
Tissue-Specific Response to Experimental Demethylation at Seed Germination in the Non-Model Herb Erodium cicutarium


Abstract
:1. Introduction
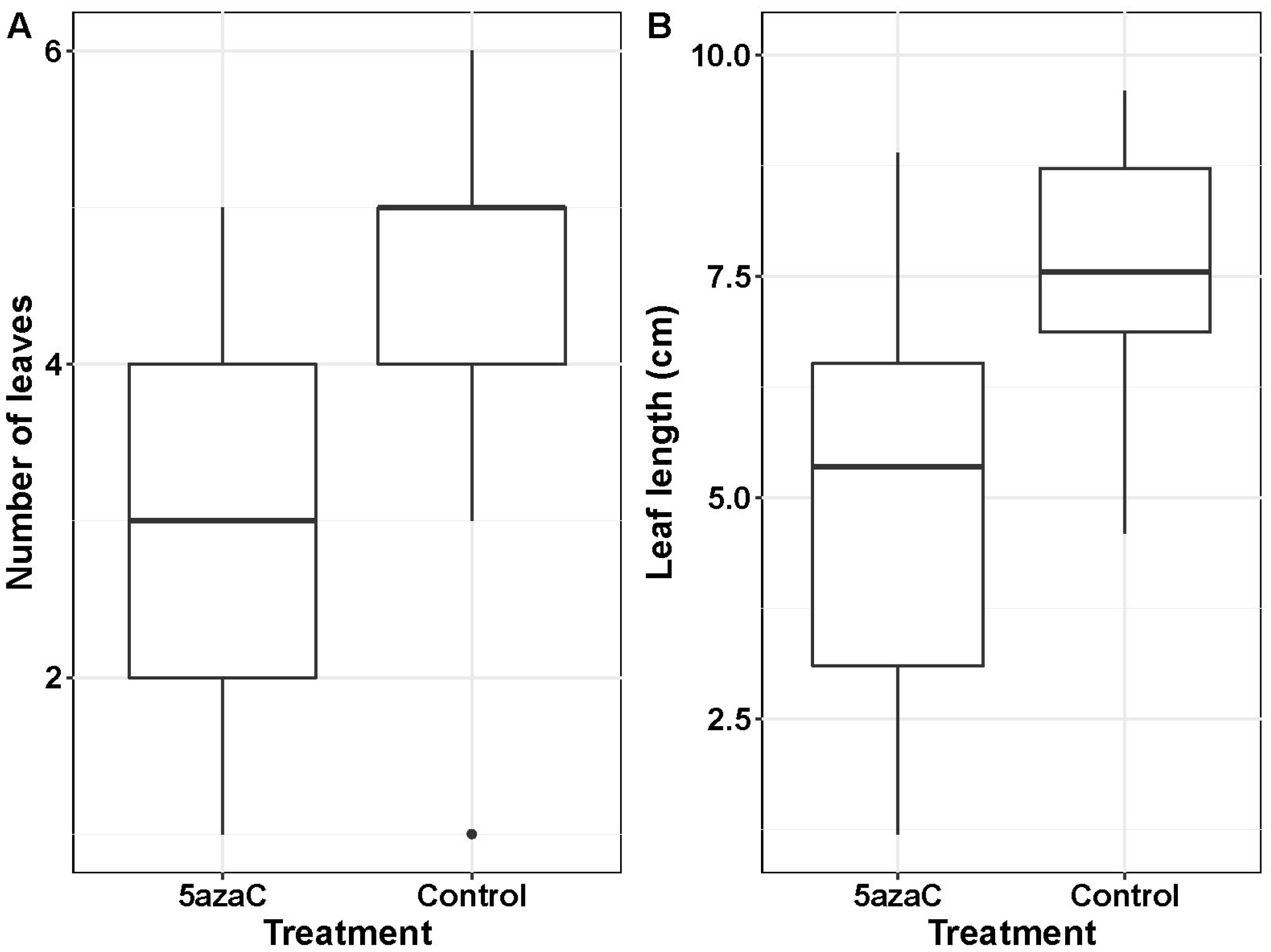
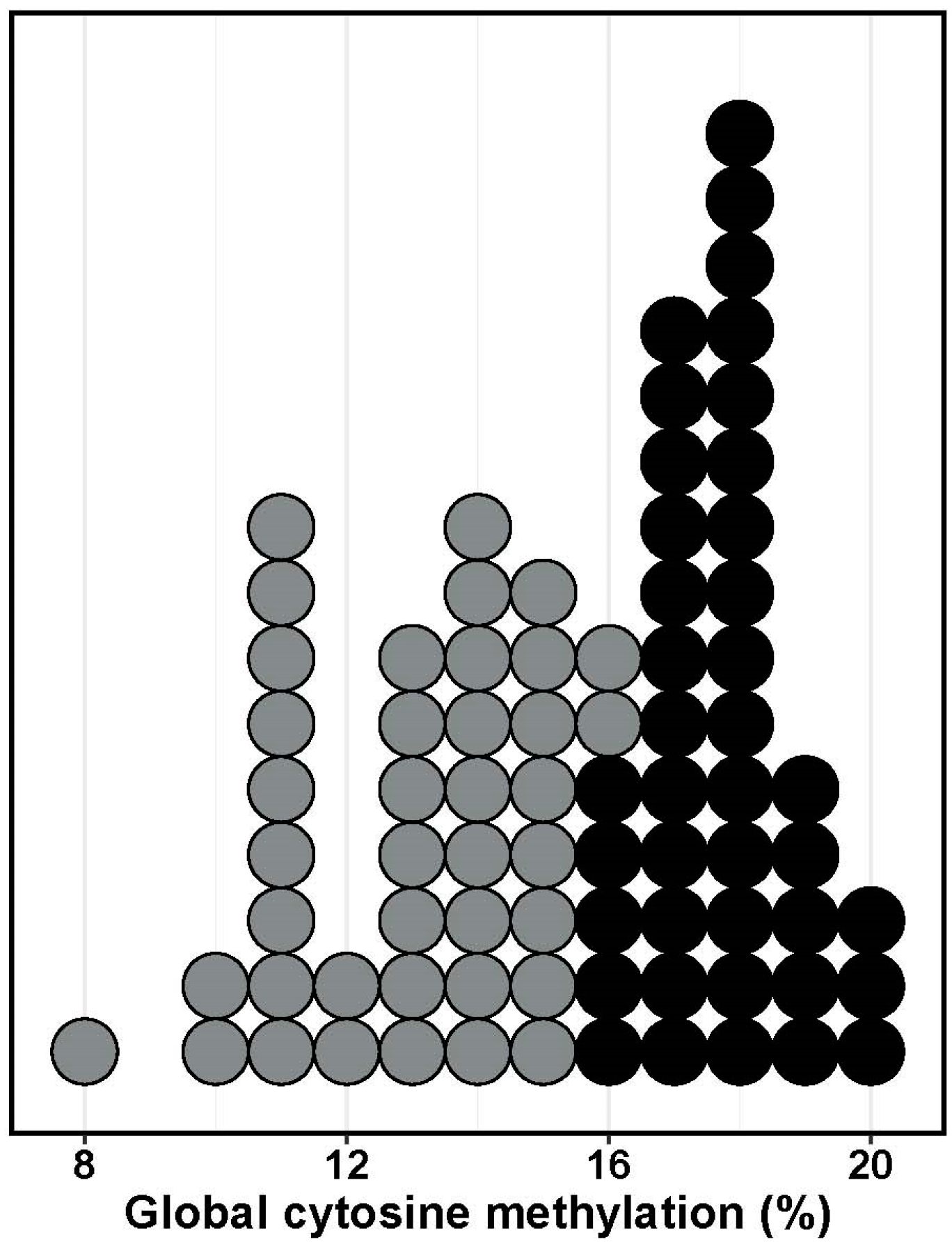
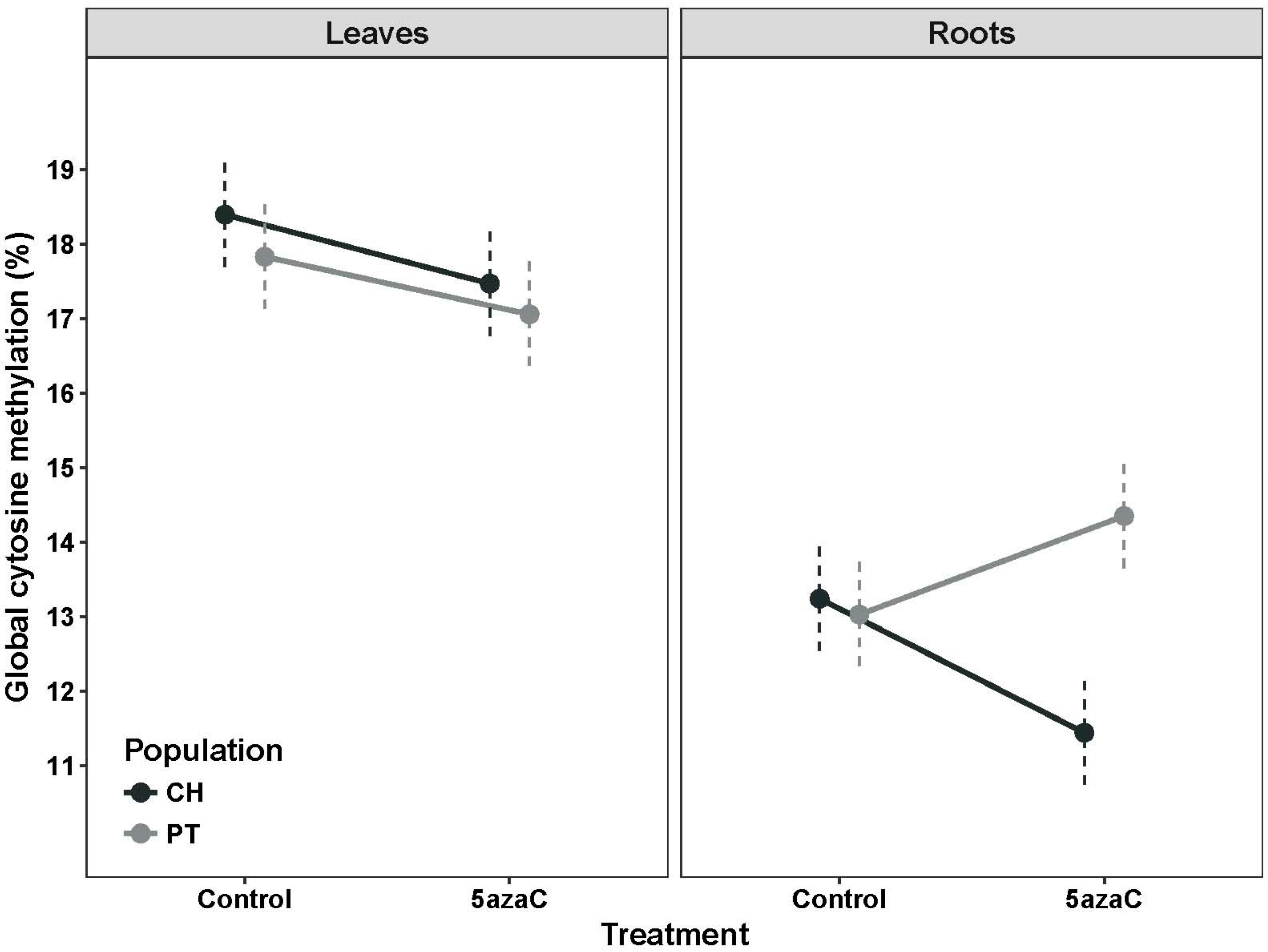
2. Results
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Laboratory Methods
4.3. Statistical Analyses
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Finnegan, E.J. DNA methylation: A dynamic regulator of genome organization and gene expression in plants. In Plant Developmental Biology—Biotechnological Perspectives: Volume 2; Pua, E.-C., Davey, M.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 2, pp. 295–323. [Google Scholar]
- Feil, R.; Fraga, M. Epigenetics and the environment: Emerging patterns and implications. Nat. Rev. Genet. 2012, 13, 97–109. [Google Scholar] [CrossRef]
- Seymour, D.K.; Koenig, D.; Hagmann, J.; Becker, C.; Weigel, D. Evolution of DNA methylation patterns in the Brassicaceae is driven by differences in genome organization. PLoS Genet. 2014, 10, e1004785. [Google Scholar] [CrossRef]
- Burggren, W. Epigenetic inheritance and its role in evolutionary biology: Re-evaluation and new perspectives. Biology 2016, 5. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.K. Epigenetic regulation of stress responses in plants. Curr. Opin. Plant Biol. 2009, 12, 133–139. [Google Scholar] [CrossRef]
- Peng, H.; Zhang, J. Plant genomic DNA methylation in response to stresses: Potential applications and challenges in plant breeding. Prog. Nat. Sci. 2009, 19, 1037–1045. [Google Scholar] [CrossRef]
- Mirouze, M.; Paszkowski, J. Epigenetic contribution to stress adaptation in plants. Curr. Opin. Plant Biol. 2011, 14, 267–274. [Google Scholar] [CrossRef]
- Herman, J.J.; Sultan, S.E. Adaptive transgenerational plasticity in plants: Case studies, mechanisms, and implications for natural populations. Front. Plant Sci. 2011, 2. [Google Scholar] [CrossRef]
- Walter, J.; Jentsch, A.; Beierkuhnlein, C.; Kreyling, J. Ecological stress memory and cross stress tolerance in plants in the face of climate extremes. Environ. Exp. Bot. 2013, 94, 3–8. [Google Scholar] [CrossRef]
