
Dynamics of Neospora caninum-Associated Abortions in a Dairy Sheep Flock and Results of a Test-and-Cull Control Programme

 , , , and
, , , and 
Abstract
:1. Introduction
2. Results
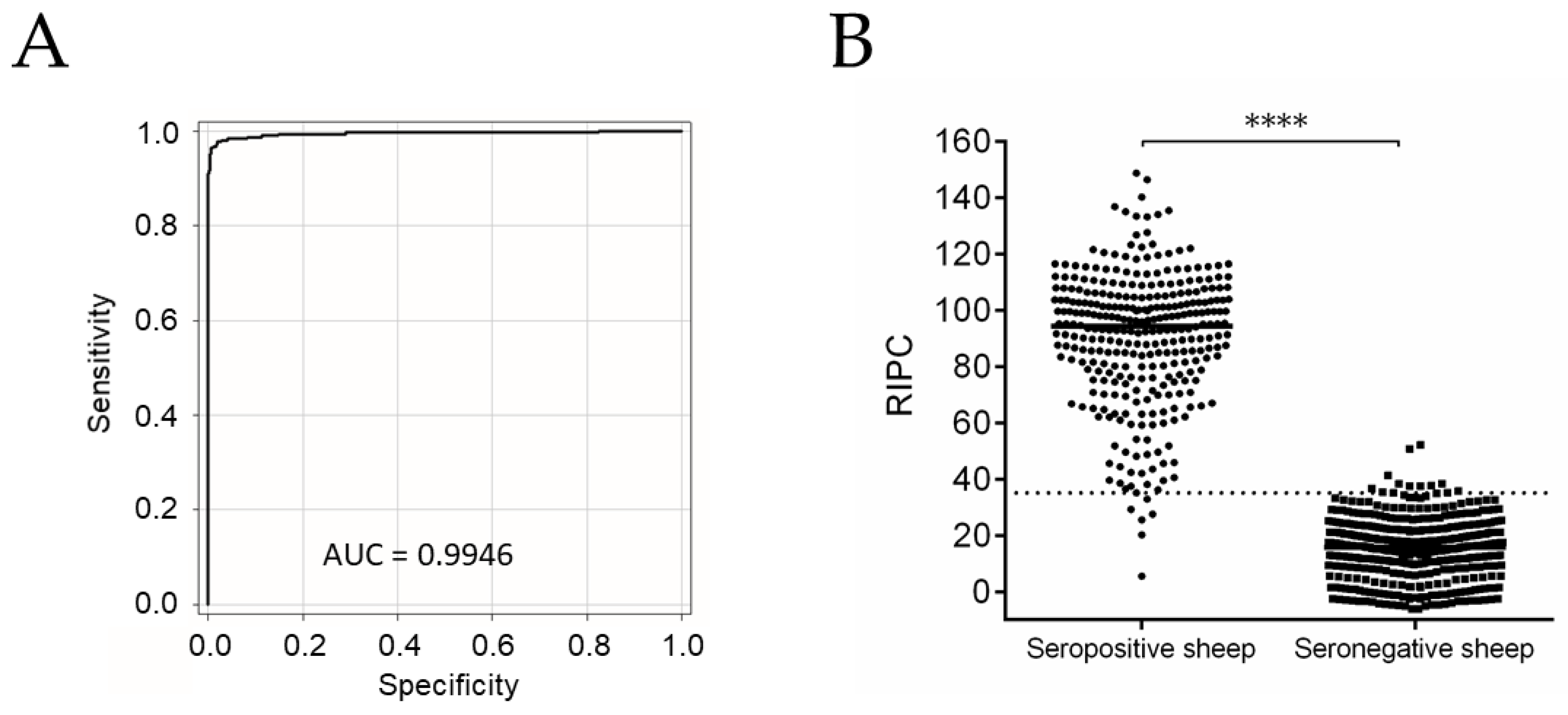
2.1. NcSALUVET ELISA: Accurate Serological Test for Naturally N. caninum-Infected Sheep
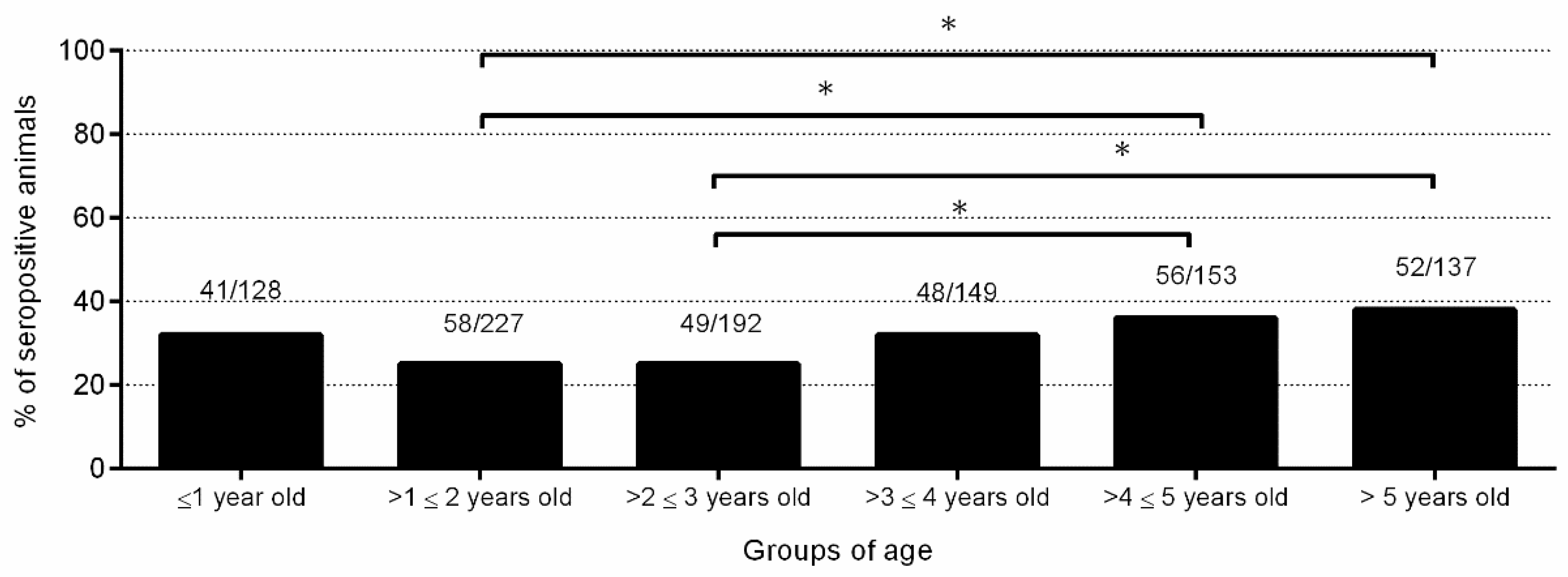
2.2. Neospora caninum Infection Was Widespread and a Major Cause of Abortion in the Flock
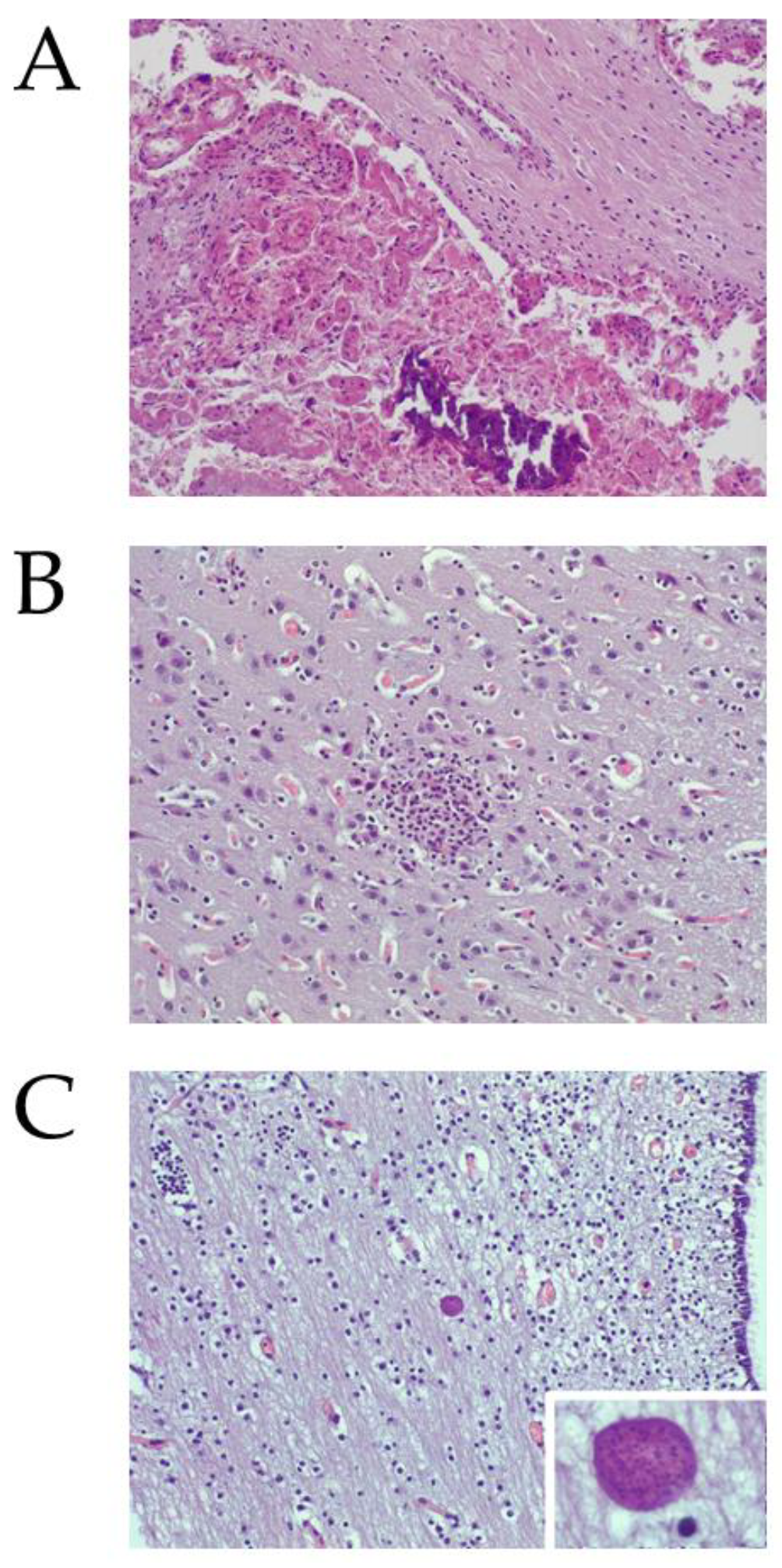
2.2.1. Neospora caninum DNA and Histological Lesions Were Detected in Placentas and Foetuses from Aborted Sheep
2.2.2. Abortions Were Associated with N. caninum Infection
2.2.3. Neospora caninum Seropositive Sheep Are More Likely to Have Repeated Abortions
2.3. Neospora caninum Genotyping Suggests a Unique Source of Infection in the Flock Maintained by Clonal Propagation
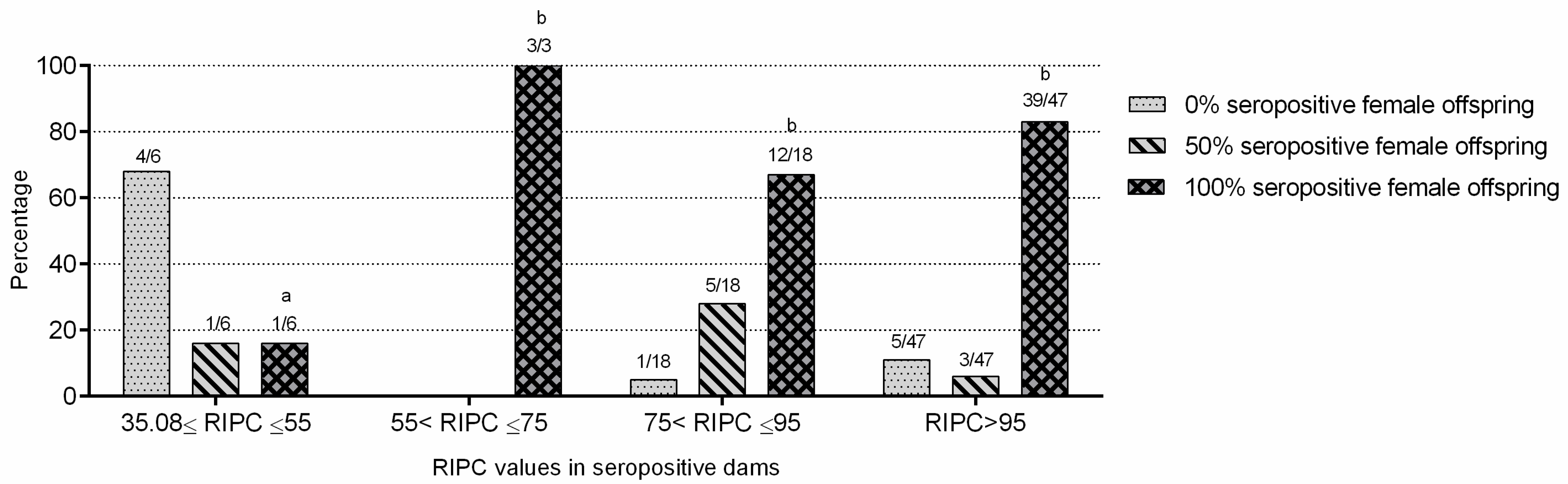
2.4. Neospora caninum Endogenous Transplacental Transmission Was Predominant in the Flock
2.5. Horizontal Transmission Might also Contribute to a Low Percentage of Infections
2.6. Progressive Selective Culling of N. caninum Seropositive Animals and Replacement with Seronegative Animals Reduced Abortion Rate in the Flock
3. Discussion
4. Materials and Methods
4.1. Flock Description and Collection of Data
4.2. Samplings
4.3. Parasites
4.4. Serological Assays
4.4.1. NcSALUVET ELISA
4.4.2. ID Screen® Neospora caninum Indirect ELISA
4.4.3. Western Blot
4.4.4. NcSALUVET Avidity ELISA
4.4.5. TgSALUVET ELISA
4.4.6. Indirect Fluorescence Antibody Test (IFAT)
4.5. Histological Examination
4.6. PCRs
4.7. Microsatellite Genotyping Method
4.8. Test and Cull Control Programme
4.9. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubey, J.P.; Hemphill, A.; Calero-Bernal, R.; Schares, G. Neosporosis in Animals; CRC Press: Boca Ratón, FL, USA, 2017. [Google Scholar]
- Trees, A.J.; Williams, D.J.L. Endogenous and exogenous transplacental infection in Neospora caninum and Toxoplasma Gondii. Trends Parasitol. 2005, 21, 558–561. [Google Scholar] [CrossRef]
- Reichel, M.P.; Alejandra Ayanegui-Alcérreca, M.; Gondim, L.F.P.; Ellis, J.T. What is the global economic impact of Neospora caninum in cattle—The billion dollar question. Int. J. Parasitol. 2013, 43, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Dubey, J.P.; Hartley, W.J.; Lindsay, D.S.; Topper, M.J. Fatal congenital Neospora caninum infection in a lamb. J. Parasitol. 1990, 76, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Hässig, M.; Sager, H.; Reitt, K.; Ziegler, D.; Strabel, D.; Gottstein, B. Neospora caninum in sheep: A herd case report. Vet. Parasitol. 2003, 117, 213–220. [Google Scholar] [CrossRef] [PubMed]
- West, D.M.; Pomroy, W.E.; Collett, M.G.; Hill, F.I.; Ridler, A.L.; Kenyon, P.R.; Morris, S.T.; Pattison, R.S. A possible role for Neospora caninum in ovine abortion in New Zealand. Small Rumin. Res. 2006, 62, 135–138. [Google Scholar] [CrossRef]
