
Global Distribution of Babesia Species in Questing Ticks: A Systematic Review and Meta-Analysis Based on Published Literature




Abstract
:1. Introduction
2. Results
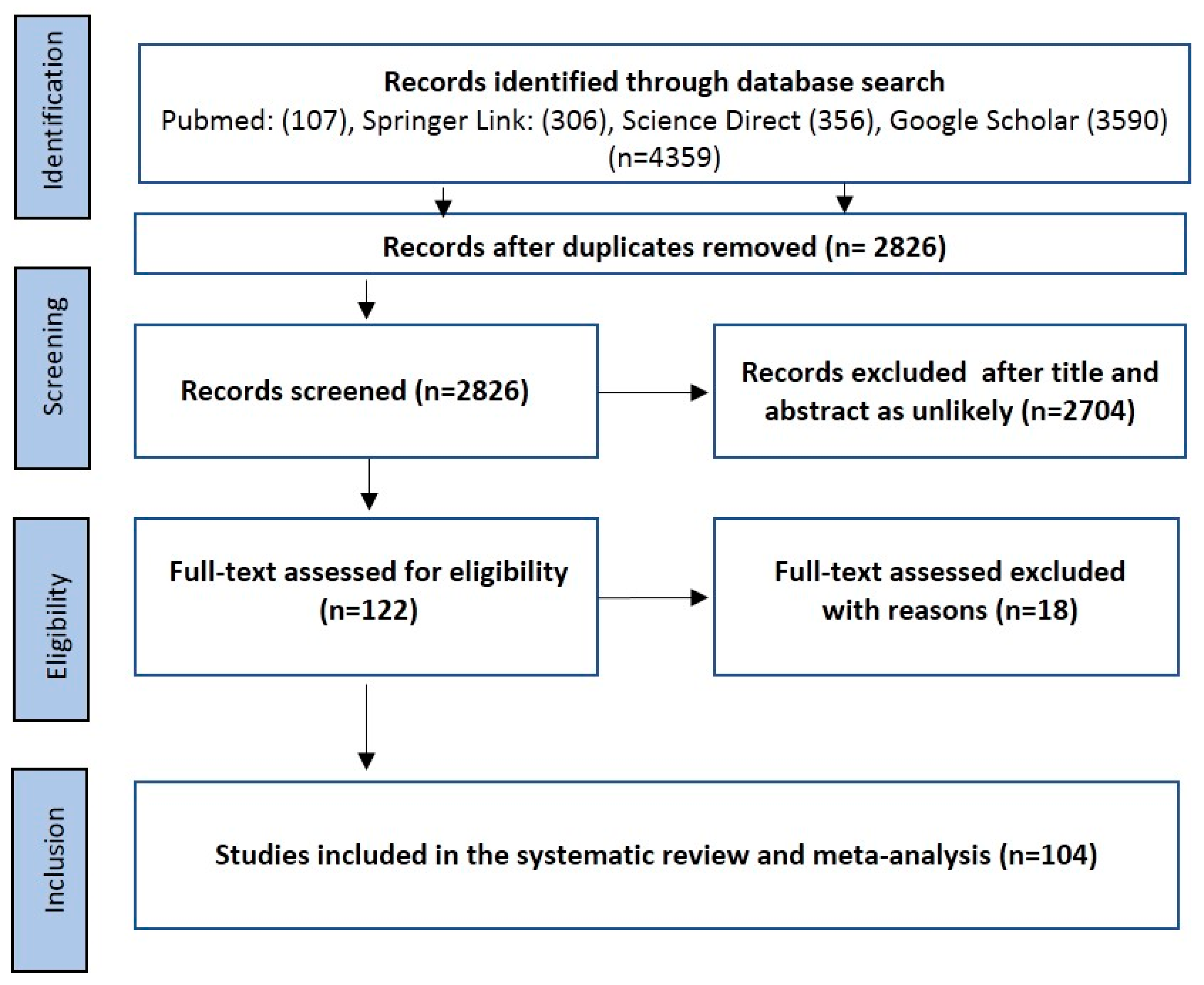
2.1. Literature Search and Eligible Studies
2.2. Characteristics of Eligible Studies
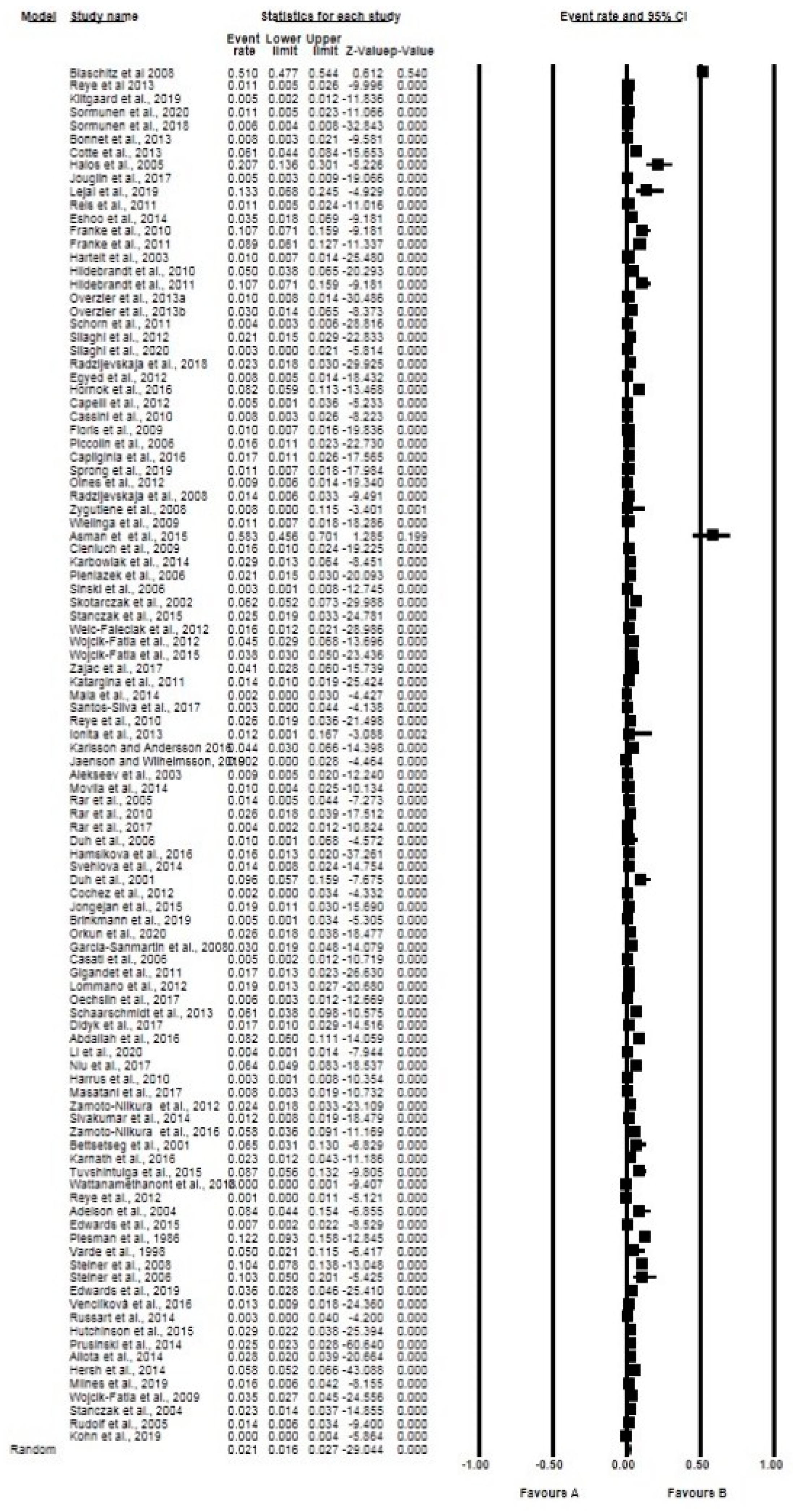
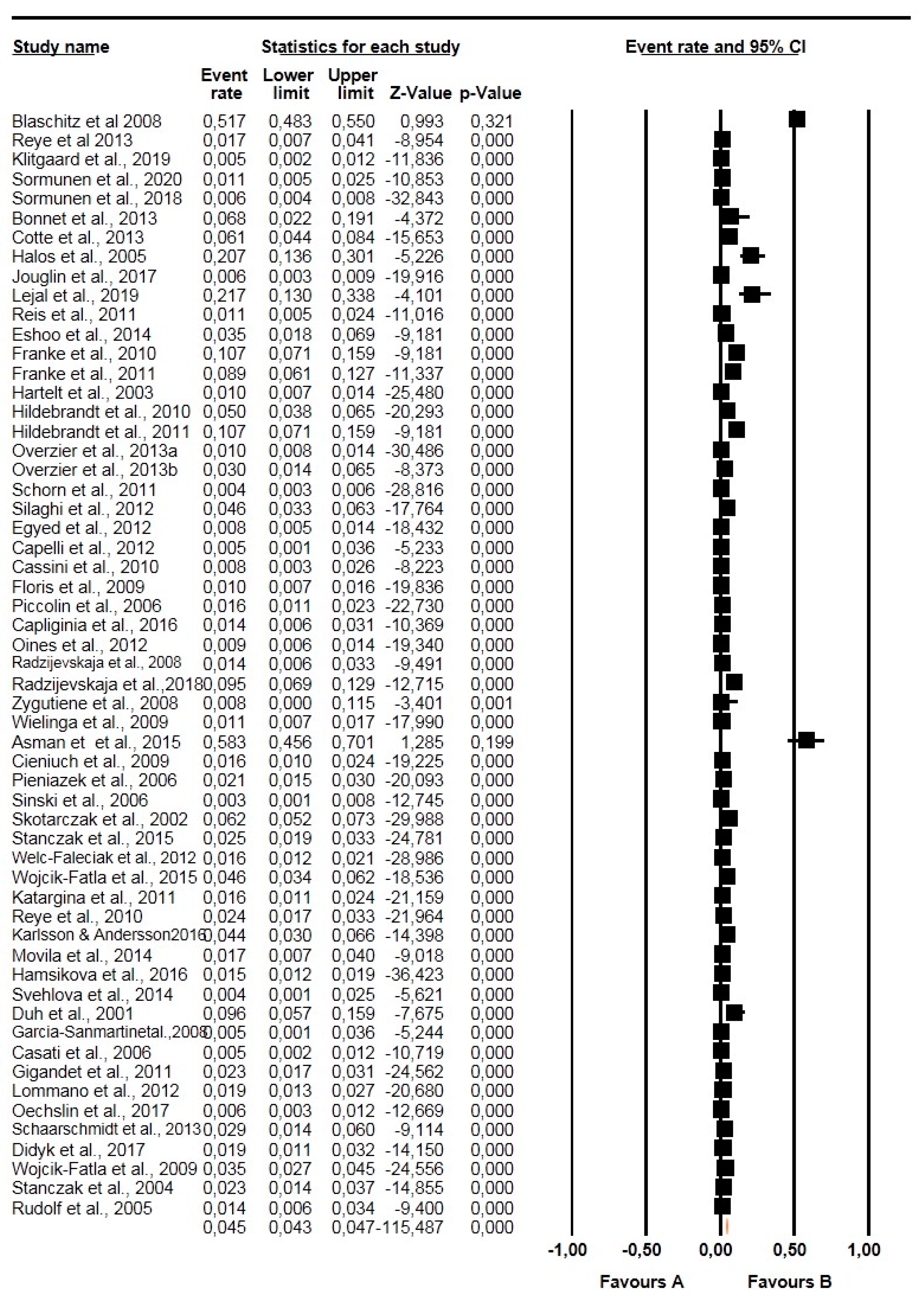
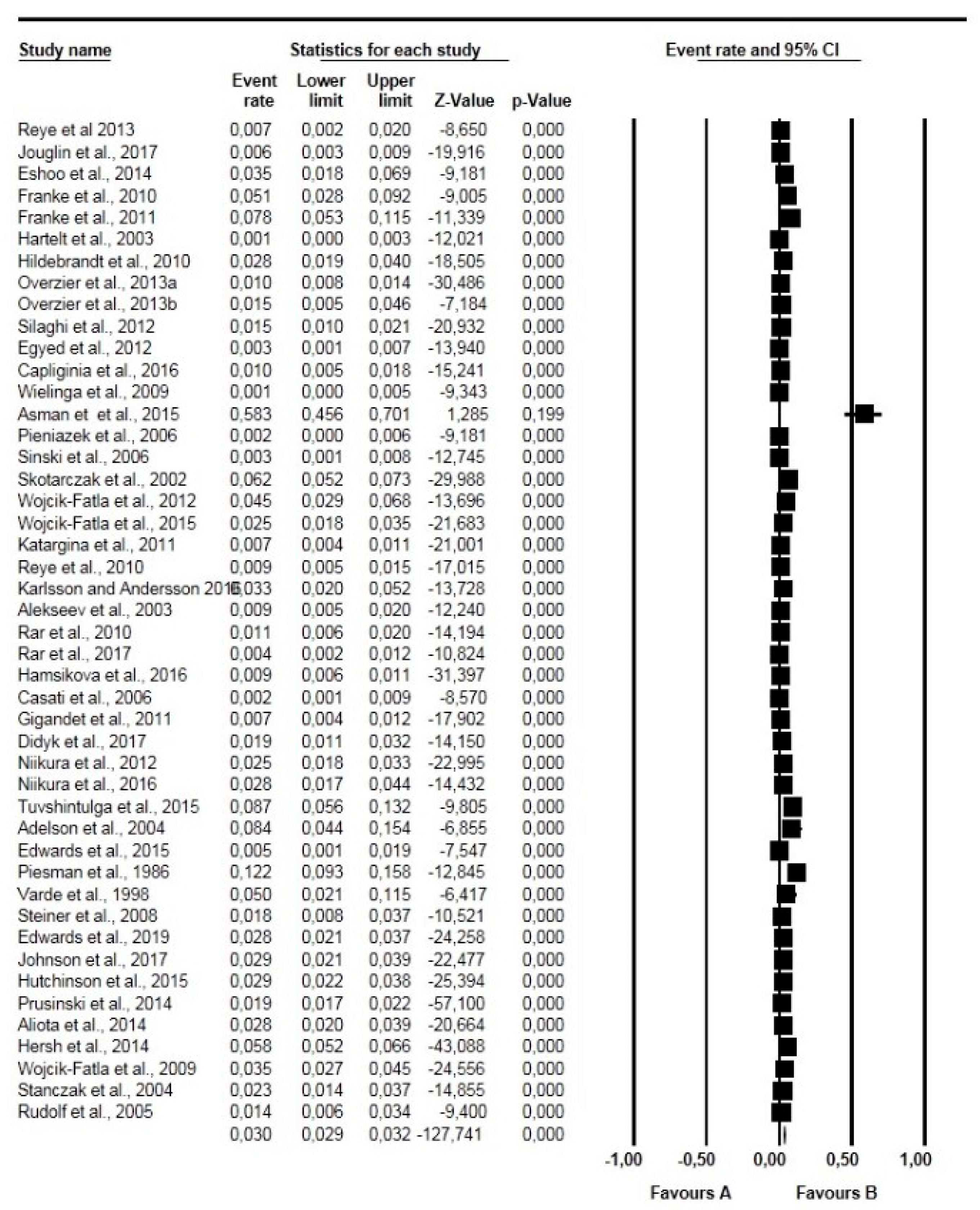
2.3. Pooling, Heterogeneity and Subgroup Analysis
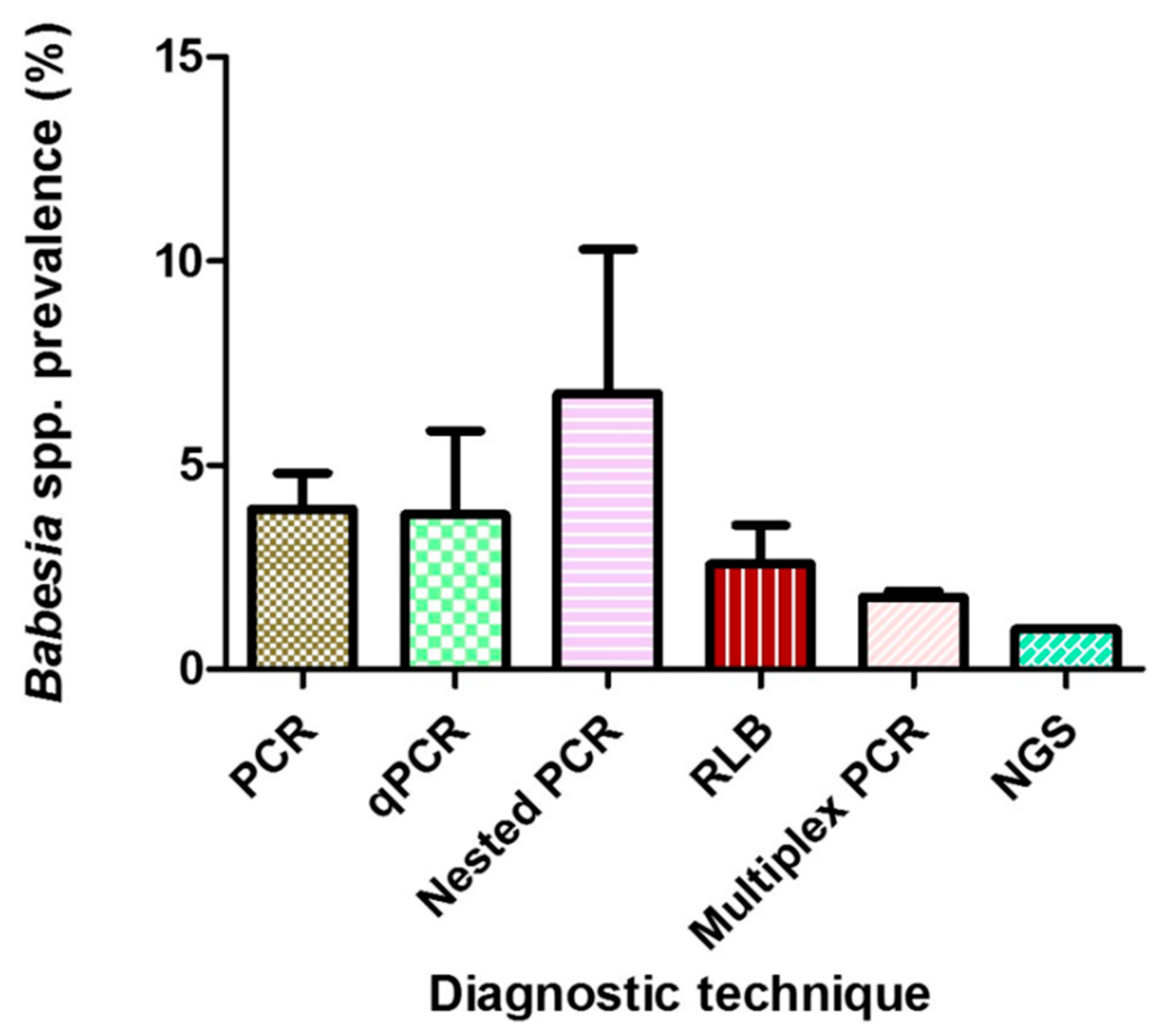
2.3.1. Prevalence Based on Tick Species, Life Stages, Sex, and Diagnostic Technique
2.3.2. Prevalence Based on Babesia Species, Region, and Sampling Periods
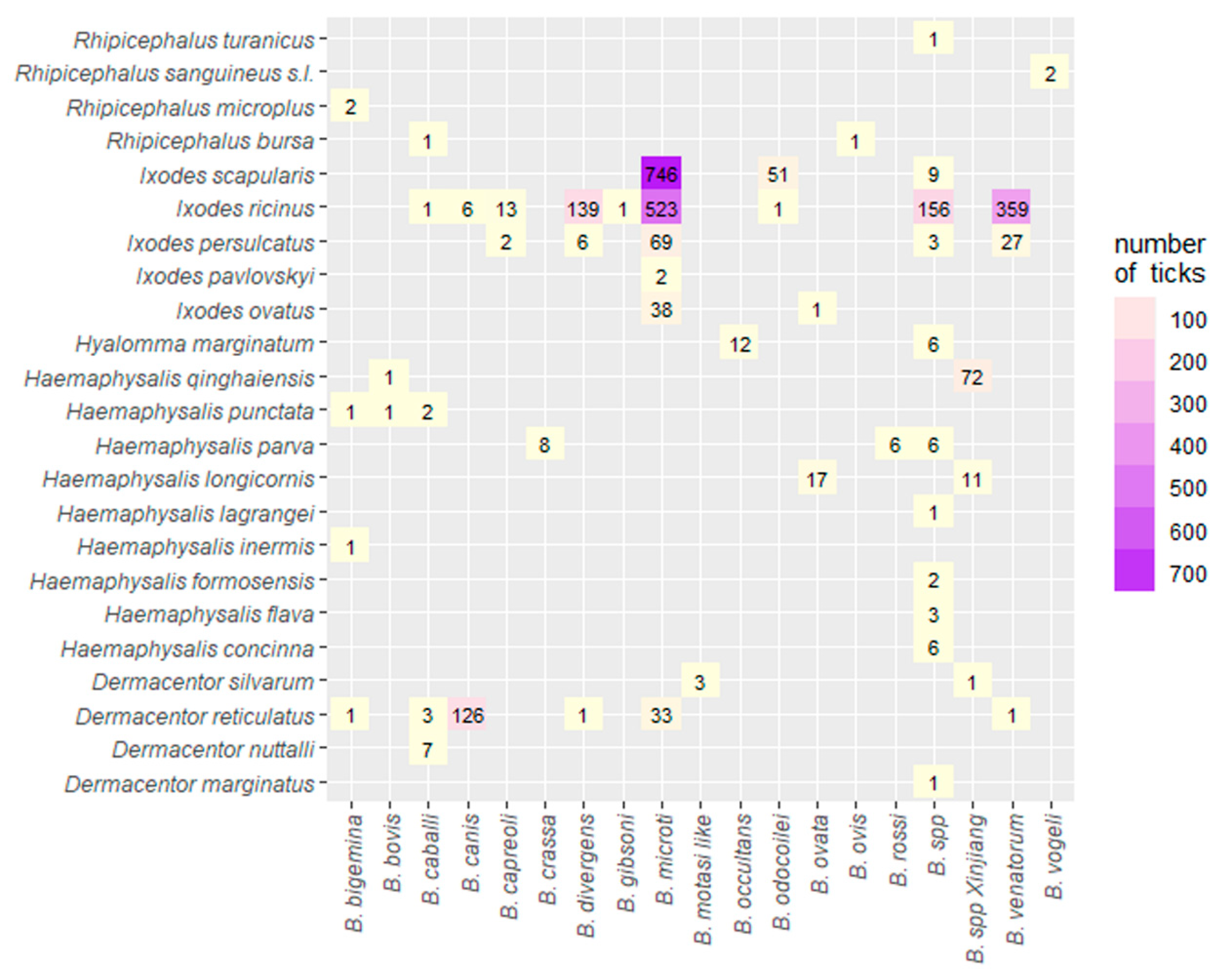
2.3.3. Species Diversity of Babesia within Different Tick Species
