Extensive Phylogenetic Analysis of Piscine Orthoreovirus Genomic Sequences Shows the Robustness of Subgenotype Classification

 , , ,
, , , 
Abstract
:1. Introduction
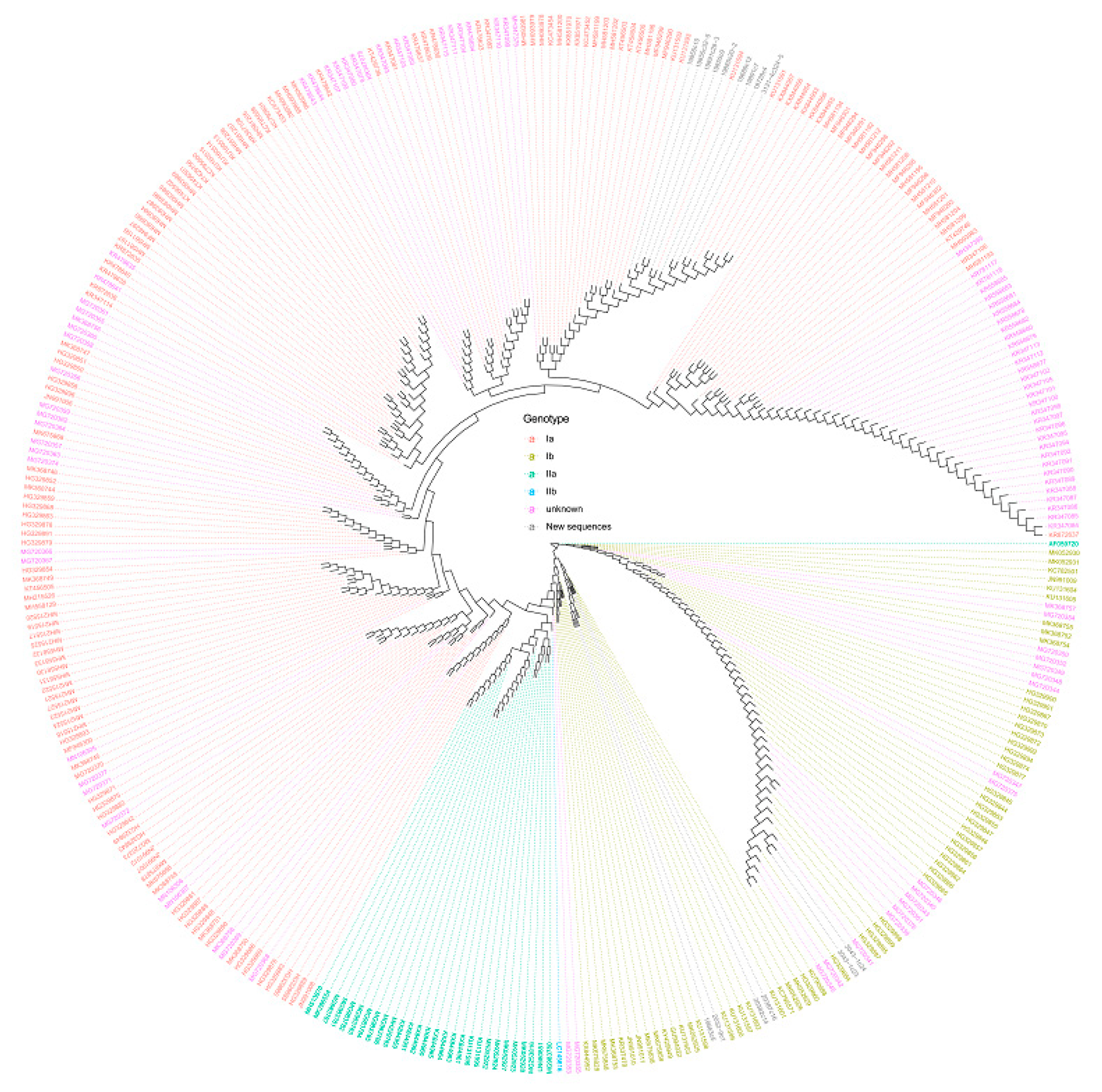
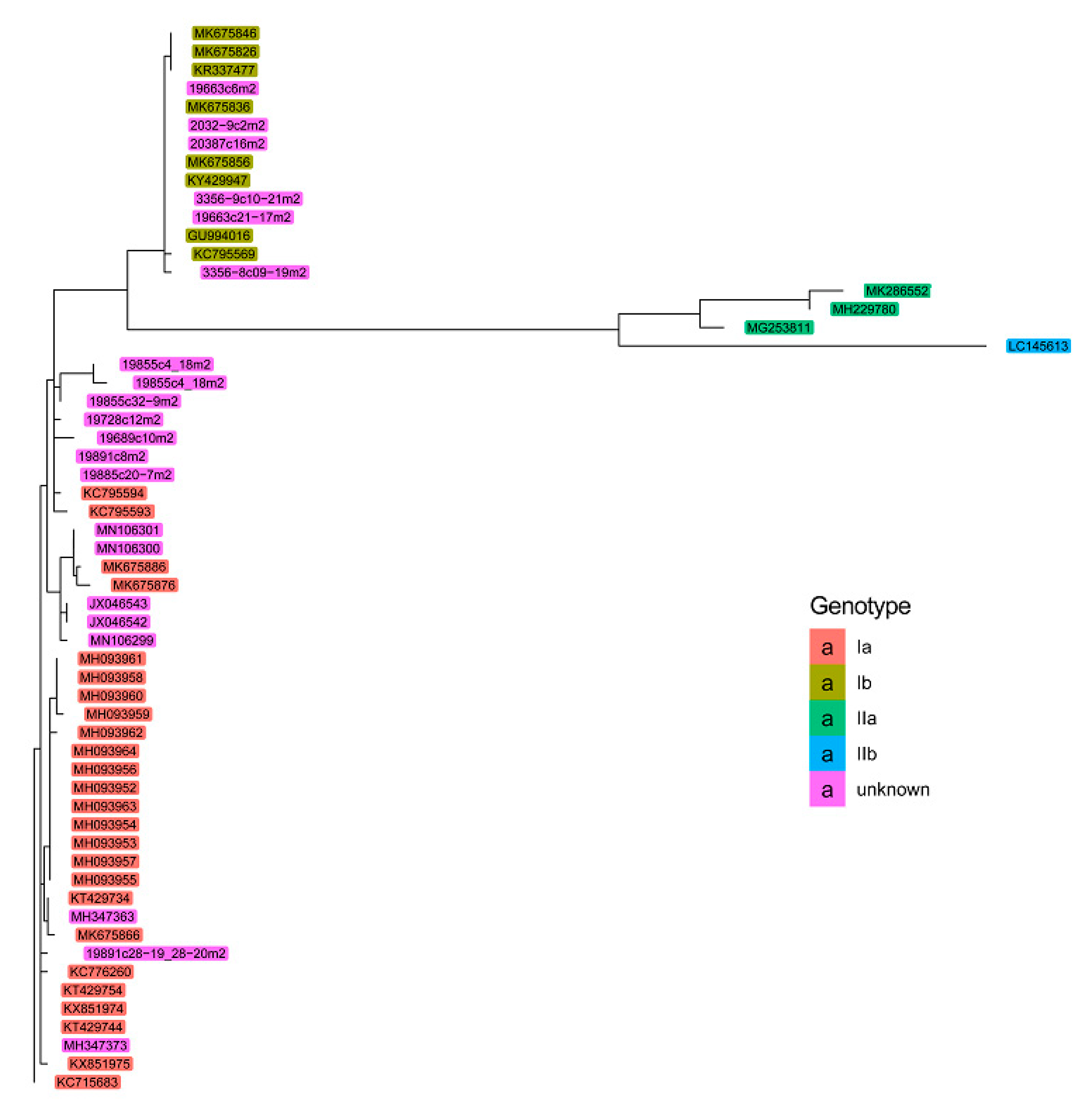
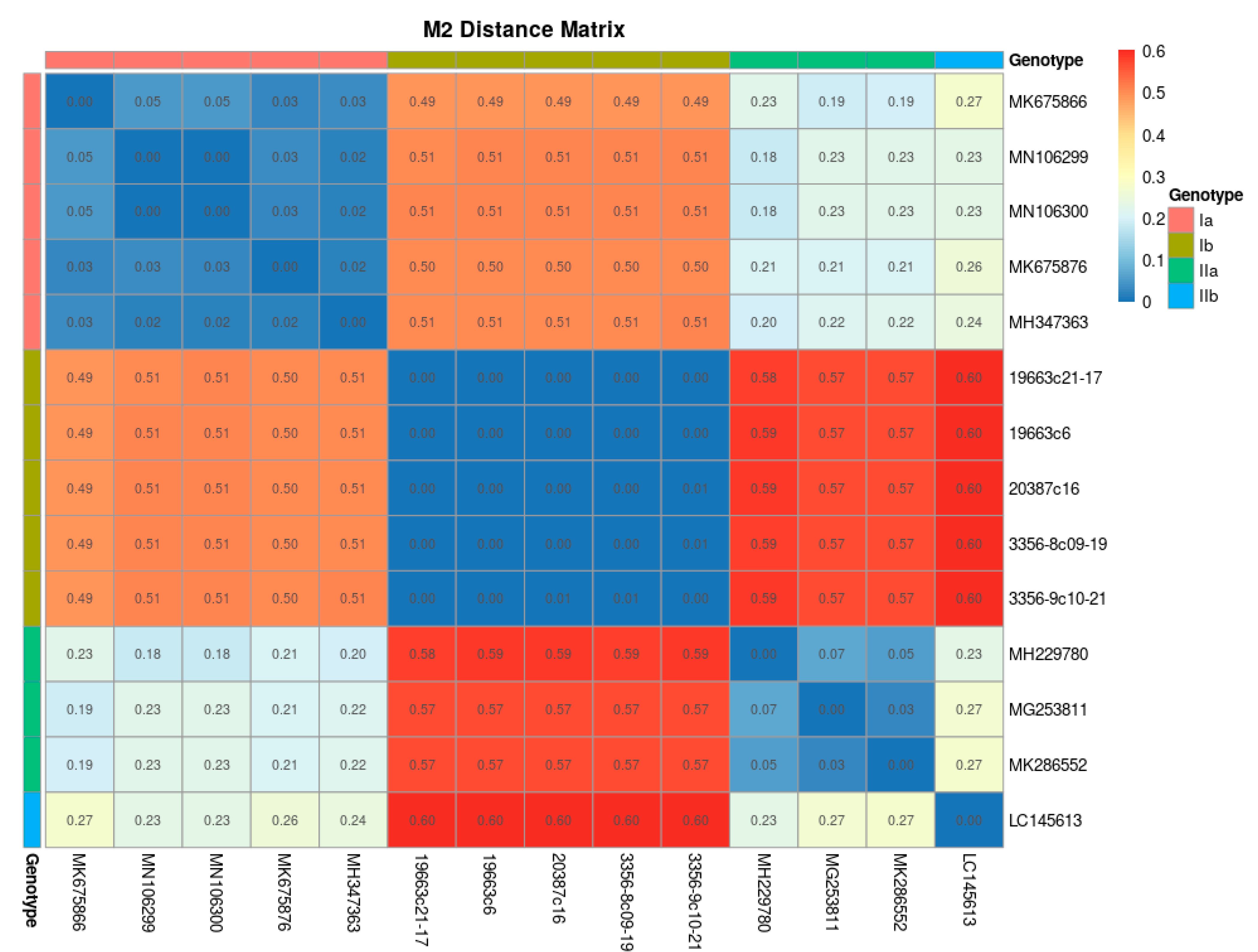
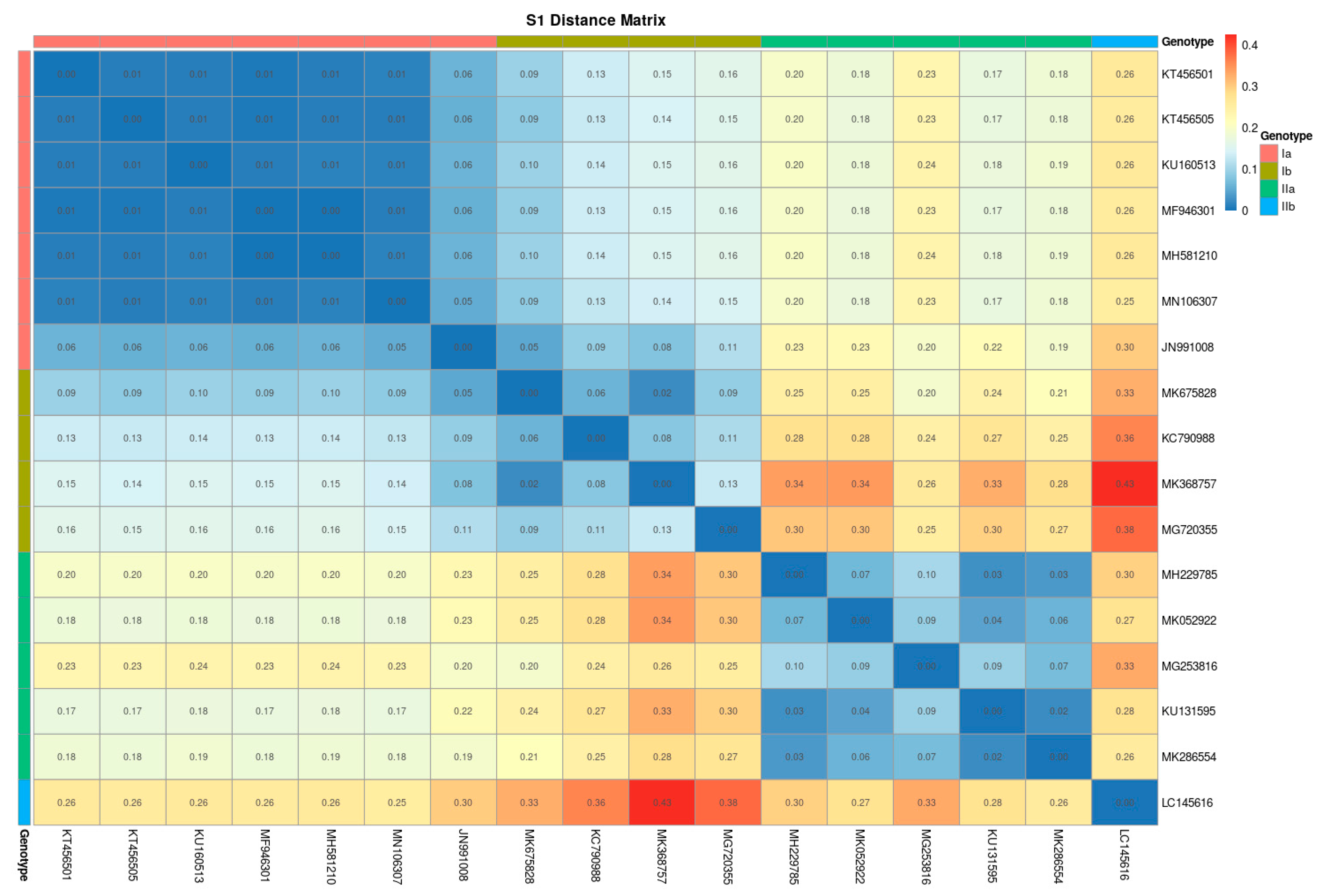
2. Results
Phylogenetic Analyses of PRV Genomic Sequences
3. Discussion
4. Materials and Methods
4.1. Extensive Phylogeny of PRV Segments
4.2. Nucleic Acid Sequencing of New Isolates
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Palacios, G.; Løvoll, M.; Tengs, T.; Hornig, M.; Hutchison, S.; Hui, J.; Kongtorp, R.-T.; Savji, N.; Bussetti, A.V.; Solovyov, A.; et al. Heart and Skeletal Muscle Inflammation of Farmed Salmon Is Associated with Infection with a Novel Reovirus. PLoS ONE 2010, 5, e11487. [Google Scholar] [CrossRef]
- Kongtorp, R.T.; Kjerstad, A.; Taksdal, T.; Guttvik, A.; Falk, K. Heart and skeletal muscle inflammation in Atlantic salmon, Salmo salar L.: A new infectious disease. J. Fish Dis. 2004, 27, 351–358. [Google Scholar] [CrossRef]