- Howe, L.; West, D.; Collett, M.; Tattersfield, G.; Pattison, R.S.; Pomroy, W.E.; Kenyon, P.R.; Morris, S.T.; Williamson, N.B. The role of Neospora caninum in three cases of unexplained ewe abortions in the southern North Island of New Zealand. Small Rumin. Res. 2008, 75, 115–122. [Google Scholar] [CrossRef]
- González-Warleta, M.; Castro-Hermida, J.A.; Regidor-Cerrillo, J.; Benavides, J.; Álvarez-García, G.; Fuertes, M.; Ortega-Mora, L.M.; Mezo, M. Neospora caninum infection as a cause of reproductive failure in a sheep flock. Vet. Res. 2014, 45, 88. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Warleta, M.; Castro-Hermida, J.A.; Calvo, C.; Perez, V.; Gutierrez-Exposito, D.; Regidor-Cerrillo, J.; Ortega-Mora, L.M.; Mezo, M. Endogenous transplacental transmission of Neospora caninum during successive pregnancies across three generations of naturally infected sheep. Vet. Res. 2018, 49, 106. [Google Scholar] [CrossRef] [Green Version]
- Hecker, Y.P.; Morrell, E.L.; Fiorentino, M.A.; Gual, I.; Rivera, E.; Fiorani, F.; Dorsch, M.A.; Gos, M.L.; Pardini, L.L.; Scioli, M.V. Ovine abortion by Neospora caninum: First case reported in Argentina. Acta Parasitol. 2019, 64, 950–955. [Google Scholar] [CrossRef]
- Romanelli, P.R.; de Matos, A.M.R.N.; Pinto-Ferreira, F.; Caldart, E.T.; Mareze, M.; Matos, R.L.N.; Freire, R.L.; Mitsuka-Breganó, R.; Headley, S.A.; Minho, A.P. Seroepidemiology of ovine toxoplasmosis and neosporosis in breeding rams from Rio Grande do Sul, Brazil. Transbound. Emerg. Dis. 2020, 67, 208–211. [Google Scholar] [CrossRef]
- Salaberry, S.R.S.; Okuda, L.H.; de Castro Nassar, A.F.; Castro, J.R.d.; Lima-Ribeiro, A.M.C. Prevalence of Neospora caninum antibodies in sheep flocks of Uberlândia county, MG. Rev. Bras. Parasitol. Vet. 2010, 19, 148–151. [Google Scholar] [CrossRef] [Green Version]
- Machado, G.P.; Kikuti, M.; Langoni, H.; Paes, A.C. Seroprevalence and risk factors associated with neosporosis in sheep and dogs from farms. Vet. Parasitol. 2011, 182, 356–358. [Google Scholar] [CrossRef]
- Villagra-Blanco, R.; Barrantes-Granados, O.; Montero-Caballero, D.; Romero-Zúñiga, J.J.; Dolz, G. Seroprevalence of Toxoplasma gondii and Neospora caninum infections and associated factors in sheep from Costa Rica. Parasite Epidemiol. Control. 2019, 4, e00085. [Google Scholar] [CrossRef]
- Romanelli, P.R.; de Matos, A.M.R.N.; Pinto-Ferreira, F.; Caldart, E.T.; do Carmo, J.L.M.; Santos, N.G.d.; Silva, N.R.d.; Loeffler, B.B.; Sanches, J.F.Z.; Francisquini, L.S. Anti-Toxoplasma gondii and anti-Neospora caninum antibodies in sheep from Paraná state, South Brazil: Prevalence and associated factors. Rev. Bras. Parasitol. Vet. 2021, 30, e023220. [Google Scholar] [CrossRef]
- Gazzonis, A.; Álvarez-García, G.; Zanzani, S.; Ortega-Mora, L.M.; Invernizzi, A.; Manfredi, M. Neospora caninum infection in sheep and goats from north-eastern Italy and associated risk factors. Small Rumin. Res. 2016, 140, 7–12. [Google Scholar] [CrossRef]
