Secondary Rhinaria Contribute to Major Sexual Dimorphism of Antennae in the Aphid Semiaphis heraclei (Takahashi)


 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Insects
2.2. Sample Observation, Photography and Morphometric Measurement
2.2.1. Light Microscopy
2.2.2. Scanning Electron Microscopy
2.3. Data Collection and Statistics
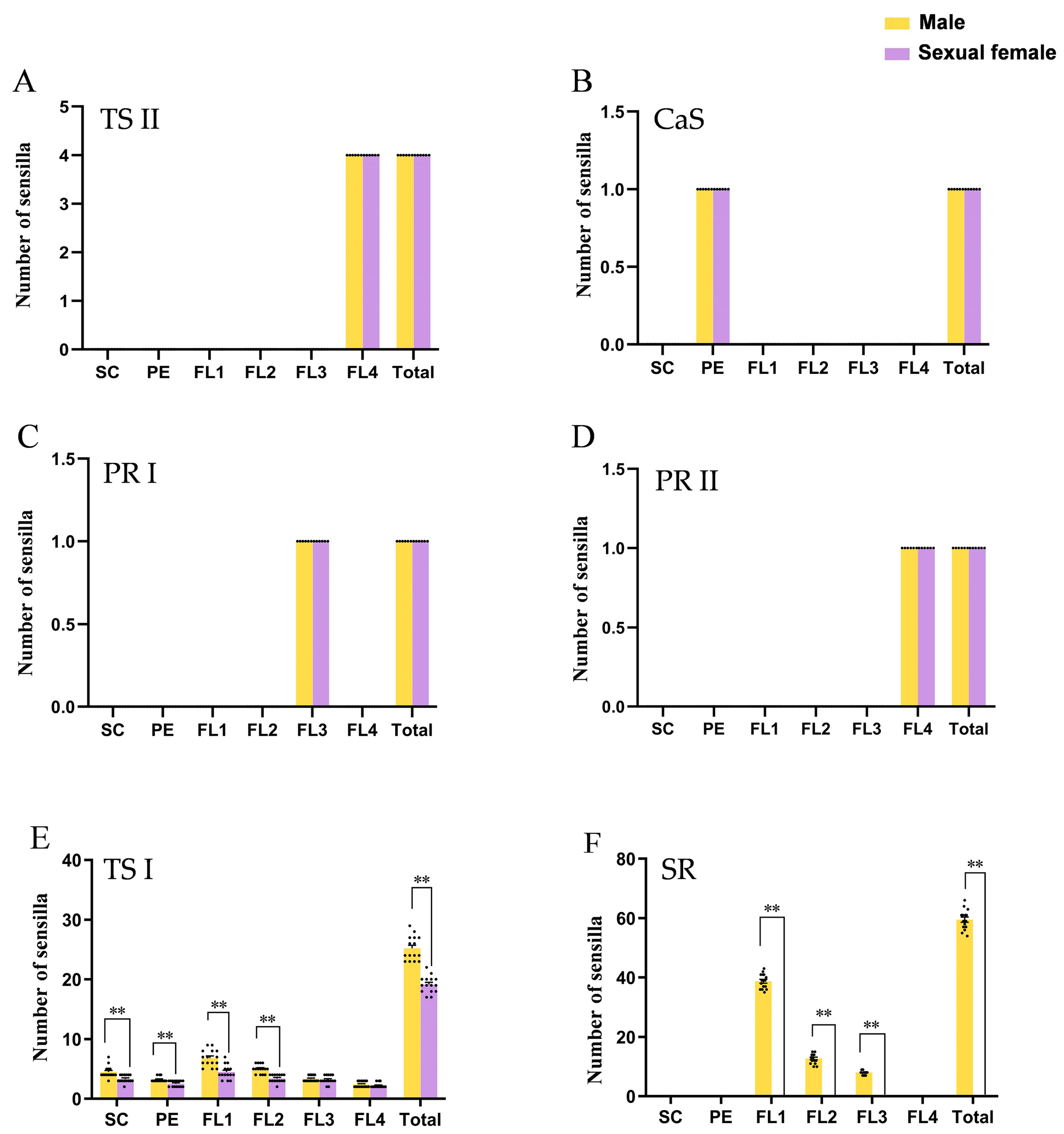
3. Results
3.1. Flagellum Length Differentiation Contributed the Majority of Sexual Dimorphism of Antennae
3.2. Types and Morphology of Antennal Sensilla
- Trichoid sensilla
- Campaniform sensilla
- Primary rhinaria
- Secondary rhinaria
3.3. Sensilla Sizes Were Universally Enlarged in Males
3.4. Secondary Rhinaria Contributed the Most to Sexual Dimorphism
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Emden, H.F.; Harrington, R. Aphids as Crop Pests, 2nd ed.; Cabi Press: Boston, MA, USA, 2017; pp. 1–40. [Google Scholar]
- Wieczorek, K. Sexuales of aphids (Insecta, Hemiptera, Aphididae)—An alternative target in the pest control. Entomol. Ornithol. Herpetol. 2016, 5, e115. [Google Scholar] [CrossRef]
- Blackman, R.L.; Sorin, M.; Miyazaki, M. Sexual morphs and colour variants of Aphis (formerly Toxoptera) odinae (Hemiptera, Aphididae) in Japan. Zootaxa 2011, 3110, 53–60. [Google Scholar] [CrossRef]
- Kanturski, M.; Wieczorek, K.; Junkiert, L. Sexual morphs and biology of Aphis Verbasci Schrank (Hemiptera: Aphididae). Zootaxa 2014, 3755, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Casiraghi, A.; Moreno-González, V.; Pérez Hidalgo, N. Description of sexuales of Brachycolus cucubali (Passerini, 1863) (Hemiptera Aphididae). Redia 2020, 103, 47–53. [Google Scholar]
- Campbell, C.A.M.; Tregidga, E.L. Photoperiodic determination of gynoparae and males of damson-hop aphid Phorodon humuli. Physiol. Entomol. 2005, 30, 189–194. [Google Scholar] [CrossRef]
- Matsuda, N.; Kanbe, T.; Endo, J.; Akimoto, S.I.; Numata, H. Suppression of autumnal sexual morph production in spring by a seasonal timer in an aphid. Physiol. Entomol. 2020, 45, 103–109. [Google Scholar] [CrossRef]
- Ngakan, P.O.; Yukawa, J. Reproduction site preference and performance by sexuparae, and mating behavior of their sexual generation on the primary host plant in a heteroecious aphid, Neothoracaphis yanonis (Homoptera: Aphididae). Entomol. Sci. 2021, 24, 68–75. [Google Scholar] [CrossRef]
