
Genetic Factors That Could Affect Concussion Risk in Elite Rugby

 , ,
, ,  ,
, 
 and
and 
Abstract
:1. Introduction
2. Incidence Rate and Severity of Concussion in Rugby
3. Mechanisms of Concussion
4. Pathophysiology of Concussion
5. Genetic Associations with Concussion
5.1. Candidate Genetic Variants
5.1.1. Apolipoprotein E
5.1.2. Microtubule-Associated Protein Tau Polymorphisms
5.1.3. Neurofilament Heavy Polymorphism
5.1.4. Membrane Metalloendopeptidase Polymorphism
5.1.5. Brain-Derived Neurotrophic Factor Polymorphism
5.1.6. Glutamate Ionotropic Receptor NMDA Type Subunit 2A Variant
5.1.7. Catechol-O-methyltransferase Polymorphism
5.1.8. Ankyrin Repeat and Kinase Domain Containing 1 Polymorphism
5.1.9. Dopamine Receptor-Related Polymorphisms
5.1.10. Serotonin Transporter Polymorphisms
5.1.11. Endothelial Nitric Oxide Synthase Polymorphism
5.1.12. Angiotensin I-Converting Enzyme Variants
5.1.13. Tumour Necrosis Factor Polymorphisms
5.1.14. Transforming Growth Factor Beta1 Polymorphism
5.1.15. Interleukin 1 Alpha and Interleukin 1 Beta Polymorphisms
5.1.16. Interleukin 6 Receptor Polymorphism
6. Conclusions and Future Directions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tierney, R.; Mansell, J.; Higgins, M.; McDevitt, J.; Toone, N.; Gaughan, J.; Mishra, A.; Krynetskiy, E. Apolipoprotein E Genotype and Concussion in College Athletes. Clin. J. Sport Med. 2010, 20, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Terrell, T.; Bostick, R.; Abramson, R.; Xie, D.; Barfield, W.; Cantu, R.; Stanek, M.; Ewing, T. APOE, APOE Promoter, and Tau Genotypes and Risk for Concussion in College Athletes. Clin. J. Sport Med. 2008, 18, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Terrell, T.; Bostick, R.; Barth, J.; McKeag, D.; Cantu, R.; Sloane, R.; Galloway, L.; Erlanger, D.; Valentine, V.; Bielak, K. Genetic Polymorphisms, Concussion Risk, and Post Concussion Neurocognitive Deficits in College and High School Athletes. Br. J. Sports Med. 2013, 47, e1. [Google Scholar] [CrossRef]
- Cross, M.; Kemp, S.; Smith, A.; Trewartha, G.; Stokes, K. Professional Rugby Union Players Have a 60% Greater Risk of Time Loss Injury after Concussion: A 2-Season Prospective Study of Clinical Outcomes. Br. J. Sports Med. 2016, 50, 926–931. [Google Scholar] [CrossRef] [Green Version]
- McCrory, P.; Meeuwisse, W.; Dvorak, J.; Aubry, M.; Bailes, J.; Broglio, S.; Cantu, B.; Cassidy, D.; Echemendia, R.J.; Castellani, R.J.; et al. Consensus Statement on Concussion in Sport—the 5th International Conference on Concussion in Sport Held in Berlin, October 2016. Br. J. Sports Med. 2018, 51, 838–847. [Google Scholar]
- Cunningham, J.; Broglio, S.; Wilson, F. Influence of Playing Rugby on Long-Term Brain Health Following Retirement: A Systematic Review and Narrative Synthesis. BMJ Open Sport Exerc. Med. 2018, 4, e000356. [Google Scholar] [CrossRef]
- Blennow, K.; de Leon, M.; Zetterberg, H. Alzheimer’s Disease. Lancet 2006, 368, 387–403. [Google Scholar] [CrossRef]
- Hume, P.; Theadom, A.; Lewis, G.; Quarrie, K.; Brown, S.; Hill, R.; Marshall, S. A Comparison of Cognitive Function in Former Rugby Union Players Compared with Former Non-Contact-Sport Players and the Impact of Concussion History. Sports Med. 2017, 47, 1209–1220. [Google Scholar] [CrossRef]
- Quintana, L.M. Second Impact Syndrome in Sports. World Neurosurg. 2016, 91, 647–649. [Google Scholar] [CrossRef]
- Broglio, S.P.; Eckner, J.T.; Paulson, H.L.; Kutcher, J. Cognitive Decline and Aging. Exerc. Sport Sci. Rev. 2012, 40, 138–144. [Google Scholar] [CrossRef]
- Lee, Y.-K.; Hou, S.-W.; Lee, C.-C.; Hsu, C.-Y.; Huang, Y.-S.; Su, Y.-C. Increased Risk of Dementia in Patients with Mild Traumatic Brain Injury: A Nationwide Cohort Study. PLoS ONE 2013, 8, e62422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerr, Z.Y.; Evenson, K.R.; Rosamond, W.D.; Mihalik, J.P.; Guskiewicz, K.M.; Marshall, S.W. Association between Concussion and Mental Health in Former Collegiate Athletes. Inj. Epidemiol. 2014, 1, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guskiewicz, K.M.; Marshall, S.W.; Bailes, J.; McCrea, M.; Harding, H.P.; Matthews, A.; Mihalik, J.R.; Cantu, R.C. Recurrent Concussion and Risk of Depression in Retired Professional Football Players. Med. Sci. Sport. Exerc. 2007, 39, 903–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stulemeijer, M.; Andriessen, T.M.; Brauer, J.M.P.; Vos, P.E.; Van Der Werf, S. Cognitive Performance after Mild Traumatic Brain Injury: The Impact of Poor Effort on Test Results and Its Relation to Distress, Personality and Litigation. Brain Inj. 2007, 21, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Raikes, A.C.; Athey, A.; Alfonso-Miller, P.; Killgore, W.D.S.; Grandner, M.A. Insomnia and Daytime Sleepiness: Risk Factors for Sports-Related Concussion. Sleep Med. 2019, 58, 66–74. [Google Scholar] [CrossRef] [Green Version]
- Lust, C.A.C.; Mountjoy, M.; Robinson, L.E.; Oliver, J.M.; Ma, D.W.L. Sports-Related Concussions and Subconcussive Impacts in Athletes: Incidence, Diagnosis, and the Emerging Role of EPA and DHA. Appl. Physiol. Nutr. Metab. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrahams, S.; Mc Fie, S.; Patricios, J.; Posthumus, M.; September, A.V. Risk Factors for Sports Concussion: An Evidence-Based Systematic Review. Br. J. Sports Med. 2013. [Google Scholar] [CrossRef]
- Gardner, A.J.; Iverson, G.L.; Williams, W.H.; Baker, S.; Stanwell, P. A Systematic Review and Meta-Analysis of Concussion in Rugby Union. Sports Med. 2014, 44, 1717–1731. [Google Scholar] [CrossRef]
- Gardner, A.; Iverson, G.L.; Levi, C.R.; Schofield, P.W.; Kay-Lambkin, F.; Kohler, R.M.N.; Stanwell, P. A Systematic Review of Concussion in Rugby League. Br. J. Sports Med. 2015, 49, 495–498. [Google Scholar] [CrossRef] [Green Version]
- Abrahams, S.; Mc Fie, S.; Patricios, J.; Suter, J.; September, A.V.; Posthumus, M. Toxic Tau: The TAU Gene Polymorphisms Associate with Concussion History in Rugby Union Players. J. Sci. Med. Sport 2019, 22, 22–28. [Google Scholar] [CrossRef]
