Coccoidea) based on nuclear small-subunit ribosomal DNA. Mol. Phylogenet. Evol. 2002, 25, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Koteja, J.; Azar, D. Scale insects from Lower Cretaceous amber of Lebanon (Hemiptera: Sternorrhyncha: Coccinea). Alavesia 2008, 2, 133–167. [Google Scholar]
- Gavrilov-Zimin, I.A. Ontogenesis, morphology and higher classification of archacococcids (Homoptera: Coccinea: Orthezoiidea); Zoosystematica Rossica, Zoological Institute Russian Academy od Sciences: St. Petersburg, Russia, 2018; Volume 2, pp. 1–264. [Google Scholar]
- Gavrilov-Zimin, I.A.; Danzig, E.M. Taxonomic position of the genus Puto Signoret (Homoptera: Coccinea: Pseudococcidae) and separation of higher taxa in Coccinea. Zoosyst. Ross. 2012, 21, 97–111. [Google Scholar] [CrossRef]
- Danzig, E.M.; Gavrilov-Zimin, I.A. Fauna of Russia and Neighbouring Countries, New series, No. 148. In Palaearctic Mealybugs (Homoptera: Coccinea: Pseudococcidae); Zoological Institute, Russian Academy of Sciences: St. Petersburg, Russia, 2015; pp. 54–55. [Google Scholar]
- Gullan, P.J.; Kosztarab, M.P. Adaptations in scale insects. Annu. Rev. Entomol. 1997, 42, 23–50. [Google Scholar] [CrossRef]
- Koteja, J. Essay on the prehistory of the scale insects (Homoptera, Coccinea). Ann. Zool. 1985, 38, 461–504. [Google Scholar]
- Danzig, E.M. Coccoids of the Far East USSR (Homoptera, Coccinea). In Phylogenetic Analysis of Scale Insects Fauna of the World; Amerind Pub. Co. for U.S.D.A.: New Delhi, India, 1986; p. 450. [Google Scholar]
- Kawecki, Z. On the suitable term for the second pair of wings in male scale insects (Homoptera-Coccoidea). Frustula Entomol. Pisa 1965, 7, 1–4. [Google Scholar]
- Wu, S.A.; Xu, H. Nomenclature of the veins of the fore wings of male scale insects (Hemiptera, Coccomorpha). ZooKeys 2022, 1136, 163–174. [Google Scholar] [CrossRef]