- Wessel, Ø.; Braaen, S.; Alarcón, M.; Haatveit, H.; Roos, N.; Markussen, T.; Tengs, T.; Dahle, M.K.; Rimstad, E. Infection with purified Piscine orthoreovirus demonstrates a causal relationship with heart and skeletal muscle inflammation in Atlantic salmon. PLoS ONE 2017, 12, e0183781. [Google Scholar] [CrossRef] [Green Version]
- Markussen, T.; Dahle, M.K.; Tengs, T.; Løvoll, M.T.; Finstad, Ø.W.; Wiik-Nielsen, C.R.; Grove, S.; Lauksund, R.S.; Robertsen, B.; Rimstad, E. Sequence Analysis of the Genome of Piscine Orthoreovirus (PRV) Associated with Heart and Skeletal Muscle Inflammation (HSMI) in Atlantic Salmon (Salmo salar). PLoS ONE 2013, 8, e70075. [Google Scholar] [CrossRef]
- Wiik-Nielsen, C.; Løvoll, M.; Sandlund, N.; Faller, R.; Wiik-Nielsen, J.; Jensen, B.B. First detection of piscine reovirus (PRV) in marine fish species. Dis. Aquat. Org. 2012, 97, 255–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garseth, Å.H.; Fritsvold, C.; Opheim, M.; Skjerve, E.; Biering, E. Piscine reovirus (PRV) in wild Atlantic salmon, Salmo salar L., and sea-trout, Salmo trutta L., in Norway. J. Fish Dis. 2012, 36, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.B.; Hjortaas, M.; Tengs, T.; Hellberg, H.; Johansen, R. First Description of a New Disease in Rainbow Trout (Oncorhynchus mykiss (Walbaum)) Similar to Heart and Skeletal Muscle Inflammation (HSMI) and Detection of a Gene Sequence Related to Piscine Orthoreovirus (PRV). PLoS ONE 2015, 10, e0131638. [Google Scholar] [CrossRef] [PubMed]
