
Making Mitotic Chromosomes in a Test Tube


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Establishment of Frog Egg Extracts for Studying Large-Scale Chromatin Structures
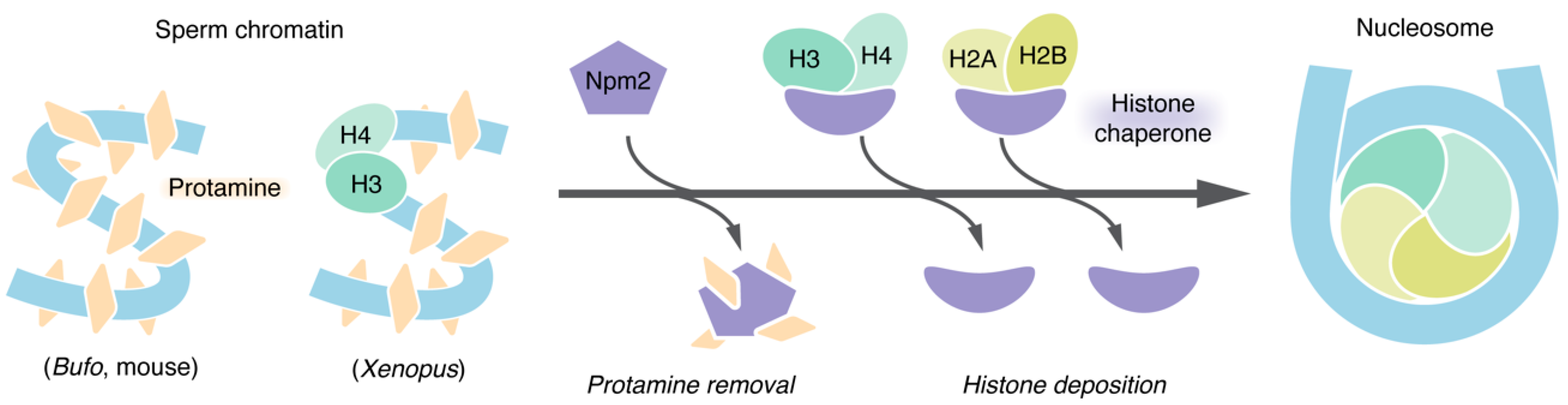
3. Sperm Chromatin Remodeling: An Essential Preparatory Step for Chromosome Assembly in In Vitro Assays
4. Mitotic Chromosome Assembly Recapitulated in Egg Extracts
5. Mitotic Chromatids Can Be Made by Using Purified Proteins
6. Our Current Understanding of Major Chromosome-Associated Proteins
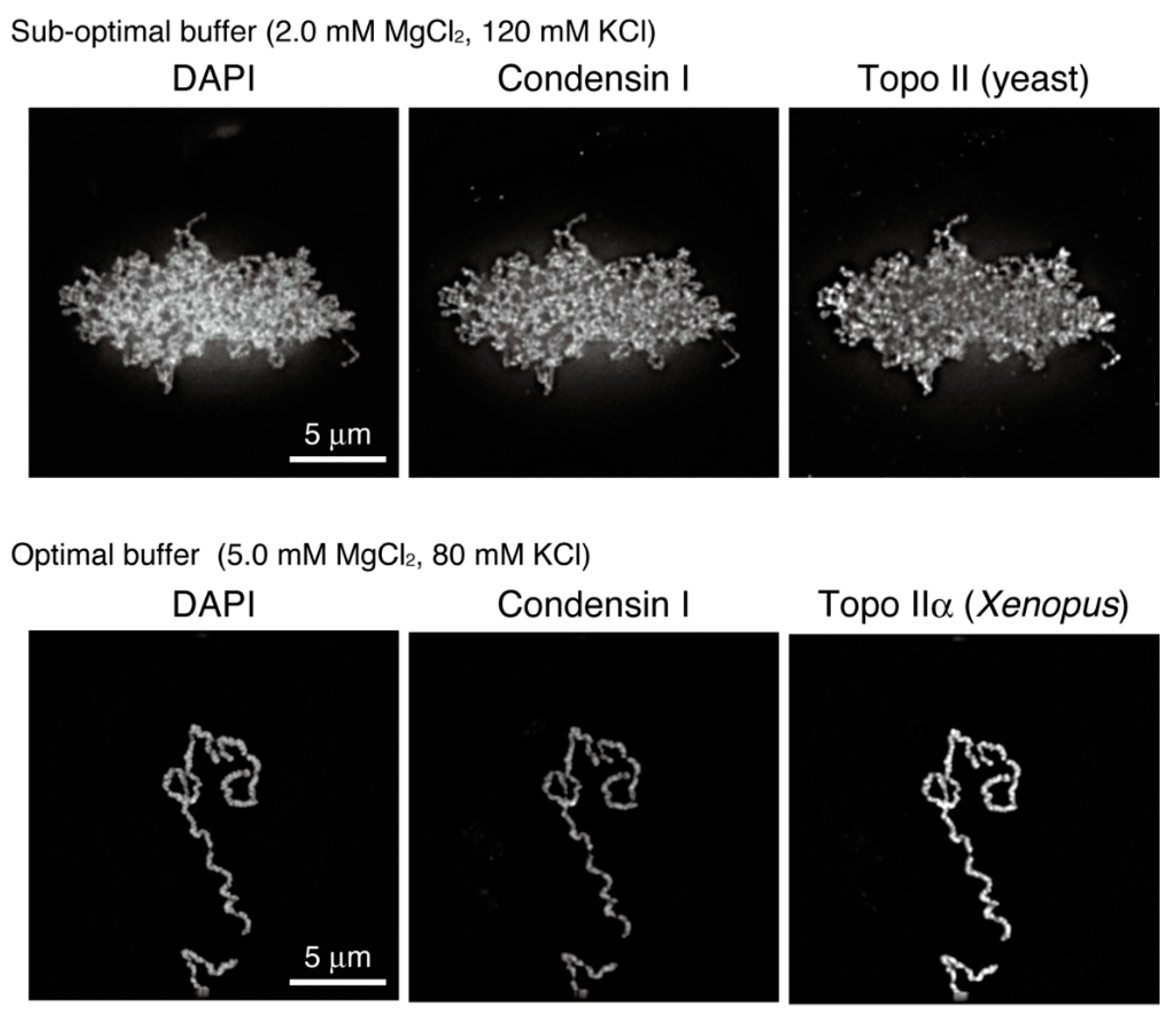
6.1. Condensins: ATP-Utilizing Machines That Fold a DNA Strand into a Chromosome
6.2. Topo IIα: A Catalyst for Disentanglement and Entanglement of Chromosomal DNA
6.3. Histones: Dynamic Bricks of Chromosomes
7. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Hirano, T.; Mitchison, T.J. A heterodimeric coiled-coil protein required for mitotic chromosome condensation in vitro. Cell 1994, 79, 449–458. [Google Scholar] [CrossRef]
- Hirano, T.; Kobayashi, R.; Hirano, M. Condensins, chromosome condensation protein complexes containing XCAP-C, XCAP-E and a Xenopus homolog of the Drosophila Barren protein. Cell 1997, 89, 511–521. [Google Scholar] [CrossRef] [Green Version]
- Shintomi, K.; Takahashi, T.S.; Hirano, T. Reconstitution of mitotic chromatids with a minimum set of purified factors. Nat. Cell Biol. 2015, 17, 1014–1023. [Google Scholar] [CrossRef]
- Lohka, M.J.; Masui, Y. Formation in vitro of sperm pronuclei and mitotic chromosomes induced by amphibian ooplasmic components. Science 1983, 220, 719–721. [Google Scholar] [CrossRef]
- Iwao, Y.; Katagiri, C. In vitro induction of sperm nucleus decondensation by cytosol from mature toad eggs. J. Exp. Zool. 1984, 230, 115–124. [Google Scholar] [CrossRef]
- Lohka, M.J.; Masui, Y. Effects of Ca2+ ions on the formation of metaphase chromosomes and sperm pronuclei in cell-free preparations from unactivated Rana pipiens eggs. Dev. Biol. 1984, 103, 434–442. [Google Scholar] [CrossRef]
- Lohka, M.J.; Maller, J.L. Induction of nuclear envelope breakdown, chromosome condensation, and spindle formation in cell-free extracts. J. Cell Biol. 1985, 101, 518–523. [Google Scholar] [CrossRef] [Green Version]
- Miake-Lye, R.; Kirschner, M.W. Induction of early mitotic events in a cell-free system. Cell 1985, 41, 165–175. [Google Scholar] [CrossRef]
- Murray, A.W. Cell cycle extracts. Methods Cell Biol. 1991, 36, 581–605. [Google Scholar]
- Jenness, C.; Wynne, D.J.; Funabiki, H. Protein Immunodepletion and Complementation in Xenopus laevis Egg Extracts. Cold Spring Harb. Protoc. 2018, 2018, pdb.prot097113. [Google Scholar] [CrossRef]
- Lohka, M.J.; Hayes, M.K.; Maller, J.L. Purification of maturation-promoting factor, an intracellular regulator of early mitotic events. Proc. Natl. Acad. Sci. USA 1988, 85, 3009–3013. [Google Scholar] [CrossRef] [Green Version]
- Blow, J.J.; Laskey, R.A. A role for the nuclear envelope in controlling DNA replication within the cell cycle. Nature 1988, 332, 546–548. [Google Scholar] [CrossRef]