- Moreno, B.; Collantes-Fernandez, E.; Villa, A.; Navarro, A.; Regidor-Cerrillo, J.; Ortega-Mora, L.M. Occurrence of Neospora caninum and Toxoplasma gondii infections in ovine and caprine abortions. Vet. Parasitol. 2012, 187, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Razmi, G.; Naseri, Z. Molecular detection of Neospora caninum infection in ovine aborted foetuses in the Mashhad area, Iran. Ann. Parasitol. 2017, 63, 45–47. [Google Scholar]
- Amouei, A.; Sharif, M.; Sarvi, S.; Nejad, R.B.; Aghayan, S.A.; Hashemi-Soteh, M.B.; Mizani, A.; Hosseini, S.A.; Gholami, S.; Sadeghi, A. Aetiology of livestock fetal mortality in Mazandaran province, Iran. PeerJ 2019, 6, e5920. [Google Scholar] [CrossRef] [PubMed]
- Bartley, P.; Guido, S.; Mason, C.; Stevenson, H.; Chianini, F.; Carty, H.; Innes, E.; Katzer, F. Detection of Neospora caninum DNA in cases of bovine and ovine abortion in the South-West of Scotland. Parasitology 2019, 146, 979–982. [Google Scholar] [CrossRef]
- Horcajo, P.; Regidor-Cerrillo, J.; Aguado-Martínez, A.; Hemphill, A.; Ortega-Mora, L.M. Vaccines for bovine neosporosis: Current status and key aspects for development. Parasite Immunol. 2016, 38, 709–723. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Sánchez, R.; Vázquez, P.; Ferre, I.; Ortega-Mora, L.M. Treatment of toxoplasmosis and neosporosis in farm ruminants: State of knowledge and future trends. Curr. Top. Med. Chem. 2018, 18, 1304–1323. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Schares, G.; Ortega-Mora, L.M. Epidemiology and control of neosporosis and Neospora Caninum. Clin. Microbiol. Rev. 2007, 20, 323–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regidor-Cerrillo, J.; Diez-Fuertes, F.; Garcia-Culebras, A.; Moore, D.P.; Gonzalez-Warleta, M.; Cuevas, C.; Schares, G.; Katzer, F.; Pedraza-Diaz, S.; Mezo, M.; et al. Genetic diversity and geographic population structure of bovine Neospora caninum determined by microsatellite genotyping analysis. PLoS ONE 2013, 8, e72678. [Google Scholar] [CrossRef] [Green Version]
- Maia, M.O.; da Silva, A.R.S.; Gomes, A.A.D.; de Aguiar, D.M.; de Campos Pacheco, R.; da Costa, A.J.; dos Santos-Doni, T.R. Seroprevalence of Toxoplasma gondii and Neospora caninum in sheep intended for human consumption in the Rondônia state, Western Brazilian Amazon. Comp. Immunol. Microbiol. Infect. Dis. 2021, 74, 101599. [Google Scholar] [CrossRef] [PubMed]
- Selim, A.; Khater, H.; Almohammed, H.I. A recent update about seroprevalence of ovine neosporosis in Northern Egypt and its associated risk factors. Sci. Rep. 2021, 11, 14043. [Google Scholar] [CrossRef]
- Fthenakis, G.; Arsenos, G.; Brozos, C.; Fragkou, I.; Giadinis, N.; Giannenas, I.; Mavrogianni, V.; Papadopoulos, E.; Valasi, I. Health management of ewes during pregnancy. Anim. Reprod. Sci. 2012, 130, 198–212. [Google Scholar] [CrossRef]