- Byers, J.A. Modelling female mating success during mass trapping and natural competitive attraction of searching males or females. Entomol. Exp. Appl. 2012, 145, 228–237. [Google Scholar] [CrossRef]
- Stewart-Jones, A.; Dewhirst, S.Y.; Durrant, L.; Fitzgerald, J.D.; Hardie, J.; Hooper, A.M.; Pickett, J.A.; Poppy, G.M. Structure, ratios and patterns of release in the sex pheromone of an aphid, Dysaphis plantaginea. Exp. Biol. 2007, 210, 4335–4344. [Google Scholar] [CrossRef]
- Dewhirst, S.Y.; Pickett, J.A.; Hardie, J. Aphid Pheromones. In Vitamins and Hormones; Litwack, G., Ed.; Academic Press: Pittsburgh, PA, USA, 2010; Volume 83, pp. 551–574. [Google Scholar]
- Murano, K.; Ogawa, K.; Kaji, T.; Miura, T. Pheromone gland development and monoterpenoid synthesis specific to oviparous females in the pea aphid. Zool. Lett. 2018, 4, 9. [Google Scholar] [CrossRef]
- Symmes, E.J.; Dewhirst, S.Y.; Birkett, M.A.; Campbell, C.A.M.; Chamberlain, K.; Pickett, J.A.; Zalom, F.G. The sex pheromones of mealy plum (Hyalopterus pruni) and leaf-curl plum (Brachycaudus helichrysi) aphids: Identification and field trapping of male and gynoparous aphids in prune orchards. Chem. Ecol. 2012, 38, 576–583. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.A.M.; Hardie, J.; Wadhams, L.J. Attraction range of a sex pheromone trap for the damson-hop aphid Phorodon humuli (Hemiptera: Aphididae). Physiol. Entomol. 2017, 42, 389–396. [Google Scholar] [CrossRef]
- Schneider, D. Insect antennae. Annu. Rev. Entomol. 1964, 9, 103–122. [Google Scholar] [CrossRef]
- Wu, Y.T.; Wu, M.C.; Hui, Z.; Hu, X.S.; Xu, X.L. Polyphenism in antennal sensilla among different adult morphs of nonhost-alternating, holocyclic Sitobion avenae (Hemiptera: Aphididae). J. Insect Sci. 2022, 22, 4. [Google Scholar] [CrossRef]
- Kanturski, M.; Świątek, P.; Trela, J.; Borowiak-Sobkowiak, B.; Wieczorek, K. Micromorphology of the model species pea aphid Acyrthosiphon pisum (Hemiptera, Aphididae) with special emphasis on the sensilla structure. Eur. Zool. J. 2020, 87, 336–356. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, Z.N. Comparative study of antennal sensilla of various morphs of Myzus persicae. Acta Entomol. Sin. 2000, 43, 131–136, 215. [Google Scholar]
- Du, Y.J.; Yan, F.S.; Tang, J. Structure, and function of olfactory sensilla on the antennae of soybean aphids, Aphis glycines. Acta Entomol. Sin. 1995, 38, 1–7. [Google Scholar]