- Morrison, H. A classification of the higher groups and genera of the coccid family Margarodidae. United States Dep. Agric. Tech. Bull. 1928, 52, 1–239. [Google Scholar]
- Koteja, J. Comparative studies on the labium in the Coccinea (Homoptera). Zesz. Nauk. Akad. Rol. W Warszawie Rozpr. Nauk. 1974, 89, 1–162. [Google Scholar]
- Hodgson, C.J.; Foldi, I. A review of the Margarodidae sensu Morrison (Hemiptera: Coccoidea) and some related taxa based on the morphology of adult males. Zootaxa 2006, 1263, 1–250. [Google Scholar] [CrossRef]
- Watson, G.W. Towards identification of the scale insects (Hemiptera: Coccomorpha) of continental Africa: 2. Checklists and keys to six archaeococcoid families. Zootaxa 2022, 5105, 301–356. [Google Scholar] [CrossRef]
- Foldi, I. 2005 The Matsucoccidae in the Mediterranean basin with a world list of species (Hemiptera: Sternorrhyncha: Coccoidea). Ann. Soc. Entomol. Fr. 2004, 40, 145–168. [Google Scholar] [CrossRef]
- Koteja, J. The Baltic amber Matsucoccidae (Homoptera, Coccinea). Ann. Zool. 1984, 37, 437–496. [Google Scholar]
- Beardsley, J.W., Jr. External morphology of the adult male of Matsucoccus bisetosus. Ann. Entomol. Soc.Am. 1968, 61, 1449–1459. [Google Scholar] [CrossRef]
- Siewniak, M. Kiefernborkenschildlaus Zur Morphologie und Bionomie der Kiefernborkenschildlaus Matsucoccus pini (Green) (Hom., Coccoidea: Margarodidae). On the morphology and bionomy of the pine scale Matsucoccus pini (Green) (Hom., Coccoidea: Margarodidae). Z. Angew. Entomol. 1976, 81, 337–362. [Google Scholar] [CrossRef]
- Rieux, R. Matsucoccus pini Green (1925) (Homoptera, Margarodidae) dans le Sud-Est de la France. Variations intraspécifiques. Comparaison avec des espèces les plus proches. Ann. Zool. Ecol. Anim. 1976, 8, 231–263. [Google Scholar]