- Murray, A.W.; Kirschner, M.W. Cyclin synthesis drives the early embryonic cell cycle. Nature 1989, 339, 275–280. [Google Scholar] [CrossRef]
- Murray, A.W.; Solomon, M.J.; Kirschner, M.W. The role of cyclin synthesis and degradation in the control of maturation promoting factor activity. Nature 1989, 339, 280–286. [Google Scholar] [CrossRef]
- Ohsumi, K.; Katagiri, C. Characterization of the ooplasmic factor inducing decondensation of and protamine removal from toad sperm nuclei: Involvement of nucleoplasmin. Dev. Biol. 1991, 148, 295–305. [Google Scholar] [CrossRef]
- Philpott, A.; Leno, G.H. Nucleoplasmin remodels sperm chromatin in Xenopus egg extracts. Cell 1992, 69, 759–767. [Google Scholar] [CrossRef]
- Holloway, S.L.; Glotzer, M.; King, R.W.; Murray, A.W. Anaphase is initiated by proteolysis rather than by the inactivation of maturation-promoting factor. Cell 1993, 73, 1393–1402. [Google Scholar] [CrossRef]
- Görlich, D.; Prehn, S.; Laskey, R.A.; Hartmann, E. Isolation of a protein that is essential for the first step of nuclear protein import. Cell 1994, 79, 767–778. [Google Scholar] [CrossRef]
- Kubota, Y.; Mimura, S.; Nishimoto, S.; Takisawa, H.; Nojima, H. Identification of the yeast MCM3-related protein as a component of Xenopus DNA replication licensing factor. Cell 1995, 81, 601–609. [Google Scholar] [CrossRef] [Green Version]
- Heald, R.; Tournebize, R.; Blank, T.; Sandaltzopoulos, R.; Becker, P.; Hyman, A.; Karsenti, E. Self-organization of microtubules into bipolar spindles around artificial chromosomes in Xenopus egg extracts. Nature 1996, 382, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Losada, A.; Hirano, M.; Hirano, T. Identification of Xenopus SMC protein complexes required for sister chromatid cohesion. Genes Dev. 1998, 12, 1986–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, J.; Sun, L.; Newport, J. Regulated chromosomal DNA replication in the absence of a nucleus. Mol. Cell 1998, 1, 519–529. [Google Scholar] [CrossRef]
- Ono, T.; Losada, A.; Hirano, M.; Myers, M.P.; Neuwald, A.F.; Hirano, T. Differential contributions of condensin I and condensin II to mitotic chromosome architecture in vertebrate cells. Cell 2003, 115, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Lemaitre, J.M.; Danis, E.; Pasero, P.; Vassetzky, Y.; Méchali, M. Mitotic remodeling of the replicon and chromosome structure. Cell 2005, 123, 787–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, K.; Kobayashi, T.J.; Hirano, T. Balancing acts of two HEAT subunits of condensin I support dynamic assembly of chromosome axes. Dev. Cell 2015, 33, 94–106. [Google Scholar] [CrossRef] [Green Version]
- Shintomi, K.; Inoue, F.; Watanabe, H.; Ohsumi, K.; Ohsugi, M.; Hirano, T. Mitotic chromosome assembly despite nucleosome depletion in Xenopus egg extracts. Science 2017, 356, 1284–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arimura, Y.; Shih, R.M.; Froom, R.; Funabiki, H. Structural features of nucleosomes in interphase and metaphase chromosomes. Mol. Cell 2021, 81, 4377–4397.e4312. [Google Scholar] [CrossRef]
- Kunkle, M.; Longo, F.J.; Magun, B.E. Nuclear protein changes in the maternally and paternally derived chromatin at fertilization. J. Exp. Zool. 1978, 203, 371–380. [Google Scholar] [CrossRef]