- Mc Fie, S.; Abrahams, S.; Patricios, J.; Suter, J.; Posthumus, M.; September, A.V. The Association between COMT Rs4680 and 5-HTTLPR Genotypes and Concussion History in South African Rugby Union Players. J. Sports Sci. 2018, 36, 920–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terrell, T.R.; Abramson, R.; Barth, J.T.; Bennett, E.; Cantu, R.C.; Sloane, R.; Laskowitz, D.T.; Erlanger, D.M.; McKeag, D.; Nichols, G.; et al. Genetic Polymorphisms Associated with the Risk of Concussion in 1056 College Athletes: A Multicentre Prospective Cohort Study. Br. J. Sports Med. 2018, 52, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Davidson, J.; Cusimano, M.D.; Bendena, W.G. Post-Traumatic Brain Injury. Neuroscience 2015, 21, 424–441. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.M.; Portelli, J.N.; Chau, A.; Cristofori, I.; Moretti, L.; Grafman, J. Genetic Polymorphisms and Traumatic Brain Injury: The Contribution of Individual Differences to Recovery. Brain Imaging Behav. 2014, 8, 420–434. [Google Scholar] [CrossRef]
- McAllister, T.W. Genetic Factors Modulating Outcome after Neurotrauma. PM&R 2010, 2 (Suppl. 2), S241–S252. [Google Scholar] [CrossRef]
- Wilson, M.; Montgomery, H. Impact of Genetic Factors on Outcome from Brain Injury. Br. J. Anaesth. 2007, 99, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Mayhew, A.J.; Meyre, D. Assessing the Heritability of Complex Traits in Humans: Methodological Challenges and Opportunities. Curr. Genom. 2017, 18, 332. [Google Scholar] [CrossRef] [Green Version]
- Kazl, C.; Torres, A. Definition, Classification, and Epidemiology of Concussion. Semin. Pediatr. Neurol. 2019, 30, 9–13. [Google Scholar] [CrossRef]
- Hakim, A.J.; Cherkas, L.F.; Spector, T.D.; MacGregor, A.J. Genetic Associations between Frozen Shoulder and Tennis Elbow: A Female Twin Study. Rheumatology 2003, 42, 739–742. [Google Scholar]
- Ralston, S.H.; Uitterlinden, A.G. Genetics of Osteoporosis. Endocr. Rev. 2010, 31, 629–662. [Google Scholar] [CrossRef] [Green Version]
- Carmelli, D.; DeCarli, C.; Swan, G.E.; Jack, L.M.; Reed, T.; Wolf, P.A.; Miller, B.L. Evidence for Genetic Variance in White Matter Hyperintensity Volume in Normal Elderly Male Twins. Stroke 1998, 29, 1177–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geschwind, D.H.; Miller, B.L.; DeCarli, C.; Carmelli, D. Heritability of Lobar Brain Volumes in Twins Supports Genetic Models of Cerebral Laterality and Handedness. Proc. Natl. Acad. Sci. USA 2002, 99, 3176–3181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmelli, D.; Swan, G.E.; DeCarli, C.; Reed, T. Quantitative Genetic Modeling of Regional Brain Volumes and Cognitive Performance in Older Male Twins. Biol. Psychol. 2002, 61, 139–155. [Google Scholar] [CrossRef]
- Bartley, A.J.; Jones, D.W.; Weinberger, D.R. Genetic Variability of Human Brain Size and Cortical Gyral Patterns. Brain 1997, 120, 257–269. [Google Scholar] [CrossRef]
- McKee, A.C.; Daneshvar, D.H.; Alvarez, V.E.; Stein, T.D. The Neuropathology of Sport. Acta Neuropathol. 2014, 127, 29–51. [Google Scholar] [CrossRef]
- Dashnaw, M.L.; Petraglia, A.L.; Bailes, J.E. An Overview of the Basic Science of Concussion and Subconcussion: Where We Are and Where We Are Going. Neurosurg. Focus 2012, 33, E5. [Google Scholar] [CrossRef] [Green Version]
- Sedeaud, A.; Marc, A.; Schipman, J.; Tafflet, M.; Hager, J.-P.; Toussaint, J.-F. How They Won Rugby World Cup through Height, Mass and Collective Experience. Br. J. Sports Med. 2012, 46, 580–584. [Google Scholar] [CrossRef]
- Austin, D.; Gabbett, T.; Jenkins, D. The Physical Demands of Super 14 Rugby Union. J. Sci. Med. Sport 2011, 14, 259–263. [Google Scholar] [CrossRef]
- Duthie, G.; Pyne, D.; Hooper, S. Applied Physiology and Game Analysis of Rugby Union. Sports Med. 2003, 33, 973–991. [Google Scholar] [CrossRef]
- Brazier, J.; Antrobus, M.; Stebbings, G.K.; Day, S.H.; Callus, P.; Erskine, R.M.; Bennett, M.A.; Kilduff, L.P.; Williams, A.G. Anthropometric and Physiological Characteristics of Elite Male Rugby Athletes. J. Strength Cond. Res. 2020, 34, 1790–1801. [Google Scholar] [CrossRef]
- Hill, N.; Rilstone, S.; Stacey, M.; Amiras, D.; Chew, S.; Flatman, D.; Oliver, N. Changes in Northern Hemisphere Male International Rugby Union Players’ Body Mass and Height between 1955 and 2015. BMJ Open Sport Exerc. Med. 2018, 4, e000459. [Google Scholar] [CrossRef] [PubMed]
- Johnston, R.D.; Gabbett, T.J.; Jenkins, D.G. Applied Sport Science of Rugby League. Sports Med. 2014, 44, 1087–1100. [Google Scholar] [CrossRef] [PubMed]
- Quarrie, K.L.; Hopkins, W.G. Changes in Player Characteristics and Match Activities in Bledisloe Cup Rugby Union from 1972 to 2004. J. Sports Sci. 2007, 25, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Eaves, S.; Hughes, M. Patterns of Play of International Rugby Union Teams before and after the Introduction of Professional Status. Int. J. Perform. Anal. Sport 2003, 3, 103–111. [Google Scholar] [CrossRef]
- Williams, S.; Trewartha, G.; Kemp, S.; Stokes, K. A Meta-Analysis of Injuries in Senior Men’s Professional Rugby Union. Sports Med. 2013, 43, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- King, D.A.; Hume, P.A.; Milburn, P.D.; Guttenbeil, D. Match and Training Injuries in Rugby League. Sports Med. 2010, 40, 163–178. [Google Scholar] [CrossRef]
- Fitzpatrick, A.; Naylor, A.; Myler, P.; Robertson, C. A Three-Year Epidemiological Prospective Cohort Study of Rugby League Match Injuries from the European Super League. J. Sci. Med. Sport 2018, 21, 160–165. [Google Scholar] [CrossRef]
- England Professional Rugby Injury Surveillance Project Steering Group. England Professional Rugby Injury Surveillance Project 2017–2018 Season Report; England Professional Rugby Injury Surveillance Project Steering Group: London, UK, 2019; p. 15. [Google Scholar]
- Clay, M.B.; Glover, K.L.; Lowe, D.T. Epidemiology of Concussion in Sport: A Literature Review. J. Chiropr. Med. 2013, 12, 230–251. [Google Scholar] [CrossRef] [Green Version]
- Koh, J.O.; Cassidy, J.D.; Watkinson, E.J. Incidence of Concussion in Contact Sports: A Systematic Review of the Evidence. Brain Inj. 2003, 17, 901–917. [Google Scholar] [CrossRef]