- Madhun, A.S.; Isachsen, C.H.; Omdal, L.M.; Einen, A.C.B.; Bjørn, P.A.; Nilsen, R.; Karlsbakk, E. Occurrence of salmonid alphavirus (SAV) and piscine orthoreovirus (PRV) infections in wild sea trout Salmo trutta in Norway. Dis. Aquat. Org. 2016, 120, 109–113. [Google Scholar] [CrossRef] [Green Version]
- Madhun, A.S.; Isachsen, C.H.; Omdal, L.M.; Einen, A.C.B.; Maehle, S.; Wennevik, V.; Niemelä, E.; Svåsand, T.; Karlsbakk, E. Prevalence of piscine orthoreovirus and salmonid alphavirus in sea-caught returning adult Atlantic salmon (Salmo salar L.) in northern Norway. J. Fish Dis. 2018, 41, 797–803. [Google Scholar] [CrossRef] [Green Version]
- Kibenge, M.J.T.; Iwamoto, T.; Wang, Y.; Morton, A.; Godoy, M.; Kibenge, F.S.B. Whole-genome analysis of piscine reovirus (PRV) shows PRV represents a new genus in family Reoviridae and its genome segment S1 sequences group it into two separate sub-genotypes. Virol. J. 2013, 10, 230. [Google Scholar] [CrossRef] [Green Version]