- Longo, F.J. Pronuclear events during fertilization. In Biology of Fertilization; Academic Press: New York, NY, USA, 1985; Volume 3, pp. 251–298. [Google Scholar]
- Philpott, A.; Leno, G.H.; Laskey, R.A. Sperm decondensation in Xenopus egg cytoplasm is mediated by nucleoplasmin. Cell 1991, 65, 569–578. [Google Scholar] [CrossRef]
- Ohsumi, K.; Katagiri, C. Occurrence of H1 subtypes specific to pronuclei and cleavage-stage cell nuclei of anuran amphibians. Dev. Biol. 1991, 147, 110–120. [Google Scholar] [CrossRef]
- Loppin, B.; Bonnefoy, E.; Anselme, C.; Laurencon, A.; Karr, T.L.; Couble, P. The histone H3.3 chaperone HIRA is essential for chromatin assembly in the male pronucleus. Nature 2005, 437, 1386–1390. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Zhang, Y. Nucleosome assembly is required for nuclear pore complex assembly in mouse zygotes. Nat. Struct. Mol. Biol. 2014, 21, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Zierhut, C.; Jenness, C.; Kimura, H.; Funabiki, H. Nucleosomal regulation of chromatin composition and nuclear assembly revealed by histone depletion. Nat. Struct. Mol. Biol. 2014, 21, 617–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onikubo, T.; Shechter, D. Chaperone-mediated chromatin assembly and transcriptional regulation in Xenopus laevis. Int. J. Dev. Biol. 2016, 60, 271–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammond, C.M.; Strømme, C.B.; Huang, H.; Patel, D.J.; Groth, A. Histone chaperone networks shaping chromatin function. Nat. Rev. Mol. Cell Biol. 2017, 18, 141–158. [Google Scholar] [CrossRef] [Green Version]
- Laskey, R.A.; Honda, B.M.; Mills, A.D.; Finch, J.T. Nucleosomes are assembled by an acidic protein which binds histones and transfers them to DNA. Nature 1978, 275, 416–420. [Google Scholar] [CrossRef]
- Earnshaw, W.C.; Honda, B.M.; Laskey, R.A.; Thomas, J.O. Assembly of nucleosomes: The reaction involving X. laevis nucleoplasmin. Cell 1980, 21, 373–383. [Google Scholar] [CrossRef]
- Shintomi, K.; Iwabuchi, M.; Saeki, H.; Ura, K.; Kishimoto, T.; Ohsumi, K. Nucleosome assembly protein-1 is a linker histone chaperone in Xenopus eggs. Proc. Natl. Acad. Sci. USA 2005, 102, 8210–8215. [Google Scholar] [CrossRef] [Green Version]
- Kleinschmidt, J.A.; Dingwall, C.; Maier, G.; Franke, W.W. Molecular characterization of a karyophilic, histone-binding protein: cDNA cloning, amino acid sequence and expression of nuclear protein N1/N2 of Xenopus laevis. EMBO J. 1986, 5, 3547–3552. [Google Scholar] [CrossRef]
- Dilworth, S.M.; Black, S.J.; Laskey, R.A. Two complexes that contain histones are required for nucleosome assembly in vitro: Role of nucleoplasmin and N1 in Xenopus egg extracts. Cell 1987, 51, 1009–1018. [Google Scholar] [CrossRef]
- Quivy, J.P.; Grandi, P.; Almouzni, G. Dimerization of the largest subunit of chromatin assembly factor 1: Importance in vitro and during Xenopus early development. EMBO J. 2001, 20, 2015–2027. [Google Scholar] [CrossRef] [Green Version]