- Chachad, S.; Khan, A. Concussion in the Athlete: A Review. Clin. Pediatr. 2006, 285–288. [Google Scholar] [CrossRef]
- Tucker, R.; Raftery, M.; Fuller, G.; Hester, B.; Kemp, S.; Cross, M. A Video Analysis of Head Injuries Satisfying the Criteria for a Head Injury Assessment in Professional Rugby Union: A Prospective Cohort Study. Br. J. Sports Med. 2017, 51, 1147–1151. [Google Scholar] [CrossRef] [PubMed]
- Gabbett, T.J.; Jenkins, D.G.; Abernethy, B. Physical Collisions and Injury in Professional Rugby League Match-Play. J. Sci. Med. Sport 2011, 14, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Fuller, C.W.; Ashton, T.; Brooks, J.H.M.; Cancea, R.J.; Hall, J.; Kemp, S.P.T. Injury Risks Associated with Tackling in Rugby Union. Br. J. Sports Med. 2010, 44, 159–167. [Google Scholar] [CrossRef] [PubMed]
- McCrory, P.; Meeuwisse, W.H.; Aubry, M.; Cantu, R.C.; Dvořák, J.; Echemendia, R.J.; Engebretsen, L.; Johnston, K.; Kutcher, J.S.; Raftery, M.; et al. Consensus Statement on Concussion in Sport: The 4th International Conference on Concussion in Sport, Zurich, November 2012. J. Athl. Train. 2013, 48, 554–575. [Google Scholar] [CrossRef] [Green Version]
- Iverson, G.L.; Brooks, B.L.; Collins, M.W.; Lovell, M.R. Tracking Neuropsychological Recovery Following Concussion in Sport. Brain Inj. 2006, 20, 245–252. [Google Scholar] [CrossRef]
- Pellman, E.J.; Viano, D.C.; Casson, I.R.; Arfken, C.; Powell, J. Concussion in Professional Football: Injuries Involving 7 or More Days out—Part 5. Neurosurgery 2004, 55, 1100–1116. [Google Scholar] [CrossRef]
- Fuller, C.W.; Taylor, A.; Kemp, S.P.T.; Raftery, M. Rugby World Cup 2015: World Rugby Injury Surveillance Study. Br. J. Sports Med. 2017, 51, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Fuller, C.W.; Taylor, A.; Douglas, M.; Rafter, M.; Taylor, A.; Douglas, M.; Raftery, M. Rugby World Cup 2019 Injury Surveillance Study. S. Afr. J. Sports Med. 2020, 32, 1–6. [Google Scholar] [CrossRef]
- Rafferty, J.; Ranson, C.; Oatley, G.; Mostafa, M.; Mathema, P.; Crick, T.; Moore, I.S. On Average, a Professional Rugby Union Player Is More Likely than Not to Sustain a Concussion after 25 Matches. Br. J. Sports Med. 2019, 53, 969–973. [Google Scholar] [CrossRef] [Green Version]
- Video Review and Validation Study. Sports Med. Open 2017, 3, 29. [CrossRef] [Green Version]
- Gardner, A.J.; Howell, D.R.; Iverson, G.L. National Rugby League Match Scheduling and Rate of Concussion. J. Sci. Med. Sport 2019, 22, 780–783. [Google Scholar] [CrossRef] [PubMed]
- Savage, J.; Hooke, C.; Orchard, J.; Parkinson, R. The Incidence of Concussion in a Professional Australian Rugby League Team, 1998–2012. J. Sports Med. 2013, 2013, 304576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosgrave, M.; Williams, S. The Epidemiology of Concussion in Professional Rugby Union in Ireland. Phys. Ther. Sport 2019, 35, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Hinton-Bayre, A.D.; Geffen, G.; Friis, P. Presentation and Mechanisms of Concussion in Professional Rugby League Football. J. Sci. Med. Sport 2004, 7, 400–404. [Google Scholar] [CrossRef]
- Gabbett, T.J. Influence of Training and Match Intensity on Injuries in Rugby League. J. Sports Sci. 2004, 22, 409–417. [Google Scholar] [CrossRef]
- Kemp, S.; West, S.; Brooks, J.; Cross, M.; Williams, S.; Anstiss, T.; Smith, A.; Bryan, R.; Hibbins-Butler, R.; O’Leary, B.; et al. The Professional Rugby Injury Surveillance Project (PRISP) Annual Report(s) 2016/17; Rugby Football Union: Twickenham, UK, 2018. [Google Scholar]
- Bigler, E. Anterior and Middle Cranial Fossa in Traumatic Brain Injury: Relevant Neuroanatomy and Neuropathology in the Study of Neuropsychological Outcome. Neuropsychology 2007, 21, 515–531. [Google Scholar] [CrossRef] [Green Version]
- Meaney, D.F.; Smith, D.H. Biomechanics of Concussion. Clin. Sports Med. 2011, 30, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Post, A.; Hoshizaki, B.T. Rotational Acceleration, Brain Tissue Strain, and the Relationship to Concussion. J. Biomech. Eng. 2015, 137. [Google Scholar] [CrossRef]
- Hoshizaki, B.; Post, A.; Kendall, M.; Karton, C.; Brien, S. The Relationship between Head Impact Characteristics and Brain Trauma. J. Neurol. Neurophysiol. 2013, 5, 1–8. [Google Scholar] [CrossRef]
- McAllister, T.W. Neurobiological Consequences of Traumatic Brain Injury. Dialogues Clin. Neurosci. 2011, 13, 287–300. [Google Scholar]
- Meythaler, J.M.; Peduzzi, J.D.; Eleftheriou, E.; Novack, T.A. Current Concepts: Diffuse Axonal Injury–Associated Traumatic Brain Injury. Arch. Phys. Med. Rehabil. 2001, 82, 1461–1471. [Google Scholar] [CrossRef]
- Wu, L.C.; Nangia, V.; Bui, K.; Hammoor, B.; Kurt, M.; Hernandez, F.; Kuo, C.; Camarillo, D.B. In Vivo Evaluation of Wearable Head Impact Sensors. Ann. Biomed. Eng. 2016, 44, 1234. [Google Scholar] [CrossRef] [Green Version]
- Guskiewicz, K.M.; Mihalik, J.P.; Shankar, V.; Marshall, S.W.; Crowell, D.H.; Oliaro, S.M.; Ciocca, M.F.; Hooker, D.N. Measurement of head impacts in collegiate football playerS. Neurosurgery 2007, 61, 1244–1253. [Google Scholar] [CrossRef]
- Giza, C.C.; Hovda, D.A. The New Neurometabolic Cascade of Concussion. Neurosurgery 2014, 75, S24–S33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giza, C.C.; Hovda, D.A. The Neurometabolic Cascade of Concussion. J. Athl. Train. 2001, 36, 228–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazarian, J.; Zhong, J.; Blyth, B.; Zhu, T.; Kavcic, V.; Peterson, D. Diffusion Tensor Imaging Detects Clinically Important Axonal Damage after Mild Traumatic Brain Injury: A Pilot Study. J. Neurotrauma 2007, 24, 1447–1459. [Google Scholar] [CrossRef] [PubMed]
- Blumbergs, P.; Scott, G.; Manavis, J.; Wainwright, H.; Simpson, D.; McLean, A. Staining of Amyloid Precursor Protein to Study Axonal Damage in Mild Head Injury. Lancet 1994, 344, 1055–1056. [Google Scholar] [CrossRef]
- Blumbergs, P.; Scott, G.; Vis, J.; Wainwright, H.; Simpson, D.; McLean, A.J. Topography of Axonal Injury as Defined by Amyloid Precursor Protein and the Sector Scoring Method in Mild and Severe Closed Head Injury. J. Neurotrauma 1995, 12, 565–572. [Google Scholar] [CrossRef]