2.4. Spatial Distribution of Eligible Studies
2.5. Publication Bias
3. Discussion
3.1. Babesia Species in Ticks with Medical Importance
3.2. Babesia Species in Ticks with Veterinary Importance
3.3. Ticks as Vectors of Babesia Species
3.4. Association between Ticks and Babesia Including Other Factors
4. Material and Methods
4.1. Search Strategy
4.2. Inclusion and Exclusion Criteria
4.3. Data Cleaning
4.4. Data Extraction
4.5. Quality Assessment of Included Studies
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef]
- Blaschitz, M.; Narodoslavsky-Gfoller, M.; Kanzler, M.; Stanek, G.; Walochnik, J. Babesia species occurring in Austrian Ixodes ricinus ticks. Appl. Environ. Microbiol. 2008, 74, 4841–4846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uilenberg, G. Babesia--a historical overview. Vet. Parasitol. 2006, 138, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Homer, M.J.; Aguilar-Delfin, I.; Telford, S.R., 3rd; Krause, P.J.; Persing, D.H. Babesiosis. Clin. Microbiol. Rev. 2000, 13, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Babes, V. L’etiologie d’une enzootie des moutons, denommee Carceag en Roumanie. (The etiology of an enzootic disease of sheep, called Carceag, in Romania). C. R. Hebd. Acad. Sci. 1892, 115, 359–361. [Google Scholar]
- Smith, T.; Kilbourne, E.L. Investigation into the Nature Causation and Prevention of Southern Cattle Fever; US Department of Agriculture, Bureau of Animal Industry: Washington, DC, USA, 1893; Volume 1, pp. 1–311.