- Patch, E.M. Homologies of the wing veins of the Aphididae, Psyllidae, Aleurodidae and Coccidae. Ann. Entomol. Soc. Amer. 1909, 2, 101–135. [Google Scholar] [CrossRef]
- Dudley, R. Morphology of the flight apparatus. In The Biomechanics of Insect Flight: Form, Function, Evolution; Princeton University Press: Princeton, NJ, USA, 2000; pp. 36–74. [Google Scholar]
- Franielczyk-Pietyra, B.; Depa, Ł.; Wegierek, P. Morphological and histological study of the forewing of Orthezia urticae (Linnaeus 1758) (Hemiptera, Sternorrhyncha). Zookeys 2018, 747, 101–114. [Google Scholar] [CrossRef]
- Kanturski, M.; Karcz, J.; Wieczorek, K. Morphology of the European species of the aphid genus Eulachnus (Hemiptera: Aphididae: Lachnidae)—A SEM comparative and integrative study. Micron 2015, 76, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Koteja, J. Scale insects (Homoptera: Coccinea) a day after. In Studies on Hemipteran Phylogeny; Proceedings of Thomas Say Publications in Entomology; Schaefer, C.W., Ed.; Entomological Society of America: Lanham, MD, USA, 1996; pp. 65–88. [Google Scholar]
- Koteja, J. Matsucoccidae (Homoptera, Coccinea), living fossils. Boll. Lab. Entomol. Agrar. Portici. 1987, 43, 41–44. [Google Scholar]
- Afifi, S.A. Morphology and Taxonomy of the Adult Males of the Families Pseudococcidae and Eriococcidae (Homoptera: Coccoidea); 2nd Report on Economic Zoology; British Museum (Natural History): London, UK, 1968; Volume 13, pp. 3–210. [Google Scholar]
- Koteja, J. Notes on the Polish scale insect fauna (Homoptera, Coccinea) VI. Pol. Pismo. Entomol. 1986, 56, 217–219. [Google Scholar]
- Shcherbakov, D.E. Extinct four-winged precoccids and the ancestry of scale insects and aphids (Hemiptera). Russ. Entomol. J. 2007, 16, 47–62. [Google Scholar]
- Koteja, J. Morphology and taxonomy of male Ortheziidae (Homoptera, Coccinea). Polskie Pismo Entomologiczne. Pol. J. Entomol. 1986, 56, 323–374. [Google Scholar]