- Ray-Gallet, D.; Quivy, J.P.; Scamps, C.; Martini, E.M.; Lipinski, M.; Almouzni, G. HIRA is critical for a nucleosome assembly pathway independent of DNA synthesis. Mol. Cell 2002, 9, 1091–1100. [Google Scholar] [CrossRef]
- Wuhr, M.; Freeman, R.M., Jr.; Presler, M.; Horb, M.E.; Peshkin, L.; Gygi, S.P.; Kirschner, M.W. Deep proteomics of the Xenopus laevis egg using an mRNA-derived reference database. Curr. Biol. 2014, 24, 1467–1475. [Google Scholar] [CrossRef] [Green Version]
- Shintomi, K.; Hirano, T. A Sister Chromatid Cohesion Assay Using Xenopus Egg Extracts. Methods Mol. Biol. 2017, 1515, 3–21. [Google Scholar]
- Masui, Y. A cytostatic factor in amphibian oocytes: Its extraction and partial characterization. J. Exp. Zool. 1974, 187, 141–147. [Google Scholar] [CrossRef]
- Liu, J.; Grimison, B.; Maller, J.L. New insight into metaphase arrest by cytostatic factor: From establishment to release. Oncogene 2007, 26, 1286–1289. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T.; Mitchison, T.J. Topoisomerase II does not play a scaffolding role in the organization of mitotic chromosomes assembled in Xenopus egg extracts. J. Cell Biol. 1993, 120, 601–612. [Google Scholar] [CrossRef]
- Shintomi, K.; Hirano, T. Guiding functions of the C-terminal domain of topoisomerase IIalpha advance mitotic chromosome assembly. Nat. Commun. 2021, 12, 2917. [Google Scholar] [CrossRef]
- Lewis, C.D.; Laemmli, U.K. Higher order metaphase chromosome structure: Evidence for metalloprotein interactions. Cell 1982, 29, 171–181. [Google Scholar] [CrossRef]
- Strick, R.; Strissel, P.L.; Gavrilov, K.; Levi-Setti, R. Cation-chromatin binding as shown by ion microscopy is essential for the structural integrity of chromosomes. J. Cell Biol. 2001, 155, 899–910. [Google Scholar] [CrossRef] [Green Version]
- Phengchat, R.; Takata, H.; Morii, K.; Inada, N.; Murakoshi, H.; Uchiyama, S.; Fukui, K. Calcium ions function as a booster of chromosome condensation. Sci. Rep. 2016, 6, 38281. [Google Scholar] [CrossRef] [Green Version]
- Maeshima, K.; Matsuda, T.; Shindo, Y.; Imamura, H.; Tamura, S.; Imai, R.; Kawakami, S.; Nagashima, R.; Soga, T.; Noji, H.; et al. A Transient Rise in Free Mg(2+) Ions Released from ATP-Mg Hydrolysis Contributes to Mitotic Chromosome Condensation. Curr. Biol. 2018, 28, 444–451.e446. [Google Scholar] [CrossRef]
- Hirano, T. Condensin-Based Chromosome Organization from Bacteria to Vertebrates. Cell 2016, 164, 847–857. [Google Scholar] [CrossRef] [Green Version]
- Uhlmann, F. SMC complexes: From DNA to chromosomes. Nat. Rev. Mol. Cell Biol. 2016, 17, 399–412. [Google Scholar] [CrossRef]
- Hassler, M.; Shaltiel, I.A.; Haering, C.H. Towards a Unified Model of SMC Complex Function. Curr. Biol. 2018, 28, R1266–R1281. [Google Scholar] [CrossRef] [Green Version]
- Wignall, S.M.; Deehan, R.; Maresca, T.J.; Heald, R. The condensin complex is required for proper spindle assembly and chromosome segregation in Xenopus egg extracts. J. Cell Biol. 2003, 161, 1041–1051. [Google Scholar] [CrossRef] [Green Version]
- Shintomi, K.; Hirano, T. The relative ratio of condensin I to II determines chromosome shapes. Genes Dev. 2011, 25, 1464–1469. [Google Scholar] [CrossRef] [Green Version]