- Starcovici, C. Bemerkungen über den durch Babes entdeckten Blutparasiten und die durch denselben hervorgebrachten Krankheiten, die seuchenhafte Hämoglobinurie des Rindes (Babes), des Texasfieber (Th. Smith) und der Carceag der Schafe (Babes). Zbl. Bakt. I. Abt. 1893, 14, 11–18. [Google Scholar]
- Li, L.H.; Wang, J.Z.; Zhu, D.; Li, X.S.; Lu, Y.; Yin, S.Q.; Li, S.G.; Zhang, Y.; Zhou, X.N. Detection of novel piroplasmid species and Babesia microti and Theileria orientalis genotypes in hard ticks from Tengchong County, Southwest China. Parasitol. Res. 2020, 119, 1259–1269. [Google Scholar] [CrossRef]
- Skrabalo, Z.; Deanovic, Z. Piroplasmosis in man; report of a case. Doc. Med. Geogr. Trop. 1957, 9, 11–16. [Google Scholar] [PubMed]
- Hunfeld, K.P.; Hildebrandt, A.; Gray, J.S. Babesiosis: Recent insights into an ancient disease. Int. J. Parasitol. 2008, 38, 1219–1237. [Google Scholar] [CrossRef] [PubMed]
- Moritz, E.D.; Winton, C.S.; Tonnetti, L.; Townsend, R.L.; Berardi, V.P.; Hewins, M.E.; Weeks, K.E.; Dodd, R.Y.; Stramer, S.L. Screening for Babesia microti in the U.S. Blood Supply. N. Engl. J. Med. 2016, 375, 2236–2245. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.; Zintl, A.; Hildebrandt, A.; Hunfeld, K.P.; Weiss, L. Zoonotic babesiosis: Overview of the disease and novel aspects of pathogen identity. Ticks Tick Borne Dis. 2010, 1, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Guglielmone, A.A.; Robbins, R.G.; Apanaskevich, D.A.; Petney, T.N.; Estrada-Peña, A.; Horak, I. The Hard Ticks of the World (Acari, Ixodida, Ixodidae); Springer: New York, NY, USA, 2014; pp. 978–994. [Google Scholar]
- Gherman, C.M.; Mihalca, A.D.; Dumitrache, M.O.; Gyorke, A.; Oroian, I.; Sandor, M.; Cozma, V. CO2 flagging—An improved method for the collection of questing ticks. Parasit. Vectors 2012, 5, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petney, T.N.; Robbins, R.G.; Guglielmone, A.A.; Apanaskevich, D.A.; Estrada-Pena, A.; Horak, I.G.; Shao, R. A Look at the World of Ticks. In Progress in Parasitology; Springer: Berlin/Heidelberg, Germany, 2011; pp. 283–296. [Google Scholar]
- Leonovich, S.A. Ontogenesis of the questing behavior of hard ticks (Ixodidae). Entomol. Rev. 2015, 95, 795–804. [Google Scholar] [CrossRef]
- Silaghi, C.; Woll, D.; Hamel, D.; Pfister, K.; Mahling, M.; Pfeffer, M. Babesia spp. and Anaplasma phagocytophilum in questing ticks, ticks parasitizing rodents and the parasitized rodents-analyzing the host-pathogen-vector interface in a metropolitan area. Parasit Vectors 2012, 5, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yabsley, M.J.; Shock, B.C. Natural history of Zoonotic Babesia: Role of wildlife reservoirs. Int. J. Parasitol. Parasites Wildl. 2013, 2, 18–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, J.S.; Weiss, L.M. Babesia microti. In Emerging Protozoan Pathogens; Khan, N., Ed.; Taylor and Francis: Abingdon, UK, 2008; pp. 303–349. [Google Scholar]
- Jalovecka, M.; Sojka, D.; Ascencio, M.; Schnittger, L. Babesia Life Cycle—When Phylogeny Meets Biology. Trends Parasitol. 2019, 35, 356–368. [Google Scholar] [CrossRef]
- Hornok, S.; Kartali, K.; Takacs, N.; Hofmann-Lehmann, R. Uneven seasonal distribution of Babesia canis and its two 18S rDNA genotypes in questing Dermacentor reticulatus ticks in urban habitats. Ticks Tick Borne Dis. 2016, 7, 694–697. [Google Scholar] [CrossRef] [PubMed]
- Rar, V.A.; Fomenko, N.V.; Dobrotvorsky, A.K.; Livanova, N.N.; Rudakova, S.A.; Fedorov, E.G.; Astanin, V.B.; Morozova, O.V. Tickborne pathogen detection, Western Siberia, Russia. Emerg. Infect. Dis. 2005, 11, 1708–1715. [Google Scholar] [CrossRef]
- Schaarschmidt, D.; Gilli, U.; Gottstein, B.; Marreros, N.; Kuhnert, P.; Daeppen, J.A.; Rosenberg, G.; Hirt, D.; Frey, C.F. Questing Dermacentor reticulatus harbouring Babesia canis DNA associated with outbreaks of canine babesiosis in the Swiss Midlands. Ticks Tick Borne Dis. 2013, 4, 334–340. [Google Scholar] [CrossRef] [Green Version]
- Silaghi, C.; Weis, L.; Pfister, K. Dermacentor reticulatus and Babesia canis in Bavaria (Germany)-A Georeferenced Field Study with Digital Habitat Characterization. Pathogens 2020, 9, 541. [Google Scholar] [CrossRef]
- Sprong, H.; Fonville, M.; Docters van Leeuwen, A.; Devillers, E.; Ibanez-Justicia, A.; Stroo, A.; Hansford, K.; Cull, B.; Medlock, J.; Heyman, P.; et al. Detection of pathogens in Dermacentor reticulatus in northwestern Europe: Evaluation of a high-throughput array. Heliyon 2019, 5, e01270. [Google Scholar] [CrossRef] [Green Version]
- Milnes, E.L.; Thornton, G.; Leveille, A.N.; Delnatte, P.; Barta, J.R.; Smith, D.A.; Nemeth, N. Babesia odocoilei and zoonotic pathogens identified from Ixodes scapularis ticks in southern Ontario, Canada. Ticks Tick Borne Dis. 2019, 10, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Prusinski, M.A.; Kokas, J.E.; Hukey, K.T.; Kogut, S.J.; Lee, J.; Backenson, P.B. Prevalence of Borrelia burgdorferi (Spirochaetales: Spirochaetaceae), Anaplasma phagocytophilum (Rickettsiales: Anaplasmataceae), and Babesia microti (Piroplasmida: Babesiidae) in Ixodes scapularis (Acari: Ixodidae) collected from recreational lands in the Hudson Valley Region, New York State. J. Med. Entomol. 2014, 51, 226–236. [Google Scholar] [PubMed]
- Niu, Q.; Liu, Z.; Yang, J.; Gao, S.; Pan, Y.; Guan, G.; Luo, J.; Yin, H. Genetic characterization and molecular survey of Babesia sp. Xinjiang infection in small ruminants and ixodid ticks in China. Infect. Genet. Evol. 2017, 49, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, T.; Tattiyapong, M.; Okubo, K.; Suganuma, K.; Hayashida, K.; Igarashi, I.; Zakimi, S.; Matsumoto, K.; Inokuma, H.; Yokoyama, N. PCR detection of Babesia ovata from questing ticks in Japan. Ticks Tick Borne Dis. 2014, 5, 305–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamoto-Niikura, A.; Morikawa, S.; Hanaki, K.I.; Holman, P.J.; Ishihara, C. Ixodes persulcatus Ticks as Vectors for the Babesia microti U.S. Lineage in Japan. Appl. Environ. Microbiol. 2016, 82, 6624–6632. [Google Scholar] [CrossRef] [Green Version]
- Rar, V.; Livanova, N.; Tkachev, S.; Kaverina, G.; Tikunov, A.; Sabitova, Y.; Igolkina, Y.; Panov, V.; Livanov, S.; Fomenko, N.; et al. Detection and genetic characterization of a wide range of infectious agents in Ixodes pavlovskyi ticks in Western Siberia, Russia. Parasit. Vectors 2017, 10, 258. [Google Scholar] [CrossRef] [PubMed]
- Tuvshintulga, B.; Sivakumar, T.; Battsetseg, B.; Narantsatsaral, S.O.; Enkhtaivan, B.; Battur, B.; Hayashida, K.; Okubo, K.; Ishizaki, T.; Inoue, N.; et al. The PCR detection and phylogenetic characterization of Babesia microti in questing ticks in Mongolia. Parasitol. Int. 2015, 64, 527–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkmann, A.; Hekimoglu, O.; Dincer, E.; Hagedorn, P.; Nitsche, A.; Ergunay, K. A cross-sectional screening by next-generation sequencing reveals Rickettsia, Coxiella, Francisella, Borrelia, Babesia, Theileria and Hemolivia species in ticks from Anatolia. Parasit. Vectors 2019, 12, 26. [Google Scholar] [CrossRef] [Green Version]