- Koteja, J. Scale insects (Homoptera, Coccinea) from Upper Cretaceous New Jersey amber. In Studies on Fossils in Amber, with Particular Reference to the Cretaceous New Jersey; Grimaldi, D., Ed.; Backhuys: Leiden, The Netherlands, 2000; pp. 147–229. [Google Scholar]
- Choi, J.; Cha, D.; Kim, D.S.; Lee, S. Review of Japanese Pine Bast Scale, Matsucoccus matsumurae (Kuwana) (Coccomorpha: Matsucoccidae), Occurring on Japanese Black Pine (Pinus thunbergii Parl.) and Japanese Red Pine (P. densiflora Siebold & Zucc.) from Korea. Forests 2019, 10, 639. [Google Scholar] [CrossRef]
- Koteja, J. Matsucoccus saxonicus sp. n. from Saxonian amber (Homoptera, Coccinea). Dtsch. Ent. Z. 1986, 33, 55–63. [Google Scholar] [CrossRef]
- Hodgson Ch, J.; Hardy, N.B. The phylogeny of the superfamily Coccoidea (Hemiptera: Sternorrhyncha) based on the morphology of extant and extinct macropterous males. Syst. Entomol. 2013, 38, 794–804. [Google Scholar] [CrossRef]
- Koteja, J. Xylococcidae and related groups (Hemiptera: Coccinea) from Baltic amber. Pr. Muz. Ziemi 2008, 49, 19–56. [Google Scholar]
- Lambdin, P.L. Morphology of the second instars of Polea martini and Polea selangorae (Coccoidea: Asterolecaniidae). Entomol. News 2001, 112, 167–172. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
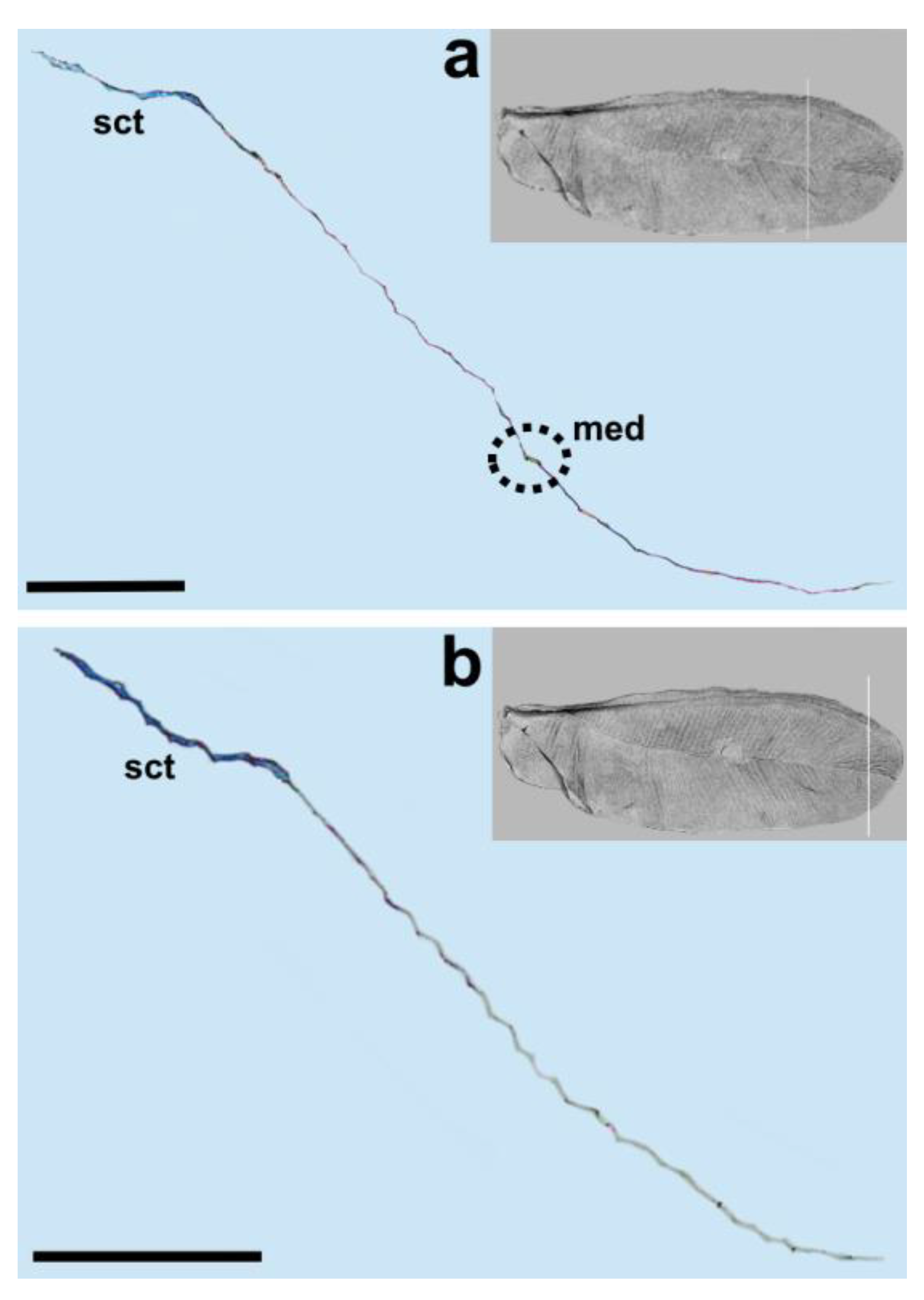{kind=link}
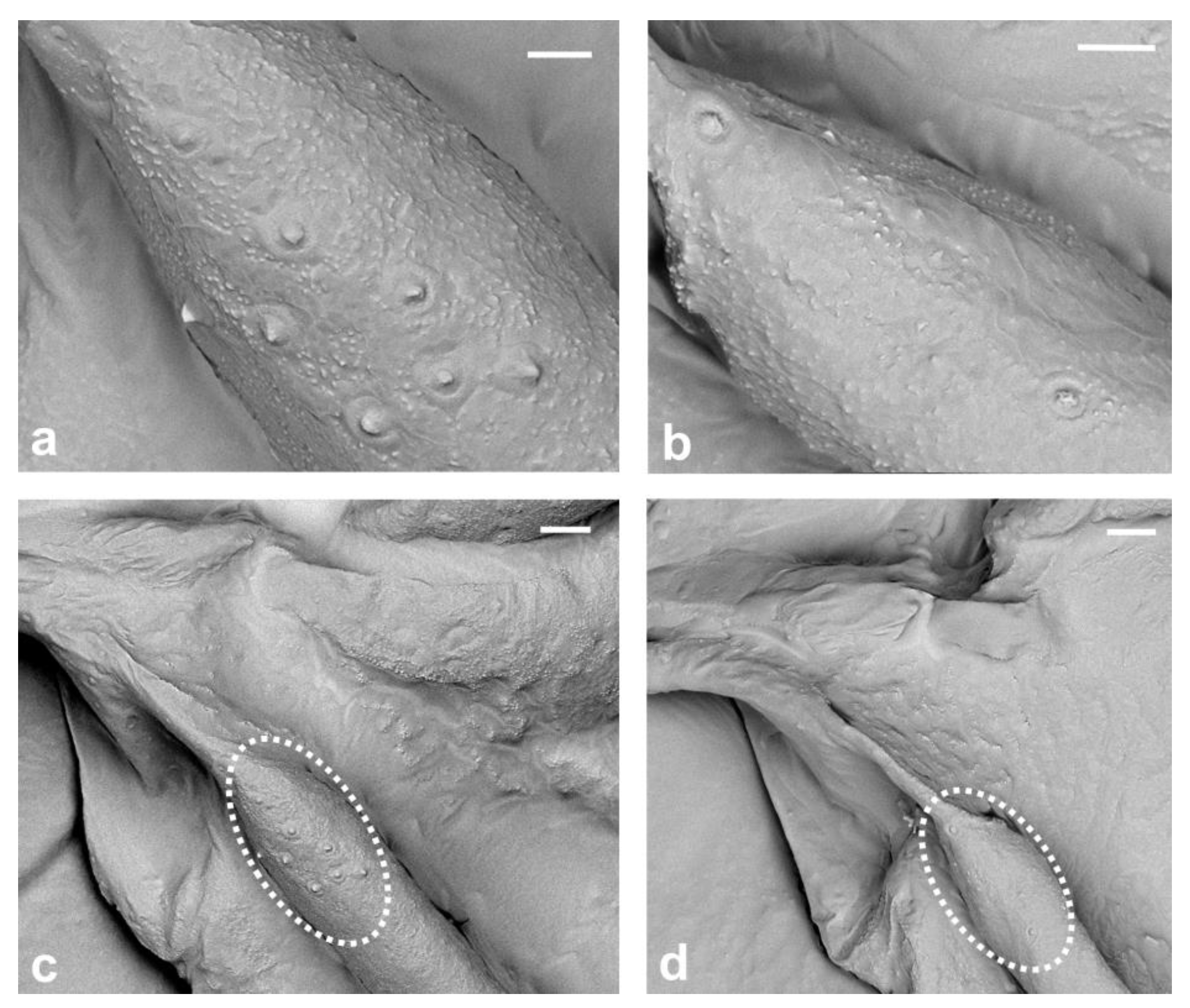{kind=link}
| Author | Taxa | Veins and Lines | |||||
|---|---|---|---|---|---|---|---|
| Morrison (1928) | Margarodidae s.l. | costal complex (Sc + R) | Apical diagonal vein (RS) | 1st diagonal light line (MS) | Basal diagonal vein (Cu + CuS) | 2nd light line (1AS) | |
| Morrison (1928) | Matsucoccus matsumurae | costal complex (Sc + R) | Single diagonal vein (Cu + CuS) | Posterior light fold | |||
| Beardsley (1968) | Matsucoccus bisetosus | subcostal thickening (sct)/s.c. | Radius (r) | Media (m) | Anal fold (af) | ||