- Green, L.C.; Kalitsis, P.; Chang, T.M.; Cipetic, M.; Kim, J.H.; Marshall, O.; Turnbull, L.; Whitchurch, C.B.; Vagnarelli, P.; Samejima, K.; et al. Contrasting roles of condensin I and condensin II in mitotic chromosome formation. J. Cell Sci. 2012, 125, 1591–1604. [Google Scholar] [CrossRef] [Green Version]
- Onn, I.; Aono, N.; Hirano, M.; Hirano, T. Reconstitution and subunit geometry of human condensin complexes. EMBO J. 2007, 26, 1024–1034. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, M.M.; Kinoshita, K.; Aizawa, Y.; Tane, S.; Yamashita, D.; Shintomi, K.; Hirano, T. Molecular dissection of condensin II-mediated chromosome assembly using in vitro assays. bioRxiv 2022. [Google Scholar] [CrossRef]
- Hara, K.; Kinoshita, K.; Migita, T.; Murakami, K.; Shimizu, K.; Takeuchi, K.; Hirano, T.; Hashimoto, H. Structural basis of HEAT-kleisin interactions in the human condensin I subcomplex. EMBO Rep. 2019, 20, e47183. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, K.; Tsubota, Y.; Tane, S.; Aizawa, Y.; Sakata, R.; Takeuchi, K.; Shintomi, K.; Nishiyama, T.; Hirano, T. A loop extrusion-independent mechanism contributes to condensin I-mediated chromosome shaping. J. Cell Biol. 2022, 221, e202109016. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Cuvier, O.; Hirano, T. Chromosome condensation by a human condensin complex in Xenopus egg extracts. J. Biol. Chem. 2001, 276, 5417–5420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemoto, A.; Kimura, K.; Yokoyama, S.; Hanaoka, F. Cell cycle-dependent phosphorylation, nuclear localization, and activation of human condensin. J. Biol. Chem. 2004, 279, 4551–4559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St-Pierre, J.; Douziech, M.; Bazile, F.; Pascariu, M.; Bonneil, E.; Sauvé, V.; Ratsima, H.; D’Amours, D. Polo kinase regulates mitotic chromosome condensation by hyperactivation of condensin DNA supercoiling activity. Mol. Cell 2009, 34, 416–426. [Google Scholar] [CrossRef]
- Kimura, K.; Hirano, T. ATP-dependent positive supercoiling of DNA by 13S condensin: A biochemical implication for chromosome condensation. Cell 1997, 90, 625–634. [Google Scholar] [CrossRef] [Green Version]
- Kimura, K.; Hirano, M.; Kobayashi, R.; Hirano, T. Phosphorylation and activation of 13S condensin by Cdc2 in vitro. Science 1998, 282, 487–490. [Google Scholar] [CrossRef]
- Ganji, M.; Shaltiel, I.A.; Bisht, S.; Kim, E.; Kalichava, A.; Haering, C.H.; Dekker, C. Real-time imaging of DNA loop extrusion by condensin. Science 2018, 360, 102–105. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Kerssemakers, J.; Shaltiel, I.A.; Haering, C.H.; Dekker, C. DNA-loop extruding condensin complexes can traverse one another. Nature 2020, 579, 438–442. [Google Scholar] [CrossRef]
- Kong, M.; Cutts, E.E.; Pan, D.; Beuron, F.; Kaliyappan, T.; Xue, C.; Morris, E.P.; Musacchio, A.; Vannini, A.; Greene, E.C. Human Condensin I and II Drive Extensive ATP-Dependent Compaction of Nucleosome-Bound DNA. Mol. Cell 2020, 79, 99–114.e119. [Google Scholar] [CrossRef]