- Reye, A.L.; Arinola, O.G.; Hubschen, J.M.; Muller, C.P. Pathogen prevalence in ticks collected from the vegetation and livestock in Nigeria. Appl. Environ. Microbiol. 2012, 78, 2562–2568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliota, M.T.; Dupuis, A.P., 2nd; Wilczek, M.P.; Peters, R.J.; Ostfeld, R.S.; Kramer, L.D. The prevalence of zoonotic tick-borne pathogens in Ixodes scapularis collected in the Hudson Valley, New York State. Vector Borne Zoonotic Dis. 2014, 14, 245–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franke, J.; Fritzsch, J.; Tomaso, H.; Straube, E.; Dorn, W.; Hildebrandt, A. Coexistence of pathogens in host-seeking and feeding ticks within a single natural habitat in Central Germany. Appl. Environ. Microbiol. 2010, 76, 6829–6836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamoto-Niikura, A.; Tsuji, M.; Qiang, W.; Nakao, M.; Hirata, H.; Ishihara, C. Detection of two zoonotic Babesia microti lineages, the Hobetsu and U.S. lineages, in two sympatric tick species, Ixodes ovatus and Ixodes persulcatus, respectively, in Japan. Appl. Environ. Microbiol. 2012, 78, 3424–3430. [Google Scholar] [CrossRef] [PubMed]
- Overzier, E.; Pfister, K.; Herb, I.; Mahling, M.; Bock, G., Jr.; Silaghi, C. Detection of tick-borne pathogens in roe deer (Capreolus capreolus), in questing ticks (Ixodes ricinus), and in ticks infesting roe deer in southern Germany. Ticks Tick Borne Dis. 2013, 4, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Lommano, E.; Bertaiola, L.; Dupasquier, C.; Gern, L. Infections and coinfections of questing Ixodes ricinus ticks by emerging zoonotic pathogens in Western Switzerland. Appl. Environ. Microbiol. 2012, 78, 4606–4612. [Google Scholar] [CrossRef] [Green Version]
- Reis, C.; Cote, M.; Paul, R.E.; Bonnet, S. Questing ticks in suburban forest are infected by at least six tick-borne pathogens. Vector Borne Zoonotic Dis. 2011, 11, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Jongejan, F.; Ringenier, M.; Putting, M.; Berger, L.; Burgers, S.; Kortekaas, R.; Lenssen, J.; van Roessel, M.; Wijnveld, M.; Madder, M. Novel foci of Dermacentor reticulatus ticks infected with Babesia canis and Babesia caballi in the Netherlands and in Belgium. Parasit. Vectors 2015, 8, 232. [Google Scholar] [CrossRef] [Green Version]
- Steiner, F.E.; Pinger, R.R.; Vann, C.N.; Abley, M.J.; Sullivan, B.; Grindle, N.; Clay, K.; Fuqua, C. Detection of Anaplasma phagocytophilum and Babesia odocoilei DNA in Ixodes scapularis (Acari: Ixodidae) collected in Indiana. J. Med. Entomol. 2006, 43, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Omar Abdallah, M.; Niu, Q.; Yu, P.; Guan, G.; Yang, J.; Chen, Z.; Liu, G.; Wei, Y.; Luo, J.; Yin, H. Identification of piroplasm infection in questing ticks by RLB: A broad range extension of tick-borne piroplasm in China? Parasitol. Res. 2016, 115, 2035–2044. [Google Scholar] [CrossRef]
- Garcia-Sanmartin, J.; Barandika, J.F.; Juste, R.A.; Garcia-Perez, A.L.; Hurtado, A. Distribution and molecular detection of Theileria and Babesia in questing ticks from northern Spain. Med. Vet. Entomol. 2008, 22, 318–325. [Google Scholar] [CrossRef]
- Battsetseg, B.; Xuan, X.N.; Ikadai, H.; Bautista, J.L.R.; Byambaa, B.; Boldbaatar, D.; Battur, B.; Battsetseg, G.; Batsukh, Z.; Igarashi, I.; et al. Detection of Babesia caballi and Babesia equi in Dermacentor nuttalli adult ticks. Int. J. Parasitol. 2001, 31, 384–386. [Google Scholar] [CrossRef]
- Overzier, E.; Pfister, K.; Thiel, C.; Herb, I.; Mahling, M.; Silaghi, C. Diversity of Babesia and Rickettsia species in questing Ixodes ricinus: A longitudinal study in urban, pasture, and natural habitats. Vector Borne Zoonotic Dis. 2013, 13, 559–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schorn, S.; Pfister, K.; Reulen, H.; Mahling, M.; Silaghi, C. Occurrence of Babesia spp., Rickettsia spp. and Bartonella spp. in Ixodes ricinus in Bavarian public parks, Germany. Parasit. Vectors 2011, 4, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reye, A.L.; Stegniy, V.; Mishaeva, N.P.; Velhin, S.; Hubschen, J.M.; Ignatyev, G.; Muller, C.P. Prevalence of tick-borne pathogens in Ixodes ricinus and Dermacentor reticulatus ticks from different geographical locations in Belarus. PLoS ONE 2013, 8, e54476. [Google Scholar]
- Klitgaard, K.; Kjaer, L.J.; Isbrand, A.; Hansen, M.F.; Bodker, R. Multiple infections in questing nymphs and adult female Ixodes ricinus ticks collected in a recreational forest in Denmark. Ticks Tick Borne Dis. 2019, 10, 1060–1065. [Google Scholar] [CrossRef]
- Sormunen, J.J.; Andersson, T.; Aspi, J.; Back, J.; Cederberg, T.; Haavisto, N.; Halonen, H.; Hanninen, J.; Inkinen, J.; Kulha, N.; et al. Monitoring of ticks and tick-borne pathogens through a nationwide research station network in Finland. Ticks Tick Borne Dis. 2020, 11, 101449. [Google Scholar] [CrossRef] [PubMed]
- Sormunen, J.J.; Klemola, T.; Hanninen, J.; Makela, S.; Vuorinen, I.; Penttinen, R.; Saaksjarvi, I.E.; Vesterinen, E.J. The importance of study duration and spatial scale in pathogen detection-evidence from a tick-infested island. Emerg. Microbes Infect. 2018, 7, 189. [Google Scholar] [CrossRef]
- Bonnet, S.; de la Fuente, J.; Nicollet, P.; Liu, X.; Madani, N.; Blanchard, B.; Maingourd, C.; Alongi, A.; Torina, A.; Fernandez de Mera, I.G.; et al. Prevalence of tick-borne pathogens in adult Dermacentor spp. ticks from nine collection sites in France. Vector Borne Zoonotic Dis. 2013, 13, 226–236. [Google Scholar] [CrossRef]
- Cotte, V.; Bonnet, S.; Cote, M.; Vayssier-Taussat, M. Prevalence of five pathogenic agents in questing Ixodes ricinus ticks from western France. Vector Borne Zoonotic Dis. 2010, 10, 723–730. [Google Scholar] [CrossRef]
- Halos, L.; Jamal, T.; Maillard, R.; Beugnet, F.; Le Menach, A.; Boulouis, H.J.; Vayssier-Taussat, M. Evidence of Bartonella sp. in questing adult and nymphal Ixodes ricinus ticks from France and co-infection with Borrelia burgdorferi sensu lato and Babesia sp. Vet. Res. 2005, 36, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Jouglin, M.; Perez, G.; Butet, A.; Malandrin, L.; Bastian, S. Low prevalence of zoonotic Babesia in small mammals and Ixodes ricinus in Brittany, France. Vet. Parasitol. 2017, 238, 58–60. [Google Scholar] [CrossRef]
- Lejal, E.; Moutailler, S.; Simo, L.; Vayssier-Taussat, M.; Pollet, T. Tick-borne pathogen detection in midgut and salivary glands of adult Ixodes ricinus. Parasit. Vectors 2019, 12, 152. [Google Scholar] [CrossRef] [Green Version]
- Eshoo, M.W.; Crowder, C.D.; Carolan, H.E.; Rounds, M.A.; Ecker, D.J.; Haag, H.; Mothes, B.; Nolte, O. Broad-range survey of tick-borne pathogens in Southern Germany reveals a high prevalence of Babesia microti and a diversity of other tick-borne pathogens. Vector Borne Zoonotic Dis. 2014, 14, 584–591. [Google Scholar] [CrossRef]
- Franke, J.; Hildebrandt, A.; Meier, F.; Straube, E.; Dorn, W. Prevalence of Lyme disease agents and several emerging pathogens in questing ticks from the German Baltic coast. J. Med. Entomol. 2011, 48, 441–444. [Google Scholar] [CrossRef]
- Hartelt, K.; Oehme, R.; Frank, H.; Brockmann, S.O.; Hassler, D.; Kimmig, P. Pathogens and symbionts in ticks: Prevalence of Anaplasma phagocytophilum (Ehrlichia sp.), Wolbachia sp., Rickettsia sp., and Babesia sp in Southern Germany. Int. J. Med. Microbiol. 2004, 293, 86–92. [Google Scholar] [CrossRef]