- Povlishock, J.T.; Pettus, E.H. Traumatically Induced Axonal Damage: Evidence for Enduring Changes in Axolemmal Permeability with Associated Cytoskeletal Change. Acta Neurochir. Suppl. 1996, 1996, 81–86. [Google Scholar] [CrossRef]
- Katayama, Y.; Becker, D.P.; Tamura, T.; Hovda, D.A. Massive Increases in Extracellular Potassium and the Indiscriminate Release of Glutamate Following Concussive Brain Injury. J. Neurosurg. 1990, 73, 889–900. [Google Scholar] [CrossRef]
- Cantu, R.; Cantu, R. Neurologic Athletic Head and Spine Injuries; W.B. Saunders Co.: Philadelphia, PA, USA, 2000. [Google Scholar]
- Büki, A.; Povlishock, J. All Roads Lead to Disconnection?—Traumatic Axonal Injury Revisited. Acta Neurochir. 2006, 148, 181–194. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, R.; Maris, D.O.; Grady, M.S.; Winn, H.R.; Janigro, D. Impaired K+ Homeostasis and Altered Electrophysiological Properties of Post-Traumatic Hippocampal Gila. J. Neurosci. 1999, 19, 8152–8162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartings, J.A.; Strong, A.J.; Fabricius, M.; Manning, A.; Bhatia, R.; Dreier, J.P.; Mazzeo, A.T.; Tortella, F.C.; Bullock, M.R. Spreading Depolarizations and Late Secondary Insults after Traumatic Brain Injury. J. Neurotrauma 2009, 26, 1857–1866. [Google Scholar] [CrossRef] [PubMed]
- Gaetz, M. The Neurophysiology of Brain Injury. Clin. Neurophysiol. 2004, 115, 4–18. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.W. Ionic Dependence of Glutamate Neurotoxicity. J. Neurosci. 1987, 7, 369–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Nelson, L.D.; Laroche, A.A.; Pfaller, A.Y.; Nencka, A.S.; Koch, K.M.; McCrea, M.A. Cerebral Blood Flow Alterations in Acute Sport-Related Concussion. J. Neurotrauma 2016, 33, 1227–1236. [Google Scholar] [CrossRef] [Green Version]
- Weber, J.T. Altered Calcium Signaling Following Traumatic Brain Injury. Front. Pharmacol. 2012, 3, 60. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Peterson, P.L.; Verweij, B.H.; Vinas, F.C.; Muizelaar, J.P.; LEE, C.P. Mitochondrial Dysfunction After Experimental Traumatic Brain Injury: Combined Efficacy of SNX-111 and U-101033E. J. Neurotrauma 1998, 15, 531–544. [Google Scholar] [CrossRef]
- Patterson, Z.R.; Holahan, M.R. Understanding the Neuroinflammatory Response Following Concussion to Develop Treatment Strategies. Front. Cell. Neurosci. 2012, 6. [Google Scholar] [CrossRef] [Green Version]
- Zaitlen, N.; Kraft, P. Heritability in the Genome-Wide Association Era. Hum. Genet. 2012, 1655–1664. [Google Scholar] [CrossRef] [Green Version]
- Zuk, O.; Hechter, E.; Sunyaev, S.R.; Lander, E.S. The Mystery of Missing Heritability: Genetic Interactions Create Phantom Heritability. Proc. Natl. Acad. Sci. USA 2012, 109, 1193–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabor, H.K.; Risch, N.J.; Myers, R.M. Candidate-Gene Approaches for Studying Complex Genetic Traits: Practical Considerations. Nat. Rev. Genet. 2002, 3, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Rankinen, T.; Bray, M.S.; Hagberg, J.M.; Pérusse, L.; Roth, S.M.; Wolfarth, B.; Bouchard, C. The Human Gene Map for Performance and Health-Related Fitness Phenotypes: The 2005 Update. Med. Sci. Sports Exerc. 2006, 38, 1863–1888. [Google Scholar] [CrossRef] [PubMed]
- Bellosta, S.; Nathan, B.; Orth, M.; Dong, L.; Mahley, R.; Pitas, R. Stable Expression and Secretion of Apolipoproteins E3 and E4 in Mouse Neuroblastoma Cells Produces Differential Effects on Neurite Outgrowth. J. Biol. Chem. 1995, 270, 27063–27071. [Google Scholar] [CrossRef] [Green Version]
- Holtzman, D.M.; Pitas, R.E.; Kilbridge, J.; Nathan, B.; Mahley, R.W.; Bu, G.; Schwartz, A.L. Low Density Lipoprotein Receptor-Related Protein Mediates Apolipoprotein E-Dependent Neurite Outgrowth in a Central Nervous System-Derived Neuronal Cell Line. Proc. Natl. Acad. Sci. USA 1995, 92, 9480–9484. [Google Scholar] [CrossRef] [Green Version]
- Namboori, P.K.K.; Vineeth, K.V.; Rohith, V.; Hassan, I.; Sekhar, L.; Sekhar, A.; Nidheesh, M. The ApoE Gene of Alzheimer’s Disease (AD). Funct. Integr. Genom. 2011, 519–522. [Google Scholar] [CrossRef]
- Kristman, V.L.; Tator, C.H.; Kreiger, N.; Richards, D.; Mainwaring, L.; Jaglal, S.; Tomlinson, G.; Comper, P. Does the Apolipoprotein Ε4 Allele Predispose Varsity Athletes to Concussion? A Prospective Cohort Study. Clin. J. Sport Med. 2008, 18, 322–328. [Google Scholar] [CrossRef]
- Lambert, J.-C.; Araria-Goumidi, L.; Myllykangas, L.; Ellis, C.; Wang, J.C.; Bullido, M.J.; Harris, J.M.; Artiga, M.J.; Hernandez, D.; Kwon, J.M.; et al. Contribution of APOE Promoter Polymorphisms to Alzheimer’s Disease Risk. Neurology 2002, 59, 59–66. [Google Scholar] [CrossRef]
- Lendon, C.L.; Harris, J.M.; Pritchard, A.L.; Nicoll, J.A.R.; Teasdale, G.M.; Murray, G. Genetic Variation of the APOE Promoter and Outcome after Head Injury. Neurology 2003, 61, 683–685. [Google Scholar] [CrossRef]
- Gabbita, S.P.; Scheff, S.W.; Menard, R.M.; Roberts, K.; Fugaccia, I.; Zemlan, F.P. Cleaved-Tau: A Biomarker of Neuronal Damage after Traumatic Brain Injury. J. Neurotrauma 2005, 22, 83–94. [Google Scholar] [CrossRef]
- Weingarten, M.D.; Lockwood, A.H.; Hwo, S.Y.; Kirschner, M.W. A Protein Factor Essential for Microtubule Assembly. Proc. Natl. Acad. Sci. USA 1975, 72, 1858–1862. [Google Scholar] [CrossRef] [Green Version]
- Wagner, O.I.; Rammensee, S.; Korde, N.; Wen, Q.; Leterrier, J.F.; Janmey, P.A. Softness, Strength and Self-Repair in Intermediate Filament Networks. Exp. Cell Res. 2007, 313, 2228–2235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwata, N.; Tsubuki, S.; Takaki, Y.; Watanabe, K.; Sekiguchi, M.; Hosoki, E.; Kawashima-Morishima, M.; Lee, H.J.; Hama, E.; Sekine-Aizawa, Y.; et al. Identification of the Major Aβ1-42-Degrading Catabolic Pathway in Brain Parenchyma: Suppression Leads to Biochemical and Pathological Deposition. Nat. Med. 2000, 6, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Iwata, N.; Tsubuki, S.; Takaki, Y.; Shirotani, K.; Lu, B.; Gerard, N.P.; Gerard, C.; Hama, E.; Lee, H.J.; Saido, T.C. Metabolic Regulation of Brain Aβ by Neprilysin. Science 2001, 292, 1550–1552. [Google Scholar] [CrossRef]