- Pikaard, C.S.; Mittelsten Scheid, O. Epigenetic regulation in plants. Cold Spring Harb. Perspect. Biol. 2014, 6, a019315. [Google Scholar] [CrossRef]
- Alonso, C.; Pérez, R.; Bazaga, P.; Medrano, M.; Herrera, C.M. MSAP markers and global cytosine methylation in plants: A literature survey and comparative analysis for a wild growing species. Mol. Ecol. Resour. 2016, 16, 80–90. [Google Scholar] [CrossRef]
- Pecinka, A.; Mittelsten Scheid, O. Stress-induced chromatin changes: A critical view on their heritability. Plant Cell Physiol. 2012, 53, 801–808. [Google Scholar] [CrossRef]
- Lopez, M.; Halby, L.; Arimondo, P.B. DNA methyltransferase inhibitors: Development and applications. In DNA Methyltransferases—Role and Function; Jeltsch, A., Jurkowska, R.Z., Eds.; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar]
- Sano, H.; Kamada, I.; Youssefian, S.; Katsumi, M.; Wabiko, H. A single treatment of rice seedlings with 5-azacytidine induces heritable dwarfism and undermethylation of genomic DNA. Mol. Gen. Genet. 1990, 220, 441–447. [Google Scholar] [CrossRef]
- Janousek, B.; Siroky, J.; Vyskot, B. Epigenetic control of sexual phenotype in a dioecious plant, Melandrium album. Mol. Gen. Genet. 1996, 250, 483–490. [Google Scholar] [CrossRef]
- Fieldes, M.A.; Amyot, L.M. Evaluating the potential of using 5-azacytidine as an epimutagen. Can. J. Bot. 1999, 77, 1617–1622. [Google Scholar] [CrossRef]
- Kondo, H.; Shiraya, T.; Wada, K.C.; Takeno, K. Induction of flowering by DNA demethylation in Perilla frutescens and Silene armeria: Heritability of 5-azacytidine-induced effects and alteration of the DNA methylation state by photoperiodic conditions. Plant Sci. 2010, 178, 321–326. [Google Scholar] [CrossRef]
- Xu, J.; Tanino, K.K.; Robinson, S.J. Stable epigenetic variants selected from an induced hypomethylated Fragaria vesca population. Front. Plant Sci. 2016, 7, 1768. [Google Scholar] [CrossRef]
- Baubec, T.; Pecinka, A.; Rozhon, W.; Mittelsten Scheid, O. Effective, homogeneous and transient interference with cytosine methylation in plant genomic DNA by zebularine. Plant J. 2009, 57, 542–554. [Google Scholar] [CrossRef]
- Griffin, P.T.; Niederhuth, C.E.; Schmitz, R.J. A comparative analysis of 5-azacytidine- and zebularine-induced DNA demethylation. Genes Genomes Genet. 2016, 6, 2773–2780. [Google Scholar] [CrossRef]