- Hildebrandt, A.; Pauliks, K.; Sachse, S.; Straube, E. Coexistence of Borrelia spp. and Babesia spp. in Ixodes ricinus ticks in Middle Germany. Vector Borne Zoonotic Dis. 2010, 10, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, A.; Fritzsch, J.; Franke, J.; Sachse, S.; Dorn, W.; Straube, E. Co-circulation of emerging tick-borne pathogens in Middle Germany. Vector Borne Zoonotic Dis. 2011, 11, 533–537. [Google Scholar] [CrossRef]
- Kohn, M.; Krucken, J.; McKay-Demeler, J.; Pachnicke, S.; Krieger, K.; von Samson-Himmelstjerna, G. Dermacentor reticulatus in Berlin/Brandenburg (Germany): Activity patterns and associated pathogens. Ticks Tick Borne Dis. 2019, 10, 191–206. [Google Scholar] [CrossRef]
- Egyed, L.; Elo, P.; Sreter-Lancz, Z.; Szell, Z.; Balogh, Z.; Sreter, T. Seasonal activity and tick-borne pathogen infection rates of Ixodes ricinus ticks in Hungary. Ticks Tick Borne Dis. 2012, 3, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Capelli, G.; Ravagnan, S.; Montarsi, F.; Ciocchetta, S.; Cazzin, S.; Porcellato, E.; Babiker, A.M.; Cassini, R.; Salviato, A.; Cattoli, G.; et al. Occurrence and identification of risk areas of Ixodes ricinus-borne pathogens: A cost-effectiveness analysis in north-eastern Italy. Parasit. Vector 2012, 5, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassini, R.; Bonoli, C.; Montarsi, F.; Tessarin, C.; Marcer, F.; Galuppi, R. Detection of Babesia EU1 in Ixodes ricinus ticks in northern Italy. Vet. Parasitol. 2010, 171, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Floris, R.; Cecco, P.; Mignozzi, K.; Boemo, B.; Cinco, M. First detection of Babesia EU1 and Babesia divergens-like in Ixodes ricinus ticks in north-eastern Italy. Parassitologia 2009, 51, 23–28. [Google Scholar]
- Piccolin, G.; Benedetti, G.; Doglioni, C.; Lorenzato, C.; Mancuso, S.; Papa, N.; Pttton, L.; Ramon, M.C.; Zasio, C.; Bertiato, G. A study of the presence of Borrelia burgdorferi, Anaplasma (previously Ehrlichia) phagocytophilum, Rickettsia, and Babesia in Ixodes ricinus collected within the territory of Belluno, Italy. Vector-Borne Zoonot. 2006, 6, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Capligina, V.; Berzina, I.; Bormane, A.; Salmane, I.; Vilks, K.; Kazarina, A.; Bandere, D.; Baumanis, V.; Ranka, R. Prevalence and phylogenetic analysis of Babesia spp. in Ixodes ricinus and Ixodes persulcatus ticks in Latvia. Exp. Appl. Acarol. 2016, 68, 325–336. [Google Scholar] [CrossRef]
- Radzijevskaja, J.; Mardosaite-Busaitiene, D.; Aleksandraviciene, A.; Paulauskas, A. Investigation of Babesia spp. in sympatric populations of Dermacentor reticulatus and Ixodes ricinus ticks in Lithuania and Latvia. Ticks Tick-Borne Dis. 2018, 9, 270–274. [Google Scholar] [CrossRef]
- Oines, O.; Radzijevskaja, J.; Paulauskas, A.; Rosef, O. Prevalence and diversity of Babesia spp. in questing Ixodes ricinus ticks from Norway. Parasit. Vectors 2012, 5, 156. [Google Scholar] [CrossRef] [Green Version]
- Radzijevskaja, J.; Paulauskas, A.; Rosef, O. Prevalence of Anaplasma phagocytophilum and Babesia divergens in Ixodes ricinus ticks from Lithuania and Norway. Int. J. Med. Microbiol. 2008, 298, 218–221. [Google Scholar] [CrossRef] [Green Version]
- Žygutienė, M.; Alekseev, A.; Dubinina, H.; Kazlauskienė, R. Evidence for a risk of tick-borne infection in the city parks of Vilnius, Lithuania. EKOLOGIJA 2008, 54, 40–43. [Google Scholar] [CrossRef]
- Wielinga, P.R.; Fonville, M.; Sprong, H.; Gaasenbeek, C.; Borgsteede, F.; van der Giessen, J.W.B. Persistent Detection of Babesia EU1 and Babesia microti in Ixodes ricinus in The Netherlands During a 5-Year Surveillance: 2003-2007. Vector-Borne Zoonot. 2009, 9, 119–121. [Google Scholar] [CrossRef]
- Asman, M.; Solarz, K.; Cuber, P.; Gasior, T.; Szilman, P.; Szilman, E.; Tondas, E.; Matzullok, A.; Kusion, N.; Florek, K. Detection of protozoans Babesia microti and Toxoplasma gondii and their co-existence in ticks (Acari: Ixodida) collected in Tarnogorski district (Upper Silesia, Poland). Ann. Agric. Environ. Med. 2015, 22, 80–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cieniuch, S.; Stanczak, J.; Ruczaj, A. The First Detection of Babesia EU1 and Babesia canis canis in Ixodes ricinus Ticks (Acari, Ixodidae) Collected in Urban and Rural Areas in Northern Poland. Pol. J. Microbiol. 2009, 58, 231–236. [Google Scholar]
- Karbowiak, G.; Vichova, B.; Slivinska, K.; Werszko, J.; Didyk, J.; Pet’ko, B.; Stanko, M.; Akimov, I. The infection of questing Dermacentor reticulatus ticks with Babesia canis and Anaplasma phagocytophilum in the Chernobyl exclusion zone. Vet. Parasitol. 2014, 204, 372–375. [Google Scholar] [CrossRef] [PubMed]
- Pieniazek, N.; Sawczuk, M.; Skotarczak, B. Molecular identification of Babesia parasites isolated from Ixodes ricinus ticks collected in northwestern Poland. J. Parasitol. 2006, 92, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Sinski, E.; Bajer, A.; Welc, R.; Pawelczyk, A.; Ogrzewalska, M.; Behnke, J.M. Babesia microti: Prevalence in wild rodents and Ixodes ricinus ticks from the Mazury Lakes District of north-eastern Poland. Int. J. Med. Microbiol. 2006, 296, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Skotarczak, B.; Wodecka, B.; Cichocka, A. Coexistence DNA of Borrelia burgdorferi sensu lato and Babesia microti in Ixodes ricinus ticks from north-western Poland. Ann. Agric. Environ. Med. 2002, 9, 25–28. [Google Scholar]
- Stanczak, J.; Cieniuch, S.; Lass, A.; Biernat, B.; Racewicz, M. Detection and quantification of Anaplasma phagocytophilum and Babesia spp. in Ixodes ricinus ticks from urban and rural environment, northern Poland, by real-time polymerase chain reaction. Exp. Appl. Acarol. 2015, 66, 63–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welc-Faleciak, R.; Bajer, A.; Paziewska-Harris, A.; Baumann-Popczyk, A.; Sinski, E. Diversity of Babesia in Ixodes ricinus ticks in Poland. Adv. Med. Sci. Pol. 2012, 57, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Wojcik-Fatla, A.; Bartosik, K.; Buczek, A.; Dutkiewicz, J. Babesia microti in adult Dermacentor reticulatus ticks from Eastern Poland. Vector-Borne Zoonot. 2012, 12, 841–843. [Google Scholar] [CrossRef] [PubMed]
- Wojcik-Fatla, A.; Zajac, V.; Sawczyn, A.; Cisak, E.; Dutkiewicz, J. Babesia spp. in questing ticks from eastern Poland: Prevalence and species diversity. Parasitol. Res. 2015, 114, 3111–3116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zajac, V.; Wojcik-Fatla, A.; Sawczyn, A.; Cisak, E.; Sroka, J.; Kloc, A.; Zajac, Z.; Buczek, A.; Dutkiewicz, J.; Bartosik, K. Prevalence of infections and co-infections with 6 pathogens in Dermacentor reticulatus ticks collected in eastern Poland. Ann. Agric. Environ. Med. 2017, 24, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Wojcik-Fatla, A.; Szymanska, J.; Wdowiak, L.; Buczek, A.; Dutkiewicz, J. Coincidence of three pathogens (Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum and Babesia microti) in Ixodes ricinus ticks in the Lublin Macroregion. Ann. Agric. Environ. Med. 2009, 16, 151–158. [Google Scholar] [PubMed]