- Johnson, V.E.; Stewart, W.; Graham, D.I.; Stewart, J.E.; Praestgaard, A.H.; Smith, D.H. A Neprilysin Polymorphism and Amyloid-β Plaques after Traumatic Brain Injury. J. Neurotrauma 2009, 26, 1197–1202. [Google Scholar] [CrossRef] [Green Version]
- Lipsky, R.H.; Marini, A.M. Brain-Derived Neurotrophic Factor in Neuronal Survival and Behavior-Related Plasticity. Ann. N. Y. Acad. Sci. 2007, 1122, 130–143. [Google Scholar] [CrossRef] [PubMed]
- McAllister, A.K.; Lo, D.C.; Katz, L.C. Neurotrophins Regulate Dendritic Growth in Developing Visual Cortex. Neuron 1995, 15, 791–803. [Google Scholar] [CrossRef] [Green Version]
- Lu, B. Pro-Region of Neurotrophins: Role in Synaptic Modulation. Neuron 2003, 39, 735–738. [Google Scholar] [CrossRef] [Green Version]
- Ali, F.; Meier, R. Primate Home Range and GRIN2A, a Receptor Gene Involved in Neuronal Plasticity: Implications for the Evolution of Spatial Memory. Genes Brain Behav. 2009, 8, 435–441. [Google Scholar] [CrossRef]
- De Quervain, D.J.F.; Papassotiropoulos, A. Identification of a Genetic Cluster Influencing Memory Performance and Hippocampal Activity in Humans. Proc. Natl. Acad. Sci. USA 2006, 103, 4270–4274. [Google Scholar] [CrossRef] [Green Version]
- Itokawa, M.; Yamada, K.; Iwayama-Shigeno, Y.; Ishitsuka, Y.; Detera-Wadleigh, S.; Yoshikawa, T. Genetic Analysis of a Functional GRIN2A Promoter (GT)n Repeat in Bipolar Disorder Pedigrees in Humans. Neurosci. Lett. 2003, 345, 53–56. [Google Scholar] [CrossRef]
- Itokawa, M.; Yamada, K.; Yoshitsugu, K.; Toyota, T.; Suga, T.; Ohba, H.; Watanabe, A.; Hattori, E.; Shimizu, H.; Kumakura, T.; et al. A Microsatellite Repeat in the Promoter of the N-Methyl-D-Aspartate Receptor 2A Subunit (GRIN2A) Gene Suppresses Transcriptional Activity and Correlates with Chronic Outcome in Schizophrenia. Pharmacogenetics 2003, 13, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Lipsky, R.H.; Sparling, M.B.; Ryan, L.M.; Xu, K.; Salazar, A.M.; Goldman, D.; Warden, D.L. Association of COMT Val158Met Genotype with Executive Functioning Following Traumatic Brain Injury. J. Neuropsychiatry Clin. Neurosci. 2005, 17, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Gallinat, J.; Bajbouj, M.; Sander, T.; Schlattmann, P.; Xu, K.; Ferro, E.F.; Goldman, D.; Winterer, G. Association of the G1947A COMT (Val(108/158)Met) Gene Polymorphism with Prefrontal P300 during Information Processing. Biol. Psychiatry 2003, 54, 40–48. [Google Scholar] [CrossRef]
- Chen, J.; Lipska, B.; Halim, N.; Ma, Q.; Matsumoto, M.; Melhem, S.; Kolachana, B.; Hyde, T.; Herman, M.; Apud, J.; et al. Functional Analysis of Genetic Variation in Catechol-O-Methyltransferase (COMT): Effects on MRNA, Protein, and Enzyme Activity in Postmortem Human Brain. Am. J. Hum. Genet. 2004, 75, 807–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nymberg, C.; Banaschewski, T.; Bokde, A.L.; Büchel, C.; Conrod, P.; Flor, H.; Frouin, V.; Garavan, H.; Gowland, P.; Heinz, A.; et al. DRD2/ANKK1 Polymorphism Modulates the Effect of Ventral Striatal Activation on Working Memory Performance. Neuropsychopharmacology 2014, 39, 2357–2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijayraghavan, S.; Wang, M.; Birnbaum, S.G.; Williams, G.V.; Arnsten, A.F. Inverted-U Dopamine D1 Receptor Actions on Prefrontal Neurons Engaged in Working Memory. Nat. Neurosci. 2007, 10, 376–384. [Google Scholar] [CrossRef]
- Neville, M.J.; Johnstone, E.C.; Walton, R.T. Identification and Characterization of ANKK1: A Novel Kinase Gene Closely Linked to DRD2 on Chromosome Band 11q23.1. Hum. Mutat. 2004, 23, 540–545. [Google Scholar] [CrossRef]
- Jönsson, E.G.; Nöthen, M.M.; Grünhage, F.; Farde, L.; Nakashima, Y.; Propping, P.; Sedvall, G.C. Polymorphisms in the Dopamine D2 Receptor Gene and Their Relationships to Striatal Dopamine Receptor Density of Healthy Volunteers. Mol. Psychiatry 1999, 4, 290–296. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, T.; Noble, E.P. Association of Seven Polymorphisms of the D2 Dopamine Receptor Gene with Brain Receptor-Binding Characteristics. Neurochem. Res. 2003, 28, 73–82. [Google Scholar] [CrossRef]
- Thompson, J.; Thomas, N.; Singleton, A.; Piggott, M.; Lloyd, S.; Perry, E.K.; Morris, C.M.; Perry, R.H.; Ferrier, I.N.; Court, J.A. D2 Dopamine Receptor Gene (DRD2) Taq 1 A Polymorphism: Reduced Dopamine D2 Receptor Binding in the Human Striatum Associated with the A1 Allele. Pharmacogenetics 1997, 7, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, Y.; Ishiguro, H.; Nankai, M.; Shibuya, H.; Watanabe, A.; Arinami, T. Identification of a Polymorphism in the Promoter Region of DRD4 Associated with the Human Novelty Seeking Personality Trait. Mol. Psychiatry 2000, 5, 64–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamidovic, A.; Dlugos, A.; Skol, A.; Palmer, A.A.; de Wit, H. Evaluation of Genetic Variability in the Dopamine Receptor D2 in Relation to Behavioral Inhibition and Impulsivity/Sensation Seeking: An Exploratory Study with d-Amphetamine in Healthy Participants. Exp. Clin. Psychopharmacol. 2009, 17, 374–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balestri, M.; Calati, R.; Serretti, A.; De Ronchi, D. Genetic Modulation of Personality Traits. Int. Clin. Psychopharmacol. 2014, 29, 1–15. [Google Scholar] [CrossRef]
- Heils, A.; Teufel, A.; Petri, S.; Stöber, G.; Riederer, P.; Bengel, D.; Lesch, K.P. Allelic Variation of Human Serotonin Transporter Gene Expression. J. Neurochem. 2002, 66, 2621–2624. [Google Scholar] [CrossRef]
- Lesch, K.P.; Bengel, D.; Heils, A.; Sabol, S.Z.; Greenberg, B.D.; Petri, S.; Benjamin, J.; Müller, C.R.; Hamer, D.H.; Murphy, D.L. Association of Anxiety-Related Traits with a Polymorphism in the Serotonin Transporter Gene Regulatory Region. Science 1996, 274, 1527–1531. [Google Scholar] [CrossRef]
- Stoltenberg, S.F.; Lehmann, M.K.; Anderson, C.; Nag, P.; Anagnopoulos, C. Serotonin Transporter (5-HTTLPR) Genotype and Childhood Trauma Are Associated with Individual Differences in Decision Making. Front. Genet. 2011, 2. [Google Scholar] [CrossRef] [Green Version]