- Marty, G.D.; Morrison, D.B.; Bidulka, J.; Joseph, T.; Siah, A. Piscine reovirus in wild and farmed salmonids in British Columbia, Canada: 1974–2013. J. Fish Dis. 2015, 38, 713–728. [Google Scholar] [CrossRef]
- Siah, A.; Morrison, D.B.; Fringuelli, E.; Savage, P.S.; Richmond, Z.; Johns, R.; Purcell, M.K.; Johnson, S.C.; Saksida, S.M. Piscine Reovirus: Genomic and Molecular Phylogenetic Analysis from Farmed and Wild Salmonids Collected on the Canada/US Pacific Coast. PLoS ONE 2015, 10, e0141475. [Google Scholar] [CrossRef] [PubMed]
- Di Cicco, E.; Ferguson, H.W.; Schulze, A.D.; Kaukinen, K.H.; Li, S.; Vanderstichel, R.; Wessel, Ø.; Rimstad, E.; Gardner, I.A.; Hammell, K.L.; et al. Heart and skeletal muscle inflammation (HSMI) disease diagnosed on a British Columbia salmon farm through a longitudinal farm study. PLoS ONE 2017, 12, e0171471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Cicco, E.; Ferguson, H.W.; Kaukinen, K.H.; Schulze, A.D.; Li, S.; Tabata, A.; Günther, O.P.; Mordecai, G.; Suttle, C.A.; Miller, K.M. The same strain of Piscine orthoreovirus (PRV-1) is involved in the development of different, but related, diseases in Atlantic and Pacific Salmon in British Columbia. Facets 2018, 3, 599–641. [Google Scholar] [CrossRef] [Green Version]