- Ryu, J.K.; Rah, S.H.; Janissen, R.; Kerssemakers, J.W.J.; Bonato, A.; Michieletto, D.; Dekker, C. Condensin extrudes DNA loops in steps up to hundreds of base pairs that are generated by ATP binding events. Nucleic Acids Res. 2022, 50, 820–832. [Google Scholar] [CrossRef] [PubMed]
- Shaltiel, I.A.; Datta, S.; Lecomte, L.; Hassler, M.; Kschonsak, M.; Bravo, S.; Stober, C.; Ormanns, J.; Eustermann, S.; Haering, C.H. A hold-and-feed mechanism drives directional DNA loop extrusion by condensin. Science 2022, 376, 1087–1094. [Google Scholar] [CrossRef] [PubMed]
- Golfier, S.; Quail, T.; Kimura, H.; Brugués, J. Cohesin and condensin extrude DNA loops in a cell cycle-dependent manner. eLife 2020, 9, e53885. [Google Scholar] [CrossRef] [PubMed]
- Yatskevich, S.; Rhodes, J.; Nasmyth, K. Organization of Chromosomal DNA by SMC Complexes. Annu. Rev. Genet. 2019, 53, 445–482. [Google Scholar] [CrossRef] [PubMed]
- Davidson, I.F.; Peters, J.M. Genome folding through loop extrusion by SMC complexes. Nat. Rev. Mol. Cell Biol. 2021, 22, 445–464. [Google Scholar] [CrossRef]
- Higashi, T.L.; Uhlmann, F. SMC complexes: Lifting the lid on loop extrusion. Curr. Opin. Cell Biol. 2022, 74, 13–22. [Google Scholar] [CrossRef]
- Oldenkamp, R.; Rowland, B.D. A walk through the SMC cycle: From catching DNAs to shaping the genome. Mol. Cell 2022, 82, 1616–1630. [Google Scholar] [CrossRef]
- Wang, J.C. Cellular roles of DNA topoisomerases: A molecular perspective. Nat. Rev. Mol. Cell Biol. 2002, 3, 430–440. [Google Scholar] [CrossRef]
- Nitiss, J.L. DNA topoisomerase II and its growing repertoire of biological functions. Nat. Rev. Cancer 2009, 9, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Newport, J.; Spann, T. Disassembly of the nucleus in mitotic extracts: Membrane vesicularization, lamin disassembly, and chromosome condensation are independent processes. Cell 1987, 48, 219–230. [Google Scholar] [CrossRef]
- Adachi, Y.; Luke, M.; Laemmli, U.K. Chromosome assembly in vitro: Topoisomerase II is required for condensation. Cell 1991, 64, 137–148. [Google Scholar] [CrossRef]
- Linka, R.M.; Porter, A.C.; Volkov, A.; Mielke, C.; Boege, F.; Christensen, M.O. C-terminal regions of topoisomerase IIalpha and IIbeta determine isoform-specific functioning of the enzymes in vivo. Nucleic Acids Res. 2007, 35, 3810–3822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohta, S.; Bukowski-Wills, J.C.; Sanchez-Pulido, L.; Alves Fde, L.; Wood, L.; Chen, Z.A.; Platani, M.; Fischer, L.; Hudson, D.F.; Ponting, C.P.; et al. The protein composition of mitotic chromosomes determined using multiclassifier combinatorial proteomics. Cell 2010, 142, 810–821. [Google Scholar] [CrossRef] [Green Version]
- Booth, D.G.; Beckett, A.J.; Molina, O.; Samejima, I.; Masumoto, H.; Kouprina, N.; Larionov, V.; Prior, I.A.; Earnshaw, W.C. 3D-CLEM Reveals that a Major Portion of Mitotic Chromosomes Is Not Chromatin. Mol. Cell 2016, 64, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Shechter, D.; Chitta, R.K.; Xiao, A.; Shabanowitz, J.; Hunt, D.F.; Allis, C.D. A distinct H2A.X isoform is enriched in Xenopus laevis eggs and early embryos and is phosphorylated in the absence of a checkpoint. Proc. Natl. Acad. Sci. USA 2009, 106, 749–754. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.; Liu, Y.; Luger, K. Histone chaperone FACT FAcilitates Chromatin Transcription: Mechanistic and structural insights. Curr. Opin. Struct. Biol. 2020, 65, 26–32. [Google Scholar] [CrossRef]