- Bossdorf, O.; Arcuri, D.; Richards, C.L.; Pigliucci, M. Experimental alteration of DNA methylation affects the phenotypic plasticity of ecologically relevant traits in Arabidopsis thaliana. Evol. Ecol. 2010, 24, 541–553. [Google Scholar] [CrossRef]
- Verhoeven, K.J.F.; van Gurp, T.P. Transgenerational effects of stress exposure on offspring phenotypes in apomictic dandelion. PLoS ONE 2012, 7, e38605. [Google Scholar] [CrossRef]
- Herman, J.J.; Sultan, S.E. DNA methylation mediates genetic variation for adaptive transgenerational plasticity. Proc. R. Soc. B 2016, 283, 20160988. [Google Scholar] [CrossRef]
- Rendina González, A.P.; Chrtek, J.; Dobrev, P.I.; Dumalasová, V.; Fehrer, J.; Mráz, P.; Latzel, V. Stress-induced memory alters growth of clonal off spring of white clover (Trifolium repens). Am. J. Bot. 2016, 103, 1567–1574. [Google Scholar] [CrossRef]
- Cao, D.; Gao, X.; Liu, J.; Kimatu, J.N.; Geng, S.; Wang, X.; Zhao, J.; Shi, D. Methylation sensitive amplified polymorphism (MSAP) reveals that alkali stress triggers more DNA hypomethylation levels in cotton (Gossypium hirsutum L.) roots than salt stress. Afr. J. Biotechnol. 2011, 10, 18971–18980. [Google Scholar]
- Cao, D.; Gao, X.; Liu, J.; Wang, X.; Geng, S.; Yang, C.; Liu, B.; Shi, D. Root-specific DNA methylation in Chloris virgata, a natural alkaline-resistant halophyte, in response to salt and alkaline stresses. Plant Mol. Biol. Rep. 2012, 30, 1102–1109. [Google Scholar] [CrossRef]
- Greco, M.; Chiappetta, A.; Bruno, L.; Bitonti, M.B. Effects of light deficiency on genome methylation in Posidonia oceanica. Mar. Ecol. Prog. Ser. 2013, 473, 103–114. [Google Scholar] [CrossRef]
- Karan, R.; DeLeon, T.; Biradar, H.; Subudhi, P.K. Salt stress induced variation in DNA methylation pattern and its influence on gene expression in contrasting rice genotypes. PLoS ONE 2012, 7, e40203. [Google Scholar] [CrossRef]
- Fiz-Palacios, O.; Vargas, P.; Vila, R.; Papadopulos, A.S.T.; Aldasoro, J.J. The uneven phylogeny and biogeography of Erodium (Geraniaceae): Radiations in the Mediterranean and recent recurrent intercontinental colonization. Ann. Bot. 2010, 106, 871–884. [Google Scholar] [CrossRef]