- Dubey, J.P.; Murata, F.H.A.; Cerqueira-Cezar, C.K.; Kwok, O.C.H.; Su, C. Economic and public health importance of Toxoplasma gondii infections in sheep: 2009–2020. Vet. Parasitol. 2020, 286, 109195. [Google Scholar] [CrossRef] [PubMed]
- Soares, H.S.; Ahid, S.M.; Bezerra, A.C.; Pena, H.F.; Dias, R.A.; Gennari, S.M. Prevalence of anti-Toxoplasma gondii and anti-Neospora caninum antibodies in sheep from Mossoro, Rio Grande do Norte, Brazil. Vet. Parasitol. 2009, 160, 211–214. [Google Scholar] [CrossRef]
- Cosendey, R.; de Oliveira, F.; Frazão-Teixeira, E.; de Souza, G.; Brandão, F.; Ferreira, A.; Lilenbaum, W. Seroprevalence of anti-Neospora caninum antibodies in sheep from the rapidly expanding flock of Rio de Janeiro, Brazil. Vet. Parasitol. Reg. Stud. Rep. 2018, 14, 59–62. [Google Scholar] [CrossRef]
- Masala, G.; Porcu, R.; Daga, C.; Denti, S.; Canu, G.; Patta, C.; Tola, S. Detection of pathogens in ovine and caprine abortion samples from Sardinia, Italy, by PCR. J. Vet. Diagn. Invest. 2007, 19, 96–98. [Google Scholar] [CrossRef] [Green Version]
- Asadpour, R.; Jafari-Joozani, R.; Salehi, N. Detection of Neospora caninum in ovine abortion in Iran. J. Parasit. Dis. 2013, 37, 105–109. [Google Scholar] [CrossRef] [Green Version]
- Panadero, R.; Painceira, A.; López, C.; Vázquez, L.; Paz, A.; Díaz, P.; Dacal, V.; Cienfuegos, S.; Fernández, G.; Lago, N.; et al. Seroprevalence of Toxoplasma gondii and Neospora caninum in wild and domestic ruminants sharing pastures in Galicia (Northwest Spain). Res. Vet. Sci. 2010, 88, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Astorga, R.; Reguillo, L.; Hernández, M.; Cardoso-Toset, F.; Tarradas, C.; Maldonado, A.; Gómez-Laguna, J. Serosurvey on Schmallenberg virus and selected ovine reproductive pathogens in culled ewes from Southern Spain. Transbound. Emerg. Dis. 2014, 61, 4–11. [Google Scholar] [CrossRef]
- Díaz, J.M.; Fernández, G.; Prieto, A.; Valverde, S.; Lago, N.; Díaz, P.; Panadero, R.; López, C.; Morrondo, P.; Díez-Baños, P. Epidemiology of reproductive pathogens in semi-intensive lamb-producing flocks in North-West Spain: A comparative serological study. Vet. J. 2014, 200, 335–338. [Google Scholar] [CrossRef]
- Ruiz-Fons, F.; González-Barrio, D.; Aguilar-Ríos, F.; Soler, A.J.; Garde, J.J.; Gortázar, C.; del Rocío Fernández-Santos, M. Infectious pathogens potentially transmitted by semen of the black variety of the Manchega sheep breed: Health constraints for conservation purposes. Anim. Reprod. Sci. 2014, 149, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Gual, I.; Giannitti, F.; Hecker, Y.P.; Shivers, J.; Entrocassi, A.; Morrell, E.L.; Pardini, L.; Fiorentino, M.A.; Fermepin, M.R.; Unzaga, J.M. First case report of Toxoplasma gondii-induced abortions and stillbirths in sheep in Argentina. Vet. Parasitol. Reg Stud. Rep. 2018, 12, 39–42. [Google Scholar] [CrossRef]
- Edwards, J.F.; Dubey, J. Toxoplasma gondii abortion storm in sheep on a Texas farm and isolation of mouse virulent atypical genotype T gondii from an aborted lamb from a chronically infected ewe. Vet. Parasitol. 2013, 192, 129–136. [Google Scholar]