| Siewniak (1976) | Matsucoccus pini | Subcosta (sc) | Radius (rad) | Media (med) | Analis (al) | ||
| Koteja (1984) | Fossil Matsucoccus | Subcostal thickening (sct) | Radius (r) | Media (m) | Anal vein (af) | ||
| Foldi (2005) | Matsucoccus feytaudi | Subcostal (sc) | Radial (r) | Median (m) | Anal fold (af) | ||
| Hodgson and Foldi (2006) | Matsucoccus josephi | Subcostal thickening (sclt) | Radius (rad) | Media (med) | Anal fold (af) | ||
| Wu and Xu (2022) | Matsucoccus bisetosus | Subcosta (Sc) | Radius (R) | Media (M) | Anal fold (Af) | ||
| Franielczyk- Pietyra et al. (2023) | Matsucoccus pini | subcostal thickening (sct) | Radius (R) | median fold (med) | anal fold (af) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franielczyk-Pietyra, B.; Kalandyk-Kołodziejczyk, M.; Drohojowska, J. Is Every Vein a Real Vein? Cross-Section of the Wing of Matsucoccus Pini (Insecta, Hemiptera, Coccoidea: Matsucoccidae). Insects 2023, 14, 390. https://doi.org/10.3390/insects14040390
Franielczyk-Pietyra B, Kalandyk-Kołodziejczyk M, Drohojowska J. Is Every Vein a Real Vein? Cross-Section of the Wing of Matsucoccus Pini (Insecta, Hemiptera, Coccoidea: Matsucoccidae). Insects. 2023; 14(4):390. https://doi.org/10.3390/insects14040390
Chicago/Turabian StyleFranielczyk-Pietyra, Barbara, Małgorzata Kalandyk-Kołodziejczyk, and Jowita Drohojowska. 2023. "Is Every Vein a Real Vein? Cross-Section of the Wing of Matsucoccus Pini (Insecta, Hemiptera, Coccoidea: Matsucoccidae)" Insects 14, no. 4: 390. https://doi.org/10.3390/insects14040390
APA StyleFranielczyk-Pietyra, B., Kalandyk-Kołodziejczyk, M., & Drohojowska, J. (2023). Is Every Vein a Real Vein? Cross-Section of the Wing of Matsucoccus Pini (Insecta, Hemiptera, Coccoidea: Matsucoccidae). Insects, 14(4), 390. https://doi.org/10.3390/insects14040390