- Francis, A.; Darbyshire, S.J.; Légère, A.; Simard, M.-J. The biology of Canadian weeds. 151. Erodium cicutarium (L.) L’Hér. Ex Aiton. Can. J. Plant Sci. 2012, 92, 1359–1380. [Google Scholar] [CrossRef]
- Blackshaw, R.E. Soil, temperature, soil moisture, and seed burial depth effects on redstem filaree (Erodium cicutarium) emergence. Weed Sci. 1992, 40, 204–207. [Google Scholar]
- Cox, J.A.; Conran, J.G. The effect of water stress on the life cycles of Erodium crinitum Carolin and Erodium cicutarium (L.) L’Herit. ex Aiton (Geraniaceae). Aust. J. Ecol. 1996, 21, 235–240. [Google Scholar] [CrossRef]
- Heger, T.; Jacobs, B.S.; Latimerb, A.M.; Johannes, K.; Kevin, J.R. Does experience with competition matter? Effects of source competitive environment on mean and plastic trait expression in Erodium cicutarium. Perspect. Plant Ecol. Evol. Syst. 2014, 16, 236–246. [Google Scholar] [CrossRef]
- Kimball, S.; Gremer, J.R.; Barron-Gafford, G.A.; Angert, A.L.; Huxman, T.E.; Venable, D.L. High water-use efficiency and growth contribute to success of non-native Erodium cicutarium in a Sonoran desert winter annual community. Conserv. Physiol. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Paparella, S.; Araújo, S.S.; Rossi, G.; Wijayasinghe, M.; Carbonera, D.; Balestrazzi, A. Seed priming: State of the art and new perspectives. Plant Cell Rep. 2015, 34, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Leck, M.A.; Parker, V.T.; Simpson, R.L. Seedling Ecology and Evolution; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Grant-Downton, R.T.; Dickinson, H.G. Epigenetics and its implications for plant biology 2. The ’Epigenetic Epiphany’: Epigenetics, evolution and beyond. Ann. Bot. 2006, 97, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Jablonka, E.; Raz, G. Transgenerational epigenetic inheritance: Prevalence, mechanisms, and implications for the study of heredity and evolution. Q. Rev. Biol. 2009, 84, 131–176. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, S.; Baumberger, R.; Grossniklaus, U. Epigenetic variation, inheritance, and selection in plant populations. Cold Spring Harb. Symp. Quant. Biol. 2013, 77, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Richards, C.L.; Verhoeven, K.J.F.; Bossdorf, O. Evolutionary significance of epigenetic variation. In Plant Genome Diversity; Wendel, J., Greilhuber, J., Dolezel, J., Leitch, I., Eds.; Springer: Vienna, Austria, 2012; Volume 1, pp. 257–274. [Google Scholar]