- Godoy, M.G.; Kibenge, M.J.T.; Wang, Y.; Suarez, R.; Leiva, C.; Vallejos, F.; Kibenge, F.S.B. First description of clinical presentation of piscine orthoreovirus (PRV) infections in salmonid aquaculture in Chile and identification of a second genotype (Genotype II) of PRV. Virol. J. 2016, 13, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purcell, M.K.; Powers, R.L.; Evered, J.; Kerwin, J.; Meyers, T.R.; Stewart, B.; Winton, J.R. Molecular testing of adult Pacific salmon and trout (Oncorhynchus spp.) for several RNA viruses demonstrates widespread distribution of piscine orthoreo-virus (PRV) in Alaska and Washington. J. Fish Dis. 2018, 41, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Takano, T.; Nawata, A.; Sakai, T.; Matsuyama, T.; Ito, T.; Kurita, J.; Terashima, S.; Yasuike, M.; Nakamura, Y.; Fujiwara, A.; et al. Full-Genome Sequencing and Confirmation of the Causative Agent of Erythrocytic Inclusion Body Syndrome in Coho Salmon Identifies a New Type of Piscine Orthoreovirus. PLoS ONE 2016, 11, e0165424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunnarsdóttir, H.M. Screening for Viruses in Atlantic Salmon in Iceland. ISAV, PMCV and PRV. 2017. Available online: https://skemman.is/handle/1946/27629 (accessed on 6 January 2021).