- Sonuga-Barke, E.J.S.; Kumsta, R.; Schlotz, W.; Lasky-Su, J.; Marco, R.; Miranda, A.; Mulas, F.; Oades, R.D.; Banaschewski, T.; Mueller, U.; et al. A Functional Variant of the Serotonin Transporter Gene ((SLC6A4) Moderates Impulsive Choice in Attention-Deficit/Hyperactivity Disorder Boys and Siblings. Biol. Psychiatry 2011, 70, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Garry, P.; Ezra, M.; Rowland, M.; Westbrook, J.; Pattinson, K. The Role of the Nitric Oxide Pathway in Brain Injury and Its Treatment—From Bench to Bedside. Exp. Neurol. 2015, 263, 235–243. [Google Scholar] [CrossRef] [Green Version]
- Toda, N.; Ayajiki, K.; Okamura, T. Cerebral Blood Flow Regulation by Nitric Oxide: Recent Advances. Pharmacol. Rev. 2009, 61, 62–97. [Google Scholar] [CrossRef] [Green Version]
- Ahn, M.J.; Sherwood, E.R.; Prough, D.S.; Cheng, Y.L.; DeWitt, D.S. The Effects of Traumatic Brain Injury on Cerebral Blood Flow and Brain Tissue Nitric Oxide Levels and Cytokine Expression. J. Neurotrauma 2004, 21, 1431–1442. [Google Scholar] [CrossRef] [PubMed]
- Tuzgen, S.; Tanriover, N.; Uzan, M.; Tureci, E.; Tanriverdi, T.; Gumustas, K.; Kuday, C. Nitric Oxide Levels in Rat Cortex, Hippocampus, Cerebellum, and Brainstem after Impact Acceleration Head Injury. Neurol. Res. 2003, 25, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Asif, A.R.; Oellerich, M.; Armstrong, V.W.; Hecker, M.; Cattaruzza, M. T-786C Polymorphism of the Nos-3 Gene and the Endothelial Cell Response to Fluid Shear Stress—A Proteome Analysis. J. Proteome Res. 2009, 8, 3161–3168. [Google Scholar] [CrossRef] [PubMed]
- Robertson, C.S.; Gopinath, S.P.; Valadka, A.B.; Van, M.; Swank, P.R.; Goodman, J.C. Variants of the Endothelial Nitric Oxide Gene and Cerebral Blood Flow after Severe Traumatic Brain Injury. J. Neurotrauma 2011, 28, 727–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayed-Tabatabaei, F.A.; Oostra, B.A.; Isaacs, A.; Van Duijn, C.M.; Witteman, J.C.M. ACE Polymorphisms. Circ. Res. 2006, 1123–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariza, M.; del Matarin, M.; Junqué, C.; Mataró, M.; Clemente, I.; Moral, P.; Antonia Poca, M.; Garnacho, A.; Sahuquillo, J. Influence of Angiotensin-Converting Enzyme Polymorphism on Neuropsychological Subacute Performance in Moderate and Severe Traumatic. J. Neuropsychiatry Clin. Neurosci. 2006, 18, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Dardiotis, E.; Paterakis, K.; Siokas, V.; Tsivgoulis, G.; Dardioti, M.; Grigoriadis, S.; Simeonidou, C.; Komnos, A.; Kapsalaki, E.; Fountas, K.; et al. Effect of Angiotensin-Converting Enzyme Tag Single Nucleotide Polymorphisms on the Outcome of Patients with Traumatic Brain Injury. Pharm. Genom. 2015, 25, 485–490. [Google Scholar] [CrossRef]
- Oshima, T.; Lee, S.; Sato, A.; Oda, S.; Hirasawa, H.; Yamashita, T. TNF-α Contributes to Axonal Sprouting and Functional Recovery Following Traumatic Brain Injury. Brain Res. 2009, 1290, 102–110. [Google Scholar] [CrossRef]
- Bermpohl, D.; You, Z.; Korsmeyer, S.J.; Moskowitz, M.A.; Whalen, M.J. Traumatic Brain Injury in Mice Deficient in Bid: Effects on Histopathology and Functional Outcome. J. Cereb. Blood Flow Metab. 2006, 26, 625–633. [Google Scholar] [CrossRef] [Green Version]
- Waters, R.J.; Murray, G.D.; Teasdale, G.M.; Stewart, J.; Day, I.; Lee, R.J.; Nicoll, J.A.R. Cytokine Gene Polymorphisms and Outcome after Traumatic Brain Injury. J. Neurotrauma 2013, 30, 1710–1716. [Google Scholar] [CrossRef] [Green Version]
- Letterio, J.J.; Roberts, A.B. Regulation of immune responses by TGF-β. Annu. Rev. Immunol. 1998, 16, 137–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, R.; Hurley, C.K.; Posch, P.E. A Molecular Mechanism for the Differential Regulation of TGF-Β1 Expression Due to the Common SNP −509C-T (c. -1347C > T). Hum. Genet. 2006, 120, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Uzan, M.; Tanriverdi, T.; Baykara, O.; Kafadar, A.; Sanus, G.Z.; Tureci, E.; Ozkara, C.; Uysal, O.; Buyra, N. Association between Interleukin-1 Beta (IL-1β) Gene Polymorphism and Outcome after Head Injury: An Early Report. Acta Neurochir. 2005, 147, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The Pro- and Anti-Inflammatory Properties of the Cytokine Interleukin-6. Biochim. Biophys. Acta 2011, 1813, 878–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisgraber, K.H. Apolipoprotein E: Structure-Function Relationships. Adv. Protein Chem. 1994, 45, 249–302. [Google Scholar] [CrossRef] [PubMed]
- Abrahams, S.; Mc Fie, S.; Patricios, J.; Sutere, J.; Posthumus, M.; Septembera, A.V. An Association between Polymorphisms within the APOE Gene and Concussion Aetiology in Rugby Union Players. J. Sci. Med. Sport 2018, 21, 117–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, B.D.; Relkin, N.R.; Ravdin, L.D.; Jacobs, A.R.; Bennett, A.; Gandy, S. Apolipoprotein E Ε4 Associated with Chronic Traumatic Brain Injury in Boxing. J. Am. Med. Assoc. 1997, 278, 136–140. [Google Scholar] [CrossRef]
- Lawrence, D.W.; Comper, P.; Hutchison, M.G.; Sharma, B. The Role of Apolipoprotein E Episilon (ε)-4 Allele on Outcome Following Traumatic Brain Injury: A Systematic Review. Brain Inj. 2015, 29, 1018–1031. [Google Scholar] [CrossRef]
- Merritt, V.; Arnett, P. Apolipoprotein E (APOE) Ε4 Allele Is Associated with Increased Symptom Reporting Following Sports Concussion. J. Int. Neuropsychol. Soc. 2016, 22, 89–94. [Google Scholar] [CrossRef]
- Bogoslovsky, T.; Wilson, D.; Chen, Y.; Hanlon, D.; Gill, J.; Jeromin, A.; Song, L.; Moore, C.; Gong, Y.; Kenney, K.; et al. Increases of Plasma Levels of Glial Fibrillary Acidic Protein, Tau, and Amyloid β up to 90 Days after Traumatic Brain Injury. J. Neurotrauma 2017, 34, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Turner, R.C.; Lucke-Wold, B.P.; Robson, M.J.; Omalu, B.I.; Petraglia, A.L.; Bailes, J.E. Repetitive Traumatic Brain Injury and Development of Chronic Traumatic Encephalopathy: A Potential Role for Biomarkers in Diagnosis, Prognosis, and Treatment? Front. Neurol. 2012, 3, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Silva, R.; Lashley, T.; Strand, C.; Shiarli, A.-M.; Shi, J.; Tian, J.; Bailey, K.L.; Davies, P.; Bigio, E.H.; Arima, K.; et al. An Immunohistochemical Study of Cases of Sporadic and Inherited Frontotemporal Lobar Degeneration Using 3R- and 4R-Specific Tau Monoclonal Antibodies. Acta Neuropathol. 2006, 111, 329–340. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, I.; Schellenberg, G.D. Regulation of Tau Isoform Expression and Dementia. Biochim. Biophys. Acta 2005, 1739, 104–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Togo, T.; Sahara, N.; Yen, S.-H.; Cookson, N.; Ishizawa, T.; Hutton, M.; de Silva, R.; Lees, A.; Dickson, D.W. Argyrophilic Grain Disease Is a Sporadic 4-Repeat Tauopathy. J. Neuropathol. Exp. Neurol. 2002, 61, 547–556. [Google Scholar] [CrossRef] [Green Version]