- Menzies, P.I. Chapter 90. Abortion in sheep: Diagnosis and control. In Current Therapy in Large Animal Theriogenology, 2nd ed.; Youngquist, R.S., Threlfall, W.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 667–680. [Google Scholar]
- Jolley, W.R.; McAllister, M.M.; McGuire, A.M.; Wills, R.A. Repetitive abortion in Neospora-infected ewes. Vet. Parasitol. 1999, 82, 251–257. [Google Scholar] [CrossRef]
- Almería, S.; López-Gatius, F.; García-Ispierto, I.; Nogareda, C.; Bech-Sabat, G.; Serrano, B.; Santolaria, P.; Yániz, J.L. Effects of crossbreed pregnancies on the abortion risk of Neospora caninum-infected dairy cows. Vet. Parasitol. 2009, 163, 323–329. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, I.M., Jr.; dos Santos Mesquita, L.E.; Miranda, D.N.P.; Gomes, T.A.; Vasconcelos, B.K.S.; Penha, L.C.; Silveira, L.C.S.; Redondo, A.R.R.; Costa, R.C.; Bruhn, F.R.P. Endogenous transplacental transmission of Neospora caninum in successive generations of congenitally infected goats. Vet. Parasitol. 2020, 284, 109191. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, L.P.; Nogueira, C.I.; Costa, R.C.; Orlando, D.R.; Bruhn, F.R.P.; Lopes, P.F.R.; Nakagaki, K.Y.R.; Peconick, A.P.; Seixas, J.N.; Júnior, P.S.B.; et al. Antibody kinetics in goats and conceptuses naturally infected with Neospora caninum. Vet. Parasitol. 2013, 196, 327–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GA Filho, P.C.; Oliveira, J.M.; Andrade, M.R.; Silva, J.G.; Kim, P.C.; Almeida, J.C.; Porto, W.J.; Mota, R.A. Incidence and vertical transmission rate of Neospora caninum in sheep. Comp. Immunol. Microbiol. Infect. Dis. 2017, 52, 19–22. [Google Scholar] [CrossRef]
- Feitosa, T.F.; Costa, F.T.R.; Bezerra, R.A.; Álvares, F.B.V.; Ferreira, L.C.; Mota, R.A.; Gennari, S.M.; Pena, H.F.J.; Azevedo, S.S.d.; Vilela, V.L.R. Vertical transmission and kinetic of antibodies anti-Neospora caninum in naturally infected lambs in the semiarid region of Brazil. Rev. Bras. Parasitol. Vet. 2021, 30, e010621. [Google Scholar] [CrossRef] [PubMed]
- Pare, J.; Thurmond, M.C.; Hietala, S.K. Neospora caninum antibodies in cows during pregnancy as a predictor of congenital infection and abortion. J. Parasitol. 1997, 83, 82–87. [Google Scholar] [CrossRef]
- Mazuz, M.L.; Fish, L.; Reznikov, D.; Wolkomirsky, R.; Leibovitz, B.; Savitzky, I.; Golenser, J.; Shkap, V. Neosporosis in naturally infected pregnant dairy cattle. Vet. Parasitol. 2014, 205, 85–91. [Google Scholar] [CrossRef]
- Wouda, W.; Moen, A.R.; Schukken, Y.H. Abortion risk in progeny of cows after a Neospora caninum epidemic. Theriogenology 1998, 49, 1311–1316. [Google Scholar] [CrossRef]
- García-Sánchez, M.; Moreno-Gonzalo, J.; González-Warleta, M.; Mezo, M.; Ortega-Mora, L.M.; Regidor-Cerrillo, J. Isolation and genetic characterization of Neospora caninum from naturally infected sheep. Vet. Parasitol. 2020, 280, 109091. [Google Scholar] [CrossRef]