- Liu, F.Z.; Ling, B.; Lv, X.X.; Luo, J.M.; Fan, Z.T. The microstructure of antennal sensoria of cotton aphid under scanning electron microscope. J. Bayi Agric. Coll. 1989, 4, 42–43, 47, 89. Available online: https://kns.cnki.net/kcms2/article/abstract?v=3uoqIhG8C44YLTlOAiTRKqd0WnNPv0wTDjtDUwHroNwQ0q_GxjX5o0p-8M0sOLAcql7BzFxErgZh7CH5X8fTOoGcv5c3PZTP&uniplatform=NZKPT (accessed on 21 December 2022).
- Zhang, G.X.; Zhong, T.S. Economic Insects of China, Vol 25 Homoptera Aphids, 1st ed.; Science Press: Beijing, China, 1983; p. 387. [Google Scholar]
- Yeh, Y.C.; Doan, L.H.; Huang, Z.Y.; Chu, L.W.; Shi, T.H.; Lee, Y.R.; Wu, C.T.; Lin, C.H.; Chiang, S.T.; Liu, H.K. Honeysuckle (Lonicera japonica) and Huangqi (Astragalus membranaceus) suppress SARS-CoV-2 entry and COVID-19 related cytokine storm in vitro. Front. Pharmacol. 2022, 12, 765553. [Google Scholar] [CrossRef]
- Wu, Q.H.; Qiu, X.; Zeng, T.X.; Huang, L.F.; Liao, W.; Pei, J. Analysis of key varieties and rational allocation of traditional Chinese medicine resources under COVID-19 epidemic situation. Chin. J. ETMF 2020, 26, 84–91. [Google Scholar]
- Chen, J.; Ding, W.L.; Cheng, H.Z. Medicinal Plant Protection; Publishing House of Electronics Industry: Beijing, China, 2019; p. 727. [Google Scholar]
- Shi, Z.H.; Song, K.X.; Wang, J.Y.; Li, M.X.; Qiao, H.L.; Guo, K.; Xu, C.Q. Morphological characteristics for identifying the instars of Semiaphis heraclei. Chin. J. Appl. Entomol. 2022, 59, 457–465. [Google Scholar]
- Zhang, Y.; Wu, Y.K.; Liu, Y.S.; Li, L.Y. Research on population dynamics of Lonicera macranthoides aphid and natural enemy in Xiushan and evolution of pesticides. Chin. J. Tradit. Chin. Med. 2012, 37, 3219–3222. [Google Scholar]
- Sun, Y. Occurrence and Comprehensive Control of Main Pests of Honeysuckle in Shandong Province. Master’s Thesis, Shandong Agricultural University, Tai’an, China, 2013. [Google Scholar]
- Staudacher, E.M.; Gebhardt, M.; Dürr, V. Antennal movements and mechanoreception: Neurobiology of active tactile sensors. In Advances in Insect Physiology; Simpson, S.J., Ed.; Academic Press: Pittsburgh, PA, USA, 2005; Volume 32, pp. 49–205. [Google Scholar]
- Zhong, J.; Guo, Y.H.; Xu, T.; Li, D.W.; Sun, M. Observation on the morphological characteristics for age identification of Macrosiphoniella sanborni Hemiptera Aphididae and the ultrastructure of its antennal sensilla. J. Acta Entomol. Sin. 2019, 62, 1315–1324. [Google Scholar]