- Fieldes, M.A. Heritable effects of 5-azacytidine treatments on the growth and development of flax (Linum usitatissimum) genotrophs and genotypes. Genome 1994, 37, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Tanino, K.K.; Horner, K.N.; Robinson, S.J. Quantitative trait variation is revealed in a novel hypomethylated population of woodland strawberry (Fragaria vesca). BMC Plant Biol. 2016, 16, 240–257. [Google Scholar] [CrossRef] [PubMed]
- Amoah, S.; Kurup, S.; Rodriguez Lopez, C.M.; Welham, S.J.; Powers, S.J.; Hopkins, C.J.; Wilkinson, M.J.; King, G.J. A hypomethylated population of Brassica rapa for forward and reverse epi-genetics. BMC Plant Biol. 2012, 12, 193–210. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Fei, Z.; Chen, Y.-R.; Zheng, Y.; Huang, M.; Vrebalov, J.; McQuinn, R.; Gapper, N.; Liu, B.; Xiang, J.; et al. Single-base resolution methylomes of tomato fruit development reveal epigenome modifications associated with ripening. Nat. Biotechnol. 2013, 31, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Pérez, J. Breeding system, flower visitors and seedling survival of two endangered species of Helianthemum (Cistaceae). Ann. Bot. 2005, 95, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Messeguer, R.; Ganal, M.; Steffens, J.; Tanksley, S. Characterization of the level, target sites and inheritance of cytosine methylation in tomato nuclear DNA. Plant Mol. Biol. 1991, 16, 753–770. [Google Scholar] [CrossRef] [PubMed]
- Demeulemeester, M.A.C.; Van Stallen, N.; De Proft, M.P. Degree of DNA methylation in chicory (Cichorium intybus l.): Influence of plant age and vernalization. Plant Sci. 1999, 142, 101–108. [Google Scholar] [CrossRef]
- Ruiz-García, L.; Cervera, M.T.; Martínez-Zapater, J.M. DNA methylation increases throughout Arabidopsis development. Planta 2005, 222, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.C.L.; De Decker, M.M.; Fieldes, M.A. A comparative analysis of developmental profiles for DNA methylation in 5-azacytidine-induced early-flowering flax lines and their control. Plant Sci. 2008, 175, 217–225. [Google Scholar] [CrossRef]