- Kuehn, R.; Stoeckle, B.C.; Young, M.; Popp, L.; Taeubert, J.-E.; Pfaffl, M.W.; Geist, J. Identification of a piscine reovirus-related pathogen in proliferative darkening syndrome (PDS) infected brown trout (Salmo trutta fario) using a next-generation technology detection pipeline. PLoS ONE 2018, 13, e0206164. [Google Scholar] [CrossRef] [Green Version]
- Dhamotharan, K.; Vendramin, N.; Markussen, T.; Wessel, Ø.; Cuenca, A.; Nyman, I.B.; Olsen, A.B.; Tengs, T.; Dahle, M.K.; Rimstad, E. Molecular and Antigenic Characterization of Piscine orthoreovirus (PRV) from Rainbow Trout (Oncorhynchus mykiss). Viruses 2018, 10, 170. [Google Scholar] [CrossRef] [Green Version]
- Vendramin, N.; Cuenca, A.; Sørensen, J.; Alencar, A.L.F.; Christiansen, D.H.; Jacobsen, J.A.; Axen, C.; Lieffrig, F.; Ruane, N.; Martin, P.; et al. Presence and genetic variability of Piscine orthoreovirus genotype 1 (PRV-1) in wild salmonids in Northern Europe and North Atlantic Ocean. J. Fish Dis. 2019, 42, 1107–1118. [Google Scholar] [CrossRef]
- Bigarré, L.; Boitard, P.M.; Labrut, S.; Jamin, M. Point sur la Situation Epidemiologique du Syndrome HSMI Chez les Salmonides en France. Bulletin Epidémiologique. Santé Animale-Alimentation. 2018. Available online: https://be.anses.fr/sites/default/files/O-026_2018-06-22_HSMI_bigarre-VF.pdf (accessed on 6 January 2021).
- Adamek, M.; Hellmann, J.; Flamm, A.; Teitge, F.; Vendramin, N.; Fey, D.; Riße, K.; Blakey, F.; Rimstad, E.; Steinhagen, D. Detection of piscine orthoreoviruses (PRV-1 and PRV-3) in Atlantic salmon and rainbow trout farmed in Germany. Transbound. Emerg. Dis. 2019, 66, 14–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pojezdal, Ľ.; Adamek, M.; Syrová, E.; Steinhagen, D.; Minářová, H.; Papežíková, I.; Seidlova, V.; Reschová, S.; Palíková, M. Health Surveillance of Wild Brown Trout (Salmo trutta fario) in the Czech Republic Revealed a Coexistence of Proliferative Kidney Disease and Piscine Orthoreovirus-3 Infection. Pathogens 2020, 9, 604. [Google Scholar] [CrossRef] [PubMed]
- Kongtorp, R.T.; Taksdal, T.; Lyngøy, A. Pathology of heart and skeletal muscle inflammation (HSMI) in farmed Atlantic salmon Salmo salar. Dis. Aquat. Org. 2004, 59, 217–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, H.W.; Kongtorp, R.T.; Taksdal, T.; Graham, D.; Falk, K. An outbreak of disease resembling heart and skeletal muscle inflammation in Scottish farmed salmon, Salmo salar L., with observations on myocardial regeneration. J. Fish Dis. 2005, 28, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Finstad, Ø.W.; Dahle, M.K.; Lindholm, T.H.; Nyman, I.B.; Løvoll, M.; Wallace, C.; Olsen, C.M.; Storset, A.K.; Rimstad, E. Piscine orthoreovirus (PRV) infects Atlantic salmon erythrocytes. Vet. Res. 2014, 45, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wessel, Ø.; Olsen, C.M.; Rimstad, E.; Dahle, M.K. Piscine orthoreovirus (PRV) replicates in Atlantic salmon (Salmo salar L.) erythrocytes ex vivo. Vet. Res. 2015, 46, 26. [Google Scholar] [CrossRef] [Green Version]
- Garseth, Å.H.; Ekrem, T.; Biering, E. Phylogenetic Evidence of Long Distance Dispersal and Transmission of Piscine Reovirus (PRV) between Farmed and Wild Atlantic Salmon. PLoS ONE 2013, 8, e82202. [Google Scholar] [CrossRef]
- Kibenge, F.S.B. Emerging viruses in aquaculture. Curr. Opin. Virol. 2019, 34, 97–103. [Google Scholar] [CrossRef]
- Garver, K.A.; Marty, G.D.; Cockburn, S.N.; Richard, J.; Hawley, L.M.; Müller, A.; Thompson, R.L.; Purcell, M.K.; Saksida, S. Piscine reovirus, but not Jaundice Syndrome, was transmissible to Chinook Salmon, Oncorhynchus tshawytscha (Walbaum), Sockeye Salmon, Oncorhynchus nerka (Walbaum), and Atlantic Salmon, Salmo salar L. J. Fish Dis. 2016, 39, 117–128. [Google Scholar] [CrossRef]