- Syed-Hussain, S.; Howe, L.; Pomroy, W.; West, D.; Smith, S.; Williamson, N. Adaptation of a commercial ELISA to determine the IgG avidity in sheep experimentally and naturally infected with Neospora Caninum. Vet. Parasitol. 2014, 203, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Hietala, S.; Thurmond, M. Postnatal Neospora caninum transmission and transient serologic responses in two dairies. Int. J. Parasitol. 1999, 29, 1669–1676. [Google Scholar] [CrossRef]
- Ciupercescu, D. Dynamics of serum immunoglobulin concentrations in sheep during pregnancy and lactation. Res. Vet. Sci. 1977, 22, 23–27. [Google Scholar] [CrossRef]
- Regidor-Cerrillo, J.; Gomez-Bautista, M.; Sodupe, I.; Aduriz, G.; Alvarez-Garcia, G.; Del Pozo, I.; Ortega-Mora, L.M. In vitro invasion efficiency and intracellular proliferation rate comprise virulence-related phenotypic traits of Neospora Caninum. Vet. Res. 2011, 42, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Sánchez, R.; Ferre, I.; Re, M.; Vázquez, P.; Ferrer, L.M.; Blanco-Murcia, J.; Regidor-Cerrillo, J.; Díaz, M.P.; González-Huecas, M.; Tabanera, E.; et al. Safety and efficacy of the bumped kinase inhibitor BKI-1553 in pregnant sheep experimentally infected with Neospora caninum tachyzoites. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 112–124. [Google Scholar] [CrossRef]
- Álvarez-García, G.; Pereira-Bueno, J.; Gomez-Bautista, M.; Ortega-Mora, L.M. Pattern of recognition of Neospora caninum tachyzoite antigens by naturally infected pregnant cattle and aborted foetuses. Vet. Parasitol. 2002, 107, 15–27. [Google Scholar] [CrossRef]
- Aguado-Martínez, A.; Álvarez-García, G.; Arnaiz-Seco, I.; Innes, E.; Ortega-Mora, L.M. Use of avidity enzyme-linked immunosorbent assay and avidity Western blot to discriminate between acute and chronic Neospora caninum infection in cattle. J. Vet. Diagn. Investig. 2005, 17, 442–450. [Google Scholar] [CrossRef] [Green Version]
- Huertas-López, A.; Martínez-Subiela, S.; Cerón, J.J.; Vázquez-Calvo, Á.; Pazmiño-Bonilla, E.D.; López-Ureña, N.M.; Martínez-Carrasco, C.; Álvarez-García, G. Development and validation of a time-resolved fluorescence immunoassay for the detection of anti-Toxoplasma gondii antibodies in goats. Vet. Parasitol. 2021, 293, 109432. [Google Scholar] [CrossRef]
- Collantes-Fernández, E.; Gómez-Bautista, M.; Miró, G.; Álvarez-García, G.; Pereira-Bueno, J.; Frisuelos, C.; Ortega-Mora, L.M. Seroprevalence and risk factors associated with Neospora caninum infection in different dog populations in Spain. Vet. Parasitol. 2008, 152, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Müller, N.; Zimmermann, V.; Hentrich, B.; Gottstein, B. Diagnosis of Neospora caninum and Toxoplasma gondii infection by PCR and DNA hybridization immunoassay. J. Clin. Microbiol. 1996, 34, 2850–2852. [Google Scholar] [CrossRef] [Green Version]