- Bruno, D.; Grossi, G.; Salvia, R.; Scala, A.; Farina, D.; Grimaldi, A.; Zhou, J.J.; Bufo, S.A.; Vogel, H.; Grosse-Wilde, E.; et al. Sensilla morphology and complex expression pattern of odorant binding proteins in the vetch aphid Megoura viciae (Hemiptera: Aphididae). Front. Physiol. 2018, 9, 777. [Google Scholar] [CrossRef] [PubMed]
- Bromley, A.K.; Dunn, J.A.; Anderson, M. Ultrastructure of the antennal sensilla of aphids. II Trichoid, chordotonal and campaniform sensilla. Cell Tissue Res. 1980, 205, 493–511. [Google Scholar] [CrossRef]
- Sun, Y.P.; Zhao, L.J.; Sun, L.; Zhang, S.G.; Ban, L.P. Immunolocalization of odorant-binding proteins on antennal chemosensilla of the peach aphid Myzus persicae (Sulzer). Chem. Senses 2013, 38, 129–136. [Google Scholar] [CrossRef]
- Park, K.C.; Hardie, J. Electrophysiological characterization of olfactory sensilla in the black bean aphid, Aphis fabae. J. Insect Physiol. 2004, 50, 647–655. [Google Scholar] [CrossRef]
- Ban, L.P.; Sun, Y.P.; Wang, Y.; Tu, X.B.; Zhang, S.G.; Zhang, Y.T.; Wu, Y.S.; Zhang, Z.H. Ultrastructure of antennal sensilla of the peach aphid Myzus persicae Sulzer, 1776. J. Morphol. 2014, 276, 219–227. [Google Scholar] [CrossRef]
- Bromley, A.K.; Dunn, J.A.; Anderson, M. Ultrastructure of the antennal sensilla of aphids. I. Coeloconic and placoid sensilla. Cell Tissue Res. 1979, 203, 427–442. [Google Scholar] [CrossRef]
- Campbell, C.A.M.; Dawson, G.W.; Griffiths, D.C.; Pettersson, J.; Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M. Sex attractant pheromone of damson-hop aphid Phorodon humuli (Homopter, Aphididae). J. Chem. Ecol. 1990, 16, 3455–3465. [Google Scholar] [CrossRef]
- Guldemond, J.A.; Dixon, A.F.G.; Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M. Specificity of sex pheromones, the role of host plant odour in the olfactory attraction of males, and mate recognition in the aphid Cryptomyzus. Physiol. Entomol. 1993, 18, 137–143. [Google Scholar] [CrossRef]
- Dawson, G.W.; Griffiths, D.C.; Merritt, L.A.; Mudd, A.; Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M. Aphid semiochemicals—A review and recent advances on the sex pheromone. J. Chem. Ecol. 1990, 16, 3019–3030. [Google Scholar] [CrossRef] [PubMed]
- Kanturski, M.; Ali Akbar, S.; Favret, C. The Bhutan pine aphid Pseudessigella brachychaeta Hille Ris Lambers (Hemiptera: Aphididae: Lachninae) from India reveals the hitherto unknown oviparous female and dwarfish Male. Zool. Stud. 2017, 56, e12. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sensilla | Sensilla within Rhinaria | Wall | Socket | Tip | Index | Male | Sexual Female | t Value | p Value |