- Siah, A.; Breyta, R.B.; Warheit, K.I.; Gagne, N.; Purcell, M.K.; Morrison, D.; Powell, J.F.F.; Johnson, S.C. Genomes reveal genetic diversity of Piscine orthoreovirus in farmed and free-ranging salmonids from Canada and USA. Virus Evol. 2020, 6, veaa054. [Google Scholar] [CrossRef]
- Polinski, M.P.; Marty, G.D.; Snyman, H.N.; Garver, K.A. Piscine othoreovirus demonstrates high infectivity but low virulence in Atlantic salmon of Pacific Canada. Sci. Rep. 2019, 9, 3297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhamotharan, K.; Tengs, T.; Wessel, Ø.; Braaen, S.; Nyman, I.B.; Hansen, E.F.; Christiansen, D.H.; Dahle, M.K.; Rimstad, E.; Markussen, T. Evolution of the Piscine orthoreovirus Genome Linked to Emergence of Heart and Skeletal Muscle Inflammation in Farmed Atlantic Salmon (Salmo salar). Viruses 2019, 11, 465. [Google Scholar] [CrossRef] [Green Version]
- Siah, A.; Knutsen, E.; Richmond, Z.; Mills, M.; Frisch, K.; Powell, J.F.F.; Brevik, Ø.; Duesund, H. Real-time RT-qPCR assay to detect sequences in the Piscine orthoreovirus-1 genome segment S1 associated with heart and skeletal muscle inflammation in Atlantic salmon. J. Fish Dis. 2020, 43, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Løvoll, M.; Alarcón, M.; Jensen, B.B.; Taksdal, T.; Kristoffersen, A.B.; Tengs, T. Quantification of piscine reovirus (PRV) at different stages of Atlantic salmon Salmo salar production. Dis. Aquat. Org. 2012, 99, 7–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wessel, Ø.; Hansen, E.F.; Dahle, M.K.; Alarcon, M.; Vatne, N.A.; Nyman, I.B.; Soleim, K.B.; Dhamotharan, K.; Timmerhaus, G.; Markussen, T.; et al. Piscine Orthoreovirus-1 Isolates Differ in Their Ability to Induce Heart and Skeletal Muscle Inflammation in Atlantic Salmon (Salmo salar). Pathogens 2020, 9, 1050. [Google Scholar] [CrossRef]
- Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2016, 44, D67–D72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winter, D.J. Rentrez: An R package for the NCBI eUtils. API R J. 2017, 9, 520–526. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Wheeler, T.J.; Kececioglu, J.D. Multiple alignment by aligning alignments. Bioinformatics 2007, 23, i559–i568. [Google Scholar] [CrossRef] [PubMed]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-coffee: A novel method for fast and accurate multiple sequence alignment. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Yu, G. Using ggtree to Visualize Data on Tree-Like Structures. Curr. Protoc. Bioinform. 2020, 69, e96. [Google Scholar] [CrossRef] [PubMed]
- Ginestet, C.E. ggplot2: Elegant Graphics for Data Analysis. J. R. Stat. Soc. Ser. A Stat. Soc. 2011, 174, 245–246. [Google Scholar] [CrossRef]
- Booth, D.S.; Szmidt-Middleton, H.; King, N.; Westbrook, M.J.; Young, S.L.; Kuo, A.; Abedin, M.; Chapman, J.; Fairclough, S.R.; Hellsten, U.; et al. RStudio: Integrated Development for R. Nature 2018. [Google Scholar] [CrossRef]
- McGinnis, S.; Madden, T.L. BLAST: At the core of a powerful and diverse set of sequence analysis tools. Nucleic Acids Res. 2004, 32, W20–W25. [Google Scholar] [CrossRef] [PubMed]
- Schliep, K.P. phangorn: Phylogenetic analysis in R. Bioinformatics 2010, 27, 592–593. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Reshaping Data with the reshape Package. J. Stat. Softw. 2007, 21, 1–20. [Google Scholar] [CrossRef]