- Hurtado, A.; Aduriz, G.; Moreno, B.; Barandika, J.; García-Pérez, A.L. Single tube nested PCR for the detection of Toxoplasma gondii in fetal tissues from naturally aborted ewes. Vet. Parasitol. 2001, 102, 17–27. [Google Scholar] [CrossRef]
- Greiner, M.; Gardner, I. Application of diagnostic tests in veterinary epidemiologic studies. Prev. Vet. Med. 2000, 45, 43–59. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microsatellite Genotype a | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Foetus | MS4 (AT)x | MS5 (TA)x | MS6A (TA)x | MS6B (AT)x | MS7 (TA)x | MS8 (AT)x | MS10 (ACT)x − (AGA)y − (TGA)z | MS12 (GT)x | MS21 (TACA)x |
| 1 b | 14 | 18 | 15 | 12 | 9.1SEQ | 13 | 6.14.9SEQ | 16 | 10 |
| 2 | 14 | 18 | 9.1SEQ | 13 | 6.14.9SEQ | 16 | 10 | ||
| 3 | 9.1 | 14 | 6.14.9 | 16 | |||||
| 4 b | 14 | 18 | 12 | 9.1 | 13 | 6.14.9 | 16 | 10 | |
| 5 b | 14 | 17 | 15 | 12 | 9.1 | 13 | 6.14.9 | 16 | 10 |
| 6 | 14 | 15 | 9.1 | 6.14.9SEQ | 16 | 10 | |||
| 7 | 18 | 12 | 9.1 | 13 | 6.14.9 | 16 | 10 | ||
| 8 | 14 | 9.1 | 13 | 6.14.9 | 16 | 10 | |||
| 9 b | 14 | 18 | 15 | 12 | 9.1 | 13 | 6.14.9 | 16 | 10 |
| 10 | 14 | 18 | 9.1 | 13 | 6.14.9 | 16 | 10 | ||
| 11 | 14 | 18 | 9.1 | 13 | 6.14.9 | 16 | 10 | ||
| 12 b | 14 | 18 | 15 | 12 | 9.1 | 13 | 6.14.9 | 16 | 10 |
| 13 b | 14 | 18 | 15 | 12 | 9.1 | 13 | 6.14.9 | 16 | 10 |
| 14 b | 14 | 18 | 15 | 9.1 | 13 | 6.15.9 | 16 | 10 | |
| 15 | 17/18 | 15 | 6.14.9 | 16 | 10 | ||||
| Dams | Female Offspring | ||
|---|---|---|---|
| Presence of Abortion | Serostatus | Presence of Abortion | Serostatus |
| Abortion (n = 84) | Seropositive (n = 45) | Abortion (n = 25; 55.5% *) | Seropositive (n = 22; 88%) |
| Seronegative (n = 3; 12%) | |||
| No abortion (n = 20; 44.5%) | Seropositive (n = 19; 95%) | ||
| Seronegative (n = 1; 5%) | |||
| Seronegative (n = 39) | Abortion (n = 6; 15.4% *) | Seropositive (n = 0; 0%) | |
| Seronegative (n = 6; 100%) | |||
| No abortion (n = 33; 84.6%) | Seropositive (n = 1; 3%) | ||
| Seronegative (n = 32; 97%) | |||
| No abortion (n = 161) | Seropositive (n = 29) | Abortion (n = 9; 31%) | Seropositive (n = 9; 100%) |
| Seronegative (n = 0; 0%) | |||
| No abortion (n = 20; 69%) | Seropositive (n = 14; 70%) | ||
| Seronegative (n = 6; 30%) | |||
| Seronegative (n = 132) | Abortion (n = 28; 21.2%) | Seropositive (n = 2; 7%) | |
| Seronegative (n = 26; 93%) | |||
| No abortion (n = 104; 78.8%) | Seropositive (n = 4; 3.8%) | ||
| Seronegative (n = 100; 96.2%) | |||
| Year | Number of Female Adult Sheep | Number of Aborted Sheep (% Abortion Rate) | Number of Culled Animals (% Culling Rate) | Number of Replaced Animals (% Replacement Rate) | |
|---|---|---|---|---|---|
| Prior to the control programme | 2011 | 678 | 55 (8.1%) | 76 (11.2%) | 95 (14%) |
| 2012 | 697 | 91 (13.1%) | 199 (28.6%) | 212 (30.4%) | |
| 2013 | 710 | 159 (22.4%) | 142 (20%) | 210 (29.6%) | |
| 2014 | 778 | 146 (18.8%) | 228 (29.3%) | 271 (34.8%) | |
| 2015 | 821 | 131 (16%) | 173 (21.1%) | 178 (21.7%) | |