- Haase, J.; Chen, R.; Parker, W.M.; Bonner, M.K.; Jenkins, L.M.; Kelly, A.E. The TFIIH complex is required to establish and maintain mitotic chromosome structure. eLife 2022, 11, e75475. [Google Scholar] [CrossRef] [PubMed]
- Brykczynska, U.; Hisano, M.; Erkek, S.; Ramos, L.; Oakeley, E.J.; Roloff, T.C.; Beisel, C.; Schübeler, D.; Stadler, M.B.; Peters, A.H. Repressive and active histone methylation mark distinct promoters in human and mouse spermatozoa. Nat. Struct. Mol. Biol. 2010, 17, 679–687. [Google Scholar] [CrossRef]
- Ray-Gallet, D.; Quivy, J.P.; Sillje, H.W.; Nigg, E.A.; Almouzni, G. The histone chaperone Asf1 is dispensable for direct de novo histone deposition in Xenopus egg extracts. Chromosoma 2007, 116, 487–496. [Google Scholar] [CrossRef]
- Bradbury, E.M.; Inglis, R.J.; Matthews, H.R. Control of cell division by very lysine rich histone (F1) phosphorylation. Nature 1974, 247, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Mueller, R.D.; Yasuda, H.; Bradbury, E.M. Phosphorylation of histone H1 through the cell cycle of Physarum polycephalum. 24 sites of phosphorylation at metaphase. J. Biol. Chem. 1985, 260, 5081–5086. [Google Scholar] [CrossRef]
- Langan, T.A.; Gautier, J.; Lohka, M.; Hollingsworth, R.; Moreno, S.; Nurse, P.; Maller, J.; Sclafani, R.A. Mammalian growth-associated H1 histone kinase: A homolog of cdc2+/CDC28 protein kinases controlling mitotic entry in yeast and frog cells. Mol. Cell. Biol. 1989, 9, 3860–3868. [Google Scholar] [PubMed] [Green Version]
- Smith, R.C.; Dworkin-Rastl, E.; Dworkin, M.B. Expression of a histone H1-like protein is restricted to early Xenopus development. Genes Dev. 1988, 2, 1284–1295. [Google Scholar] [CrossRef] [Green Version]
- Ohsumi, K.; Katagiri, C.; Kishimoto, T. Chromosome condensation in Xenopus mitotic extracts without histone H1. Science 1993, 262, 2033–2035. [Google Scholar] [CrossRef] [PubMed]
- Maresca, T.J.; Freedman, B.S.; Heald, R. Histone H1 is essential for mitotic chromosome architecture and segregation in Xenopus laevis egg extracts. J. Cell Biol. 2005, 169, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Choppakatla, P.; Dekker, B.; Cutts, E.E.; Vannini, A.; Dekker, J.; Funabiki, H. Linker histone H1.8 inhibits chromatin binding of condensins and DNA topoisomerase II to tune chromosome length and individualization. eLife 2021, 10, e68918. [Google Scholar] [CrossRef]
- Naumova, N.; Imakaev, M.; Fudenberg, G.; Zhan, Y.; Lajoie, B.R.; Mirny, L.A.; Dekker, J. Organization of the mitotic chromosome. Science 2013, 342, 948–953. [Google Scholar] [CrossRef] [Green Version]
- Kakui, Y.; Rabinowitz, A.; Barry, D.J.; Uhlmann, F. Condensin-mediated remodeling of the mitotic chromatin landscape in fission yeast. Nat. Genet. 2017, 49, 1553–1557. [Google Scholar] [CrossRef] [Green Version]