- Kolde, R. Pheatmap: Pretty Heatmaps; R Package (Version 1.0.8). 2015. Available online: https://cran.r-project.org/web/packages/pheatmap/index.html (accessed on 6 January 2019).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PRV Genomic Segment | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| L1 | L2 | L3 | M1 | M2 | M3 | S1 | S2 | S3 | S4 | |
| Number of sequences | 53 | 38 | 40 | 41 | 61 | 43 | 390 | 101 | 42 | 100 |
| GenBank Accession Number | PRV Segment | PRV Subgenotype | Collection Date | Fish Species 1 | Location in Chile | Farming Conditions |
|---|---|---|---|---|---|---|
| MT598071 | M2 | Ib | 06-04-2018 | O. kisutch | Chiloé (Calen) | Seawater |
| MT598072 | M2 | Ia | 12-04-2018 | S. salar | Chiloé (Quellón) | Seawater |
| MT598073 | M2 | Ia | 26-04-2018 | O. kisutch | Chiloé (Quellón) | Seawater |
| MT598068 | M2 | Ia | 12-06-2018 | O. kisutch | Chiloé (Quellón) | Seawater |
| MT598074 | M2 | Ia | 19-06-2018 | O. kisutch | Chiloé (Caucahue) | Seawater |
| MT598069 | M2 | Ia | 21-06-2018 | O. kisutch | Chiloé (Detico) | Seawater |
| MT598075 | M2 | Ib | 27-03-2018 | O. kisutch | Pargua (P. Montt) 2 | Freshwater |
| MT598076 | M2 | Ib | 05-12-2018 | S. salar | Chiloé (Voigue) | Seawater |
| MT598077 | M2 | Ib | 19-12-2019 | S. salar | Lenca | rFeshwater |
| MT598078 | M2 | Ib | 19-12-2019 | S. salar | Lenca | Freshwater |
| MT598070 | M2 | Ib | 04-12-2018 | S. salar | Chiloé (Quicavi) | Seawater |
| MT598079 | S1 | Ib | 06-04-2018 | O. kisutch | Chiloé (Calen) | Seawater |
| MT598080 | S1 | Ia | 12-04-2018 | S. salar | Chiloé (Quellón) | Seawater |
| MT598081 | S1 | Ia | 26-04-2018 | O. kisutch | Chiloé (Quellón) | Seawater |
| MT598082 | S1 | Ia | 12-06-2018 | O. kisutch | Chiloé (Quellón) | Seawater |
| MT598083 | S1 | Ia | 19-06-2018 | O. kisutch | Chiloé (Caucahue) | Seawater |
| MT598084 | S1 | Ia | 21-06-2018 | O. kisutch | Chiloé (Detico) | Seawater |
| MT598085 | S1 | Ib | 27-03-2018 | O. kisutch | Pargua (P. Montt) 2 | Freshwater |
| MT598086 | S1 | Ib | 04-12-2018 | S. salar | Chiloé (Quicavi) | Seawater |
| MT598087 | S1 | Ib | 05-12-2018 | S. salar | Chiloé (Voigue) | Seawater |
| MT598088 | S1 | Ib | 17-06-2019 | S. salar | Pargua (P. Montt) 2 | Seawater |
| MT598089 | S1 | Ia | 07-08-2019 | O. kisutch | Puerto Cisne | Seawater |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godoy, M.; Medina, D.A.; Suarez, R.; Valenzuela, S.; Romero, J.; Kibenge, M.; Wang, Y.; Kibenge, F. Extensive Phylogenetic Analysis of Piscine Orthoreovirus Genomic Sequences Shows the Robustness of Subgenotype Classification. Pathogens 2021, 10, 41. https://doi.org/10.3390/pathogens10010041
Godoy M, Medina DA, Suarez R, Valenzuela S, Romero J, Kibenge M, Wang Y, Kibenge F. Extensive Phylogenetic Analysis of Piscine Orthoreovirus Genomic Sequences Shows the Robustness of Subgenotype Classification. Pathogens. 2021; 10(1):41. https://doi.org/10.3390/pathogens10010041
Chicago/Turabian StyleGodoy, Marcos, Daniel A. Medina, Rudy Suarez, Sandro Valenzuela, Jaime Romero, Molly Kibenge, Yingwei Wang, and Frederick Kibenge. 2021. "Extensive Phylogenetic Analysis of Piscine Orthoreovirus Genomic Sequences Shows the Robustness of Subgenotype Classification" Pathogens 10, no. 1: 41. https://doi.org/10.3390/pathogens10010041
APA StyleGodoy, M., Medina, D. A., Suarez, R., Valenzuela, S., Romero, J., Kibenge, M., Wang, Y., & Kibenge, F. (2021). Extensive Phylogenetic Analysis of Piscine Orthoreovirus Genomic Sequences Shows the Robustness of Subgenotype Classification. Pathogens, 10(1), 41. https://doi.org/10.3390/pathogens10010041