- Stanczak, J.; Gabre, R.M.; Kruminis-Lozowska, W.; Racewicz, M.; Kubica-Biernat, B. Ixodes ricinus as a vector of Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum and Babesia microti in urban and suburban forests. Ann. Agric. Environ. Med. 2004, 11, 109–114. [Google Scholar] [PubMed]
- Katargina, O.; Geller, J.; Vasilenko, V.; Kuznetsova, T.; Jarvekulg, L.; Vene, S.; Lundkvist, A.; Golovljova, I. Detection and characterization of Babesia species in Ixodes ticks in Estonia. Vector Borne Zoonotic Dis. 2011, 11, 923–928. [Google Scholar] [CrossRef]
- Maia, C.; Ferreira, A.; Nunes, M.; Vieira, M.L.; Campino, L.; Cardoso, L. Molecular detection of bacterial and parasitic pathogens in hard ticks from Portugal. Ticks Tick Borne Dis. 2014, 5, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Santos-Silva, M.M.; Melo, P.; Santos, N.; Antunes, S.; Duarte, L.R.; Ferrolho, J.; Milhano, N.; Santos, P.T.; Domingos, A.; Santos, A.S. PCR screening of tick-borne agents in sensitive conservation areas, Southeast Portugal. Mol. Cell. Probes 2017, 31, 42–45. [Google Scholar] [CrossRef]
- Reye, A.L.; Hubschen, J.M.; Sausy, A.; Muller, C.P. Prevalence and seasonality of tick-borne pathogens in questing Ixodes ricinus ticks from Luxembourg. Appl. Environ. Microbiol. 2010, 76, 2923–2931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ionita, M.; Mitrea, I.L.; Pfister, K.; Hamel, D.; Silaghi, C. Molecular evidence for bacterial and protozoan pathogens in hard ticks from Romania. Vet. Parasitol. 2013, 196, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, M.E.; Andersson, M.O. Babesia species in questing Ixodes ricinus, Sweden. Ticks Tick Borne Dis. 2016, 7, 10–12. [Google Scholar] [CrossRef] [PubMed]
- Jaenson, T.G.T.; Wilhelmsson, P. First records of tick-borne pathogens in populations of the taiga tick Ixodes persulcatus in Sweden. Parasit. Vectors 2019, 12, 559. [Google Scholar] [CrossRef] [Green Version]
- Alekseev, A.N.; Semenov, A.V.; Dubinina, H.V. Evidence of Babesia microti infection in multi-infected Ixodes persulcatus ticks in Russia. Exp. Appl. Acarol. 2003, 29, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Movila, A.; Dubinina, H.V.; Sitnicova, N.; Bespyatova, L.; Uspenskaia, I.; Efremova, G.; Toderas, I.; Alekseev, A.N. Comparison of tick-borne microorganism communities in Ixodes spp. of the Ixodes ricinus species complex at distinct geographical regions. Exp. Appl. Acarol. 2014, 63, 65–76. [Google Scholar] [CrossRef]
- Rar, V.A.; Epikhina, T.I.; Livanova, N.N.; Panov, V.V. Genetic diversity of Babesia in Ixodes persulcatus and small mammals from North Ural and West Siberia, Russia. Parasitology 2011, 138, 175–182. [Google Scholar] [CrossRef]
- Duh, D.; Slovak, M.; Saksida, A.; Stramsek, K.; Petrovec, M.; Avsic-Zupanc, T. Molecular detection of Babesia canis in Dermacentor reticulatus ticks collected in Slovakia. Biologia 2006, 61, 231–233. [Google Scholar] [CrossRef]
- Hamsikova, Z.; Kazimirova, M.; Harustiakova, D.; Mahrikova, L.; Slovak, M.; Berthova, L.; Kocianova, E.; Schnittger, L. Babesia spp. in ticks and wildlife in different habitat types of Slovakia. Parasit. Vectors 2016, 9, 292. [Google Scholar] [CrossRef] [Green Version]
- Svehlova, A.; Berthova, L.; Sallay, B.; Boldis, V.; Sparagano, O.A.; Spitalska, E. Sympatric occurrence of Ixodes ricinus, Dermacentor reticulatus and Haemaphysalis concinna ticks and Rickettsia and Babesia species in Slovakia. Ticks Tick Borne Dis. 2014, 5, 600–605. [Google Scholar] [CrossRef]
- Duh, D.; Petrovec, M.; Avsic-Zupanc, T. Diversity of Babesia infecting European sheep ticks (Ixodes ricinus). J. Clin. Microbiol. 2001, 39, 3395–3397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudolf, I.; Golovchenko, M.; Sikutova, S.; Rudenko, N.; Grubhoffer, L.; Hubalek, Z. Babesia microti (Piroplasmida: Babesiidae) in nymphal Ixodes ricinus (Acari: Ixodidae) in the Czech Republic. Folia. Parasitol. (Praha) 2005, 52, 274–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venclikova, K.; Mendel, J.; Betasova, L.; Blazejova, H.; Jedlickova, P.; Strakova, P.; Hubalek, Z.; Rudolf, I. Neglected tick-borne pathogens in the Czech Republic, 2011–2014. Ticks Tick Borne Dis. 2016, 7, 107–112. [Google Scholar] [CrossRef]
- Cochez, C.; Lempereur, L.; Madder, M.; Claerebout, E.; Simons, L.; De Wilde, N.; Linden, A.; Saegerman, C.; Heyman, P.; Losson, B. Foci report on indigenous Dermacentor reticulatus populations in Belgium and a preliminary study of associated babesiosis pathogens. Med. Vet. Entomol. 2012, 26, 355–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casati, S.; Sager, H.; Gern, L.; Piffaretti, J.C. Presence of potentially pathogenic Babesia sp. for human in Ixodes ricinus in Switzerland. Ann. Agric. Environ. Med. 2006, 13, 65–70. [Google Scholar]
- Gigandet, L.; Stauffer, E.; Douet, V.; Rais, O.; Moret, J.; Gern, L. Prevalence of three zoonotic Babesia species in Ixodes ricinus (Linne, 1758) nymphs in a suburban forest in Switzerland. Vector Borne Zoonotic Dis. 2011, 11, 363–366. [Google Scholar] [CrossRef]
- Oechslin, C.P.; Heutschi, D.; Lenz, N.; Tischhauser, W.; Peter, O.; Rais, O.; Beuret, C.M.; Leib, S.L.; Bankoul, S.; Ackermann-Gaumann, R. Prevalence of tick-borne pathogens in questing Ixodes ricinus ticks in urban and suburban areas of Switzerland. Parasit. Vectors 2017, 10, 558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Didyk, Y.M.; Blanarova, L.; Pogrebnyak, S.; Akimov, I.; Petko, B.; Vichova, B. Emergence of tick-borne pathogens (Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum, Ricketsia raoultii and Babesia microti) in the Kyiv urban parks, Ukraine. Ticks Tick Borne Dis. 2017, 8, 219–225. [Google Scholar] [CrossRef]
- Orkun, O.; Cakmak, A.; Nalbantoglu, S.; Karaer, Z. Turkey tick news: A molecular investigation into the presence of tick-borne pathogens in host-seeking ticks in Anatolia; Initial evidence of putative vectors and pathogens, and footsteps of a secretly rising vector tick, Haemaphysalis parva. Ticks Tick Borne Dis. 2020, 11, 101373. [Google Scholar] [CrossRef]
- Harrus, S.; Perlman-Avrahami, A.; Mumcuoglu, K.Y.; Morick, D.; Eyal, O.; Baneth, G. Molecular detection of Ehrlichia canis, Anaplasma bovis, Anaplasma platys, Candidatus Midichloria mitochondrii and Babesia canis vogeli in ticks from Israel. Clin. Microbiol. Infect. 2011, 17, 459–463. [Google Scholar] [CrossRef] [Green Version]
- Masatani, T.; Hayashi, K.; Andoh, M.; Tateno, M.; Endo, Y.; Asada, M.; Kusakisako, K.; Tanaka, T.; Gokuden, M.; Hozumi, N.; et al. Detection and molecular characterization of Babesia, Theileria, and Hepatozoon species in hard ticks collected from Kagoshima, the southern region in Japan. Ticks Tick Borne Dis. 2017, 8, 581–587. [Google Scholar] [CrossRef]
- Karnath, C.; Obiegala, A.; Speck, S.; Essbauer, S.; Derschum, H.; Scholz, H.; Kiefer, D.; Tserennorov, D.; Dashdavaa, O.; Tsogbadrakh, N.; et al. Detection of Babesia venatorum, Anaplasma phagocytophilum and Candidatus Neoehrlichia mikurensis in Ixodes persulcatus ticks from Mongolia. Ticks Tick-Borne Dis. 2016, 7, 357–360. [Google Scholar] [CrossRef]