- Osabe, K.; Clement, J.D.; Bedon, F.; Pettolino, F.A.; Ziolkowski, L.; Llewellyn, D.J.; Finnegan, E.J.; Wilson, I.W. Genetic and DNA methylation changes in cotton (Gossypium) genotypes and tissues. PLoS ONE 2014, 9, e86049. [Google Scholar] [CrossRef] [PubMed]
- Eichten, S.R.; Vaughn, M.W.; Hermanson, P.J.; Springer, N.M. Variation in DNA methylation patterns is more common among maize inbreds than among tissues. Plant Genome 2013, 6. [Google Scholar] [CrossRef]
- Roessler, K.; Takuno, S.; Gaut, B.S. CG methylation covaries with differential gene expression between leaf and floral bud tissues of Brachypodium distachyon. PLoS ONE 2016, 11, e0150002. [Google Scholar] [CrossRef] [PubMed]
- Chwialkowska, K.; Nowakowska, U.; Mroziewicz, A.; Szarejko, I.; Kwasniewski, M. Water-deficiency conditions differently modulate the methylome of roots and leaves in barley (Hordeum vulgare L.). J. Exp. Bot. 2016, 67, 1109–1121. [Google Scholar] [CrossRef] [PubMed]
- Fieldes, M.A.; Schaeffer, S.M.; Krech, M.J.; Brown, J.C.L. DNA hypomethylation in 5-azacytidine-induced early-flowering lines of flax. Theor. Appl. Genet. 2005, 111, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Sano, H.; Kamada, I.; Youssefian, S.; Wabiko, H. Correlation between DNA undermethylation and dwarfism in maize. Biochim. Biophys. Acta 1989, 1009, 35–38. [Google Scholar] [CrossRef]
- Latzel, V. Pitfalls in ecological research—Transgenerational effects. Folia Geobot. 2015, 50, 75–85. [Google Scholar] [CrossRef]
- Lopez-Torres, A.; Yanez-Barrientos, E.; Wrobel, K.; Wrobel, K. Selective derivatization of cytosine and methylcytosine moieties with 2-bromoacetophenone for submicrogram DNA methylation analysis by reversed phase HPLC with spectrofluorimetric detection. Anal. Chem. 2011, 83, 7999–8005. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing, 3.3.3; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Crawley, M.J. The R Book; John Wiley & Sons Ltd.: Chichester, UK, 2007. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Nlme: Linear and nonlinear mixed effects models. R Found. Stat. Comput. 2017, 3, 107–156. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 2nd ed.; Sage: Thousand Oaks, CA, USA, 2011. [Google Scholar]