- Sergeant, N.; Wattez, A.; Delacourte, A. Neurofibrillary Degeneration in Progressive Supranuclear Palsy and Corticobasal Degeneration: Tau Pathologies with Exclusively ‘Exon 10’ Isoforms. J. Neurochem. 1999, 72, 1243–1249. [Google Scholar] [CrossRef] [Green Version]
- Couchie, D.; Mavilia, C.; Georgieff, I.S.; Liem, R.K.; Shelanski, M.L.; Nunez, J. Primary Structure of High Molecular Weight Tau Present in the Peripheral Nervous System. Proc. Natl. Acad. Sci. USA 1992, 89, 4378–4381. [Google Scholar] [CrossRef] [Green Version]
- Poorkaj, P.; Bird, T.D.; Wijsman, E.; Nemens, E.; Garruto, R.M.; Anderson, L.; Andreadis, A.; Wiederholt, W.C.; Raskind, M.; Schellenberg, G.D. Tau Is a Candidate Gene for Chromosome 17 Frontotemporal Dementia. Ann. Neurol. 1998, 43, 815–825. [Google Scholar] [CrossRef]
- Mcdevitt, J.K.; Tierney, R.T.; Mansell, J.L.; Driban, J.B.; Higgins, M.; Toone, N.; Mishra, A.; Krynetskiy, E. Brain Injury Neuronal Structural Protein Polymorphism and Concussion in College Athletes Neuronal Structural Protein Polymorphism and Concussion in College Athletes. Brain Inj. 2011, 25, 1108–1113. [Google Scholar] [CrossRef]
- Egan, M.F.; Kojima, M.; Callicott, J.H.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M.; et al. The BDNF Val66met Polymorphism Affects Activity-Dependent Secretion of BDNF and Human Memory and Hippocampal Function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Felderhoff-Mueser, U.; Sifringer, M.; Pesditschek, S.; Kuckuck, H.; Moysich, A.; Bittigau, P.; Ikonomidou, C. Pathways Leading to Apoptotic Neurodegeneration Following Trauma to the Developing Rat Brain. Neurobiol. Dis. 2002, 11, 231–245. [Google Scholar] [CrossRef] [Green Version]
- Oyesiku, N.M.; Evans, C.O.; Houston, S.; Darrell, R.S.; Smith, J.S.; Fulop, Z.L.; Dixon, C.E.; Stein, D.G. Regional Changes in the Expression of Neurotrophic Factors and Their Receptors Following Acute Traumatic Brain Injury in the Adult Rat Brain. Brain Res. 1999, 833, 161–172. [Google Scholar] [CrossRef]
- Hicks, R.R.; Numan, S.; Dhillon, H.S.; Prasad, M.R.; Seroogy, K.B. Alterations in BDNF and NT-3 MRNAs in Rat Hippocampus after Experimental Brain Trauma. Mol. Brain Res. 1997, 48, 401–406. [Google Scholar] [CrossRef]
- Dretsch, M.N.; Williams, K.; Emmerich, T.; Crynen, G.; Ait-Ghezala, G.; Chaytow, H.; Mathura, V.; Crawford, F.C.; Iverson, G.L. Brain-Derived Neurotropic Factor Polymorphisms, Traumatic Stress, Mild Traumatic Brain Injury, and Combat Exposure Contribute to Postdeployment Traumatic Stress. Brain Behav. 2016, 6, e00392. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, V.; Veeramuthu, V.; Ahmad-Annuar, A.; Ramli, N.; Waran, V.; Chinna, K.; Bondi, M.W.; Delano-Wood, L.; Ganesan, D. Missense Mutation of Brain Derived Neurotrophic Factor (BDNF) Alters Neurocognitive Performance in Patients with Mild Traumatic Brain Injury: A Longitudinal Study. PLoS ONE 2016, 11, e0158838. [Google Scholar] [CrossRef] [PubMed]
- McDevitt, J.; Tierney, R.T.; Phillips, J.; Gaughan, J.P.; Torg, J.S.; Krynetskiy, E. Association between GRIN2A Promoter Polymorphism and Recovery from Concussion. Brain Inj. 2015, 29, 1674–1681. [Google Scholar] [CrossRef]
- Willmott, C.; Withiel, T.; Ponsford, J.; Burke, R. COMT Val158Met and Cognitive and Functional Outcomes after Traumatic Brain Injury. J. Neurotrauma 2014, 31, 1507–1514. [Google Scholar] [CrossRef]
- Riba, J.; Krämer, U.; Heldmann, M.; Richter, S.; Münte, T. Dopamine Agonist Increases Risk Taking but Blunts Reward-Related Brain Activity. PLoS ONE 2008, 3, e2479. [Google Scholar] [CrossRef] [Green Version]
- Dalley, J.; Roiser, J. Dopamine, Serotonin and Impulsivity. Neuroscience 2012, 215, 42–58. [Google Scholar] [CrossRef] [Green Version]
- McAllister, T.W.; Rhodes, C.H.; Flashman, L.A.; McDonald, B.C.; Belloni, D.; Saykin, A.J. Effect of the Dopamine D2 Receptor T Allele on Response Latency after Mild Traumatic Brain Injury. Am. J. Psychiatry 2005, 162, 1749–1751. [Google Scholar] [CrossRef]
- McAllister, T.W.; Flashman, L.A.; Harker Rhodes, C.; Tyler, A.L.; Moore, J.H.; Saykin, A.J.; McDonald, B.C.; Tosteson, T.D.; Tsongalis, G.J. Single Nucleotide Polymorphisms in ANKK1 and the Dopamine D2 Receptor Gene Affect Cognitive Outcome Shortly after Traumatic Brain Injury: A Replication and Extension Study. Brain Inj. 2008, 22, 705–714. [Google Scholar] [CrossRef] [Green Version]
- Yue, J.K.; Pronger, A.M.; Ferguson, A.R.; Temkin, N.R.; Sharma, S.; Rosand, J.; Sorani, M.D.; McAllister, T.W.; Barber, J.; Winkler, E.A.; et al. Association of a Common Genetic Variant within ANKK1 with Six-Month Cognitive Performance after Traumatic Brain Injury. Neurogenetics 2015, 16, 169–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Bertolino, A.; Fazio, L.; Blasi, G.; Rampino, A.; Romano, R.; Lee, M.T.; Xiao, T.; Papp, A.; Wang, D.; et al. Polymorphisms in Human Dopamine D2 Receptor Gene Affect Gene Expression, Splicing, and Neuronal Activity during Working Memory. Source 2007, 104, 20552–20557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrahams, S.; Mcfie, S.; Lacerda, M.; Patricios, J.; Suter, J.; September, A.V.; Posthumus, M. Unravelling the Interaction between the DRD2 and DRD4 Genes, Personality Traits and Concussion Risk. BMJ Open Sport Exerc. Med. 2019, 5, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.Z.; Lipsky, R.H.; Zhu, G.; Akhtar, L.A.; Taubman, J.; Greenberg, B.D.; Xu, K.; Arnold, P.D.; Richter, M.A.; Kennedy, J.L.; et al. Serotonin Transporter Promoter Gain-of-Function Genotypes Are Linked to Obsessive-Compulsive Disorder. Am. J. Hum. Genet. 2006, 78, 815–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zubkov, A.Y.; Pilkington, A.S.; Bernanke, D.H.; Parent, A.D.; Zhang, J. Posttraumatic Cerebral Vasospasm: Clinical and Morphological Presentations. J. Neurotrauma 1999, 16, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Chodobski, A.; Zink, B.J.; Szmydynger-Chodobska, J. Blood-Brain Barrier Pathophysiology in Traumatic Brain Injury. Transl Stroke Res. 2011, 2, 492–516. [Google Scholar] [CrossRef] [Green Version]