|---|---|---|---|---|---|---|---|---|---|
| TS I | / | Smooth | Raised | Slender | L BD | 14.40 ± 0.49 ** 1.15 ± 0.03 | 10.01 ± 0.34 1.14 ± 0.02 | 7.39 0.18 | <0.01 0.86 |
| TS II | / | Smooth | Raised | Blunt and grooved | L BD | 6.13 ± 0.14 1.15 ± 0.01 ** | 5.95 ± 0.06 1.12 ± 0.04 | 1.15 −3.13 | 0.26 <0.01 |
| CaS | / | Smooth | Wide | Button-like | D | 3.22 ± 0.04 ** | 3.02 ± 0.04 | 3.47 | <0.01 |
| PR I | PS | Rough | Wide | Discoid | LD SD | 14.61 ± 0.32 ** 11.72 ± 0.24 ** | 9.70 ± 0.17 8.57 ± 0.23 | 13.61 9.32 | <0.01 <0.01 |
| The whole | Rough | Wide | Discoid | LD SD | 20.97 ± 0.20 ** 15.09 ± 0.24 | 18.55 ± 0.14 13.75 ± 0.21 | 9.79 4.12 | <0.01 <0.01 | |
| PR II | LPS | Rough | Wide | Discoid | LD SD | 9.69 ± 0.20 ** 6.66 ± 0.27 ** | 6.39 ± 0.08 5.70 ± 0.15 | 15.25 3.09 | <0.01 <0.01 |
| SPS | Rough | Wide | Discoid | LD SD | 3.22 ± 0.06 2.51 ± 0.07 ** | 3.14 ± 0.06 2.43 ± 0.04 | 0.95 −3.73 | 0.35 <0.01 | |
| CoS | Smooth | Sunken | Finger-like | D | 5.19 ± 0.06 ** | 4.66 ± 0.04 | 7.57 | <0.01 | |
| SR | PS | Rough | Wide | Discoid | LD SD | 13.71 ± 0.76 8.49 ± 0.27 | / / | 20.35 36.18 | <0.01 <0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, K.-X.; Wang, J.-Y.; Qiao, H.-L.; Wei, H.-S.; Guo, K.; Xu, C.-Q. Secondary Rhinaria Contribute to Major Sexual Dimorphism of Antennae in the Aphid Semiaphis heraclei (Takahashi). Insects 2023, 14, 468. https://doi.org/10.3390/insects14050468
Song K-X, Wang J-Y, Qiao H-L, Wei H-S, Guo K, Xu C-Q. Secondary Rhinaria Contribute to Major Sexual Dimorphism of Antennae in the Aphid Semiaphis heraclei (Takahashi). Insects. 2023; 14(5):468. https://doi.org/10.3390/insects14050468
Chicago/Turabian StyleSong, Ke-Xin, Jiang-Yue Wang, Hai-Li Qiao, Hong-Shuang Wei, Kun Guo, and Chang-Qing Xu. 2023. "Secondary Rhinaria Contribute to Major Sexual Dimorphism of Antennae in the Aphid Semiaphis heraclei (Takahashi)" Insects 14, no. 5: 468. https://doi.org/10.3390/insects14050468
APA StyleSong, K. -X., Wang, J. -Y., Qiao, H. -L., Wei, H. -S., Guo, K., & Xu, C. -Q. (2023). Secondary Rhinaria Contribute to Major Sexual Dimorphism of Antennae in the Aphid Semiaphis heraclei (Takahashi). Insects, 14(5), 468. https://doi.org/10.3390/insects14050468