- Lenth, R.V. Least-squares means: The R package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Factor | χ2 | p |
|---|---|---|
| Treatment (TR) | 3.57 | 0.059 |
| Tissue (TI) | 620.59 | <0.0001 |
| Provenance (PR) | 2.30 | 0.129 |
| TR * TI | 2.45 | 0.117 |
| TR * PR | 7.94 | 0.0048 |
| TI * PR | 23.82 | <0.0001 |
| TR * TI * PR | 15.59 | <0.0001 |
| Compound | Concentration (mM) | Time (h) | Species | Analysis | Reference |
|---|---|---|---|---|---|
| 5azadC | 0.3 | 16–72 | Oryza sativa 1 | 2D-TLC | [14] |
| 5azaC | 0.3 | 16 | Zea mays 1 | 2D-TLC | [55] |
| 5azaC | 0.3 | 16–72 | Oryza sativa 1 | 2D-TLC | [14] |
| 5azaC | 0.5–1.5 | 24–72 | Linum usitatissimum | HPLC | [41] |
| 5azaC | 0.01–1.5 | 72 | Brassica rapa 2 | MSAP | [42] |
| 5azaC | 0.025–1.0 | 192 | Arabidopsis thaliana | NGS | [20] |
| 5azaC | 1.0–100 | 144 | Fragaria vesca 1,4 | MSAP | [42] |
| zebu | 0.02–0.08 | 336–504 | Arabidopsis thaliana 2 | HPLC | [19] |
| zebu | 0.04 | 168 | Medicago sativa | HPLC | [19] |
| zebu | 0.01–0.1 | 336 | Taraxacum officinale 3 | - | [22] |
| zebu | 0.045 | 144 | Polygonum persicaria | - | [23] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alonso, C.; Medrano, M.; Pérez, R.; Bazaga, P.; Herrera, C.M. Tissue-Specific Response to Experimental Demethylation at Seed Germination in the Non-Model Herb Erodium cicutarium. Epigenomes 2017, 1, 16. https://doi.org/10.3390/epigenomes1030016
Alonso C, Medrano M, Pérez R, Bazaga P, Herrera CM. Tissue-Specific Response to Experimental Demethylation at Seed Germination in the Non-Model Herb Erodium cicutarium. Epigenomes. 2017; 1(3):16. https://doi.org/10.3390/epigenomes1030016
Chicago/Turabian StyleAlonso, Conchita, Mónica Medrano, Ricardo Pérez, Pilar Bazaga, and Carlos M. Herrera. 2017. "Tissue-Specific Response to Experimental Demethylation at Seed Germination in the Non-Model Herb Erodium cicutarium" Epigenomes 1, no. 3: 16. https://doi.org/10.3390/epigenomes1030016
APA StyleAlonso, C., Medrano, M., Pérez, R., Bazaga, P., & Herrera, C. M. (2017). Tissue-Specific Response to Experimental Demethylation at Seed Germination in the Non-Model Herb Erodium cicutarium. Epigenomes, 1(3), 16. https://doi.org/10.3390/epigenomes1030016