- Shohami, E.; Beit-Yannai, E.; Horowitz, M.; Kohen, R. Oxidative Stress in Closed-Head Injury: Brain Antioxidant Capacity as an Indicator of Functional Outcome. J. Cereb. Blood Flow Metab. 1997, 1007–1019. [Google Scholar] [CrossRef] [Green Version]
- Kroeger, K.M.; Carville, K.S.; Abraham, L.J. The -308 Tumor Necrosis Factor-α Promoter Polymorphism Effects Transcription. Mol. Immunol. 1997, 34, 391–399. [Google Scholar] [CrossRef]
- Dalgard, C.L.; Cole, J.T.; Kean, W.S.; Lucky, J.J.; Sukumar, G.; McMullen, D.C.; Pollard, H.B.; Watson, W.D. The Cytokine Temporal Profile in Rat Cortex after Controlled Cortical Impact. Front. Mol. Neurosci. 2012, 5. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
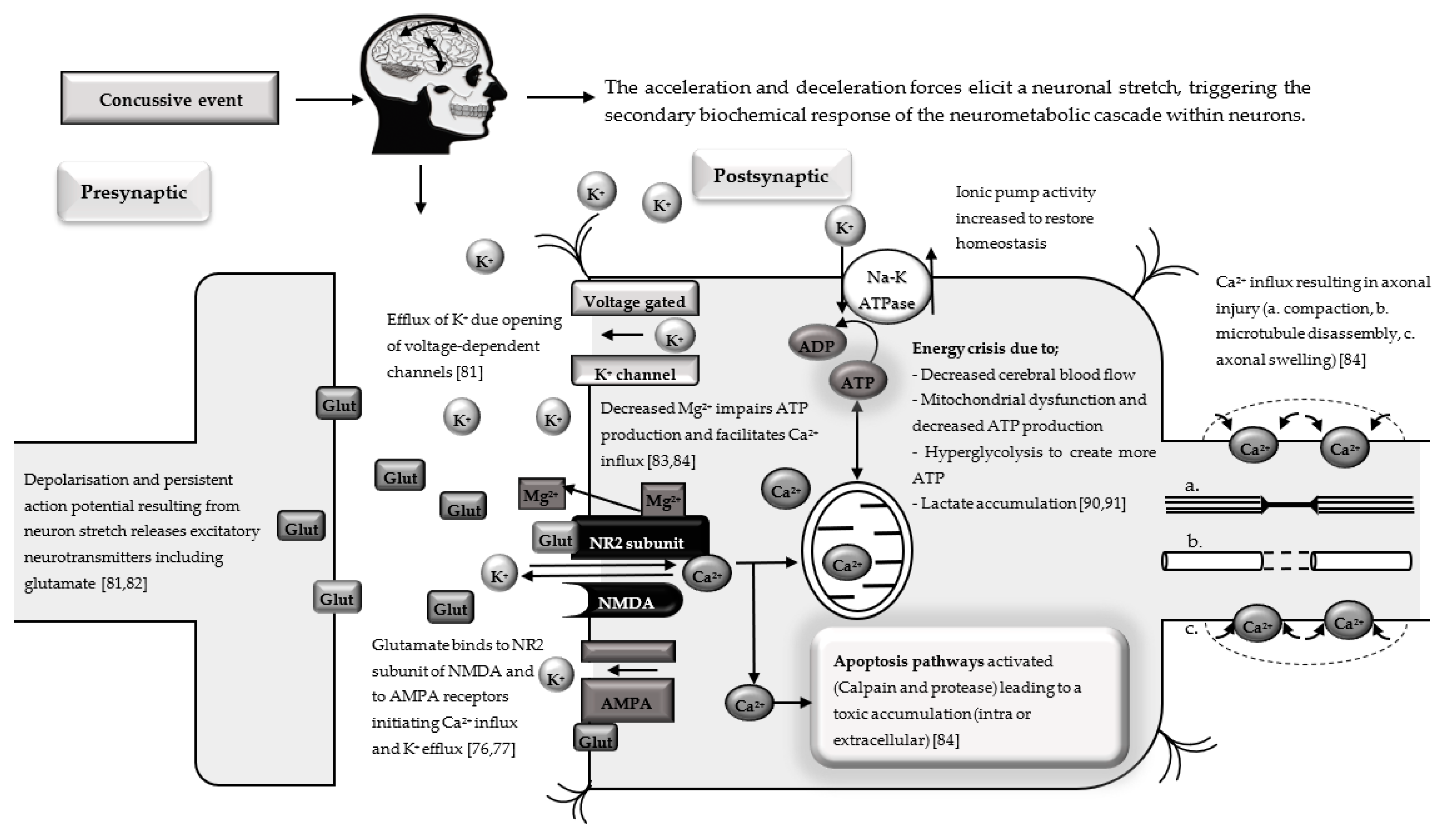{kind=link}
| Gene Name | Gene Abbreviation | Polymorphism Identifier | Relevant Effects Associated with TBI |
|---|---|---|---|
| Apolipoprotein E | APOE | rs429358 rs7412 rs405509 | Affects repair and plasticity of the brain [97,98]. APOE isoforms have differing effects on neurite extension, which can influence ability to recover post-concussion [97,98,99,100].Associated with functional regulation of APOE transcription [101,102]. |
| Microtubule-associated protein tau | MAPT | rs10445337 rs2435211 rs2435200 | Affects repair and plasticity of the brain via modulation of microtubule formation, structural stabilisation of the neuronal axons and drives growth of neurites [103,104]. |
| Neurofilament heavy | NEFH | rs165602 | Affects repair and plasticity of the brain via modulation of the neuronal cytoskeleton is to resist the resultant strain caused by biomechanical forces [105]. |
| Membrane metalloendopeptidase | MME | GT repeat promoter polymorphism of neprilysin | Affects repair and plasticity of the brain as this gene encodes for the neprilysin protease which degrades Aβ proteins [106,107,108]. |
| Brain-derived neurotrophic factorantisense RNA | BDNF-AS | rs6265 | Affects repair and plasticity of the brain via strengthening existing synaptic connections and modulating the creation of new synapses [109,110,111]. |
| Glutamate ionotropic receptor NMDA type subunit 2A promoter | GRIN2A | rs3219790 | Affects duration of concussion via potential modulation of glutamate-gated ion channel proteins [112,113,114,115]. |
| Catechol-O-methyltransferase | COMT | rs4680 | Affects cognitive behavioural capacity post-concussion and could increase impulsivity and risk taking [116,117,118]. |
| Ankyrin repeat and kinase domain containing 1 | ANKK1 | rs1800497 | Affects cognitive behavioural capacity via modulation of expression of D2 receptors [119,120,121,122,123,124]. |
| Dopamine receptor D2 Dopamine receptor D4 | DRD2 DRD4 | rs12364283 rs1076560 rs1800955 | Affects personality traits, associated with risk-taking behaviours (impulsivity, behavioural inhibition and novelty seeking) [125,126]. |
| Solute carrier family 6 member 4 | SLC6A4 | rs4795541 rs25531 | Reported to play a role in personality and behavior via increased harm avoidance and impulsivity behaviours [127,128,129,130,131]. |
| Endothelial nitric oxide synthase | NOS3 | rs2070744 | Could affect severity of concussion and cognitive behavioural capacity post-concussion via modulation of cerebral vasospasm [132,133,134,135,136,137]. |
| Angiotensin I-converting enzyme | ACE | rs4646994 rs7221780 rs8066276 | Affects cognitive behavioural capacity post-concussion via modulation of cerebral blood flow [138,139,140]. |
| Tumour necrosis factor | TNF | rs1800629 rs1800468 rs1800469 | Could affect neuroinflammation and severity of concussion [141,142,143]. |
| Transforming growth factor beta 1 | TGFB1 | rs1800468 rs1800469 | Regulation of the anti-inflammatory mediator TGFB1 could affect severity of concussion [144,145]. |
| Interleukin 1 alpha interleukin 1 beta | IL1A IL1B | rs1800587 rs16944 rs1143634 | Affects severity of TBI via potential modulation of the inflammatory process and secondary conditions [146]. |
| Interleukin 6 receptor | IL6R | rs2228145 | Affects severity of concussion potential via modulation of the inflammatory process and cognitive behavioural capacity post-concussion [147]. |
| APOE Isoform | APOE Genotype | rs429358 | rs7412 |
|---|---|---|---|
| ε2 | T | T | |
| ε3 | T | C | |
| ε4 | C | C | |
| ε2/ε2 | TT | TT | |
| ε2/ε3 | TT | CT | |
| ε2/ε4 | CT | CT | |
| ε3/ε3 | TT | CC | |
| ε3/ε4 | CT | CC | |
| ε4/ε4 | CC | CC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antrobus, M.R.; Brazier, J.; Stebbings, G.K.; Day, S.H.; Heffernan, S.M.; Kilduff, L.P.; Erskine, R.M.; Williams, A.G. Genetic Factors That Could Affect Concussion Risk in Elite Rugby. Sports 2021, 9, 19. https://doi.org/10.3390/sports9020019
Antrobus MR, Brazier J, Stebbings GK, Day SH, Heffernan SM, Kilduff LP, Erskine RM, Williams AG. Genetic Factors That Could Affect Concussion Risk in Elite Rugby. Sports. 2021; 9(2):19. https://doi.org/10.3390/sports9020019
Chicago/Turabian StyleAntrobus, Mark R., Jon Brazier, Georgina K. Stebbings, Stephen H. Day, Shane M. Heffernan, Liam P. Kilduff, Robert M. Erskine, and Alun G. Williams. 2021. "Genetic Factors That Could Affect Concussion Risk in Elite Rugby" Sports 9, no. 2: 19. https://doi.org/10.3390/sports9020019
APA StyleAntrobus, M. R., Brazier, J., Stebbings, G. K., Day, S. H., Heffernan, S. M., Kilduff, L. P., Erskine, R. M., & Williams, A. G. (2021). Genetic Factors That Could Affect Concussion Risk in Elite Rugby. Sports, 9(2), 19. https://doi.org/10.3390/sports9020019