| 2016 | 826 | 164 (19.9%) | 152 (18.4%) | 181 (21.9%) | |
| 2017 | 855 | 141 (16.5%) | 300 (35.1%) | 307 (35.9%) | |
| 2018 | 862 | 221 (25.6%) | 350 (40.6%) | 300 (34.8%) | |
| During the control programme | 2019 | 812 | 132 (16.3%) | 266 (32.8%) | 373 (46%) |
| 2020 | 919 | 66 (7.2%) | 208 (22.6%) | 299 (32.5%) |
| Sampling Date | Number of Sampled Animals | Samples | Techniques Employed (Number of Samples Analysed) |
|---|---|---|---|
| March–May 2018 | 986 female sheep | Serum | NcSALUVET ELISA (all samples) |
| NcSALUVET avidity ELISA (in 8 seropositive aborted animals and in 6 seropositive non-aborted animals) | |||
| TgSALUVET ELISA (in 160 aborted animals) | |||
| March 2018 | 2 dogs | Serum | IFAT (all samples) |
| During 2018 | 50 aborted sheep | Brains of 39 aborted foetuses (from 34 sheep) and 16 placentas (cotyledons) from 16 sheep were analysed | N. caninum PCR (all samples) |
| Microsatellite genotyping (in 15 N. caninum PCR-positive foetal brains) | |||
| T. gondii PCR (all samples) | |||
| Histology (HE staining) (all samples) | |||
| From June 2018 to December 2020 (control program) | 972 female lambs (one month of age) born from N. caninum seronegative dams. Of them, 278 were resampled at 7–10 months of age. | Serum | NcSALUVET ELISA (all samples) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Sánchez, R.; Vázquez-Calvo, Á.; Fernández-Escobar, M.; Regidor-Cerrillo, J.; Benavides, J.; Gutiérrez, J.; Gutiérrez-Expósito, D.; Crespo-Ramos, F.J.; Ortega-Mora, L.M.; Álvarez-García, G. Dynamics of Neospora caninum-Associated Abortions in a Dairy Sheep Flock and Results of a Test-and-Cull Control Programme. Pathogens 2021, 10, 1518. https://doi.org/10.3390/pathogens10111518
Sánchez-Sánchez R, Vázquez-Calvo Á, Fernández-Escobar M, Regidor-Cerrillo J, Benavides J, Gutiérrez J, Gutiérrez-Expósito D, Crespo-Ramos FJ, Ortega-Mora LM, Álvarez-García G. Dynamics of Neospora caninum-Associated Abortions in a Dairy Sheep Flock and Results of a Test-and-Cull Control Programme. Pathogens. 2021; 10(11):1518. https://doi.org/10.3390/pathogens10111518
Chicago/Turabian StyleSánchez-Sánchez, Roberto, Ángela Vázquez-Calvo, Mercedes Fernández-Escobar, Javier Regidor-Cerrillo, Julio Benavides, Jorge Gutiérrez, Daniel Gutiérrez-Expósito, Francisco José Crespo-Ramos, Luis Miguel Ortega-Mora, and Gema Álvarez-García. 2021. "Dynamics of Neospora caninum-Associated Abortions in a Dairy Sheep Flock and Results of a Test-and-Cull Control Programme" Pathogens 10, no. 11: 1518. https://doi.org/10.3390/pathogens10111518
APA StyleSánchez-Sánchez, R., Vázquez-Calvo, Á., Fernández-Escobar, M., Regidor-Cerrillo, J., Benavides, J., Gutiérrez, J., Gutiérrez-Expósito, D., Crespo-Ramos, F. J., Ortega-Mora, L. M., & Álvarez-García, G. (2021). Dynamics of Neospora caninum-Associated Abortions in a Dairy Sheep Flock and Results of a Test-and-Cull Control Programme. Pathogens, 10(11), 1518. https://doi.org/10.3390/pathogens10111518