- Lazar-Stefanita, L.; Scolari, V.F.; Mercy, G.; Muller, H.; Guérin, T.M.; Thierry, A.; Mozziconacci, J.; Koszul, R. Cohesins and condensins orchestrate the 4D dynamics of yeast chromosomes during the cell cycle. EMBO J. 2017, 36, 2684–2697. [Google Scholar] [CrossRef] [PubMed]
- Schalbetter, S.A.; Goloborodko, A.; Fudenberg, G.; Belton, J.M.; Miles, C.; Yu, M.; Dekker, J.; Mirny, L.; Baxter, J. SMC complexes differentially compact mitotic chromosomes according to genomic context. Nat. Cell Biol. 2017, 19, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Gibcus, J.H.; Samejima, K.; Goloborodko, A.; Samejima, I.; Naumova, N.; Nuebler, J.; Kanemaki, M.T.; Xie, L.; Paulson, J.R.; Earnshaw, W.C.; et al. A pathway for mitotic chromosome formation. Science 2018, 359, eaao6135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almagro, S.; Riveline, D.; Hirano, T.; Houchmandzadeh, B.; Dimitrov, S. The mitotic chromosome is an assembly of rigid elastic axes organized by structural maintenance of chromosomes (SMC) proteins and surrounded by a soft chromatin envelope. J. Biol. Chem. 2004, 279, 5118–5126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Maresca, T.J.; Skoko, D.; Adams, C.D.; Xiao, B.; Christensen, M.O.; Heald, R.; Marko, J.F. Micromanipulation studies of chromatin fibers in Xenopus egg extracts reveal ATP-dependent chromatin assembly dynamics. Mol. Biol. Cell 2007, 18, 464–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, B.; Freedman, B.S.; Miller, K.E.; Heald, R.; Marko, J.F. Histone H1 compacts DNA under force and during chromatin assembly. Mol. Biol. Cell 2012, 23, 4864–4871. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Biggs, R.; Hornick, J.; Marko, J.F. Condensin controls mitotic chromosome stiffness and stability without forming a structurally contiguous scaffold. Chromosome Res. 2018, 26, 277–295. [Google Scholar] [CrossRef] [PubMed]
- Meijering, A.E.C.; Sarlós, K.; Nielsen, C.F.; Witt, H.; Harju, J.; Kerklingh, E.; Haasnoot, G.H.; Bizard, A.H.; Heller, I.; Broedersz, C.P.; et al. Nonlinear mechanics of human mitotic chromosomes. Nature 2022, 605, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Wakayama, T.; Perry, A.C.; Zuccotti, M.; Johnson, K.R.; Yanagimachi, R. Full-term development of mice from enucleated oocytes injected with cumulus cell nuclei. Nature 1998, 394, 369–374. [Google Scholar] [CrossRef]
- Egli, D.; Rosains, J.; Birkhoff, G.; Eggan, K. Developmental reprogramming after chromosome transfer into mitotic mouse zygotes. Nature 2007, 447, 679–685. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shintomi, K. Making Mitotic Chromosomes in a Test Tube. Epigenomes 2022, 6, 20. https://doi.org/10.3390/epigenomes6030020
Shintomi K. Making Mitotic Chromosomes in a Test Tube. Epigenomes. 2022; 6(3):20. https://doi.org/10.3390/epigenomes6030020
Chicago/Turabian StyleShintomi, Keishi. 2022. "Making Mitotic Chromosomes in a Test Tube" Epigenomes 6, no. 3: 20. https://doi.org/10.3390/epigenomes6030020
APA StyleShintomi, K. (2022). Making Mitotic Chromosomes in a Test Tube. Epigenomes, 6(3), 20. https://doi.org/10.3390/epigenomes6030020