- Wattanamethanont, J.; Kaewthamasorn, M.; Tiawsirisup, S. Natural infection of questing ixodid ticks with protozoa and bacteria in Chonburi Province, Thailand. Ticks Tick Borne Dis. 2018, 9, 749–758. [Google Scholar] [CrossRef]
- Adelson, M.E.; Rao, R.V.; Tilton, R.C.; Cabets, K.; Eskow, E.; Fein, L.; Occi, J.L.; Mordechai, E. Prevalence of Borrelia burgdorferi, Bartonella spp., Babesia microti, and Anaplasma phagocytophila in Ixodes scapularis ticks collected in Northern New Jersey. J. Clin. Microbiol. 2004, 42, 2799–2801. [Google Scholar] [CrossRef] [Green Version]
- Edwards, M.J.; Barbalato, L.A.; Makkapati, A.; Pham, K.D.; Bugbee, L.M. Relatively low prevalence of Babesia microti and Anaplasma phagocytophilum in Ixodes scapularis ticks collected in the Lehigh Valley region of eastern Pennsylvania. Ticks Tick Borne Dis. 2015, 6, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Piesman, J.; Mather, T.N.; Telford, S.R., 3rd; Spielman, A. Concurrent Borrelia burgdorferi and Babesia microti infection in nymphal Ixodes dammini. J. Clin. Microbiol. 1986, 24, 446–447. [Google Scholar] [CrossRef] [Green Version]
- Varde, S.; Beckley, J.; Schwartz, I. Prevalence of tick-borne pathogens in Ixodes scapularis in a rural New Jersey County. Emerg. Infect. Dis. 1998, 4, 97–99. [Google Scholar] [CrossRef]
- Steiner, F.E.; Pinger, R.R.; Vann, C.N.; Grindle, N.; Civitello, D.; Clay, K.; Fuqua, C. Infection and co-infection rates of Anaplasma phagocytophilum variants, Babesia spp., Borrelia burgdorferi, and the rickettsial endosymbiont in Ixodes scapularis (Acari: Ixodidae) from sites in Indiana, Maine, Pennsylvania, and Wisconsin. J. Med. Entomol. 2008, 45, 289–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, M.J.; Russell, J.C.; Davidson, E.N.; Yanushefski, T.J.; Fleischman, B.L.; Heist, R.O.; Leep-Lazar, J.G.; Stuppi, S.L.; Esposito, R.A.; Suppan, L.M. A 4-Yr Survey of the Range of Ticks and Tick-Borne Pathogens in the Lehigh Valley Region of Eastern Pennsylvania. J. Med. Entomol. 2019, 56, 1122–1134. [Google Scholar] [CrossRef]
- Russart, N.M.; Dougherty, M.W.; Vaughan, J.A. Survey of ticks (Acari: Ixodidae) and tick-borne pathogens in North Dakota. J. Med. Entomol. 2014, 51, 1087–1090. [Google Scholar] [CrossRef]
- Hutchinson, M.L.; Strohecker, M.D.; Simmons, T.W.; Kyle, A.D.; Helwig, M.W. Prevalence Rates of Borrelia burgdorferi (Spirochaetales: Spirochaetaceae), Anaplasma phagocytophilum (Rickettsiales: Anaplasmataceae), and Babesia microti (Piroplasmida: Babesiidae) in host-seeking Ixodes scapularis (Acari: Ixodidae) from Pennsylvania. J. Med. Entomol. 2015, 52, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Hersh, M.H.; Ostfeld, R.S.; McHenry, D.J.; Tibbetts, M.; Brunner, J.L.; Killilea, M.E.; LoGiudice, K.; Schmidt, K.A.; Keesing, F. Co-infection of blacklegged ticks with Babesia microti and Borrelia burgdorferi is higher than expected and acquired from small mammal hosts. PLoS ONE 2014, 9, e99348. [Google Scholar] [CrossRef]
- Swei, A.; O’Connor, K.E.; Couper, L.I.; Thekkiniath, J.; Conrad, P.A.; Padgett, K.A.; Burns, J.; Yoshimizu, M.H.; Gonzales, B.; Munk, B.; et al. Evidence for transmission of the zoonotic apicomplexan parasite Babesia duncani by the tick Dermacentor albipictus. Int. J. Parasitol. 2019, 49, 95–103. [Google Scholar] [CrossRef]
- Young, C.; Chawla, A.; Berardi, V.; Padbury, J.; Skowron, G.; Krause, P.J.; Containment, B.T.I. Preventing transfusion-transmitted babesiosis: Preliminary experience of the first laboratory-based blood donor screening program. Transfusion 2012, 52, 1523–1529. [Google Scholar] [CrossRef]
- Guan, G.; Ma, M.; Moreau, E.; Liu, J.; Lu, B.; Bai, Q.; Luo, J.; Jorgensen, W.; Chauvin, A.; Yin, H. A new ovine Babesia species transmitted by Hyalomma anatolicum anatolicum. Exp. Parasitol. 2009, 122, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Bai, Q.; Liu, G.Y.; Zhang, L.; Zhou, J.Y. Studies on the isolation and preservation of a single species of bovine haematocytozoon: The finding and isolation of Babesia ovata in China. Chin. J. Vet. Med. 1990, 16, 2–4. [Google Scholar]
- Aktas, M. A survey of ixodid tick species and molecular identification of tick-borne pathogens. Vet. Parasitol. 2014, 200, 276–283. [Google Scholar] [CrossRef]
- Hoogstraal, H. The epidemiology of tick-borne Crimean-Congo hemorrhagic fever in Asia, Europe, and Africa. J. Med. Entomol. 1979, 15, 307–417. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Pena, A.; George, J.C.; Golovljova, I.; Jaenson, T.G.; Jensen, J.K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasit. Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubalek, Z.; Foldvari, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Spitalska, E.; et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: New hazards and relevance for public health. Front. Public Health 2014, 2, 251. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.; Eze, J.I.; Gilbert, L.; Wint, G.R.W.; Gunn, G.; Macrae, A.; Medlock, J.M.; Auty, H. Using imperfect data in predictive mapping of vectors: A regional example of Ixodes ricinus distribution. Parasit. Vector 2019, 12, 536. [Google Scholar] [CrossRef] [PubMed]
- Livanova, N.N.; Tikunov, A.Y.; Kurilshikov, A.M.; Livanov, S.G.; Fomenko, N.V.; Taranenko, D.E.; Kvashnina, A.E.; Tikunova, N.V. Genetic diversity of Ixodes pavlovskyi and I. persulcatus (Acari: Ixodidae) from the sympatric zone in the south of Western Siberia and Kazakhstan. Exp. Appl. Acarol. 2015, 67, 441–456. [Google Scholar] [CrossRef] [PubMed]
- Friedhoff, K.T. Transmission of Babesia. In Babesiosis of Domestic Animals and Man; Ristic, M., Ed.; CRC Press: Boca Raton, FL, USA, 1988. [Google Scholar] [CrossRef]
- Eisen, R.J.; Eisen, L. The Blacklegged Tick, Ixodes scapularis: An increasing public health concern. Trends Parasitol. 2018, 34, 295–309. [Google Scholar] [CrossRef]
- Gray, J.S.; Estrada-Pena, A.; Zintl, A. Vectors of Babesiosis. Annu. Rev. Entomol. 2019, 64, 149–165. [Google Scholar] [CrossRef]
- Strnad, M.; Honig, V.; Ruzek, D.; Grubhoffer, L.; Rego, R.O.M. Europe-Wide Meta-Analysis of Borrelia burgdorferi sensu lato prevalence in questing Ixodes ricinus Ticks. Appl. Environ. Microbiol. 2017, 83, e00609-17. [Google Scholar] [CrossRef] [Green Version]
- Moher, D.; Shamseer, L.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A.; Group, P.-P. Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015 statement. Syst. Rev. 2015, 4, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munn, Z.; Moola, S.; Lisy, K.; Riitano, D.; Tufanaru, C. Methodological guidance for systematic reviews of observational epidemiological studies reporting prevalence and cumulative incidence data. Int. J. Evid. Based Healthc. 2015, 13, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.T.; Thompson, S.G. Quantifying heterogeneity in a meta-analysis. Stat. Med. 2002, 21, 1539–1558. [Google Scholar] [CrossRef] [PubMed]
- Begg, C.B.; Mazumdar, M. Operating characteristics of a rank correlation test for publication bias. Biometrics 1994, 50, 1088–1101. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Year | Country | Continent | Molecular Technique | Sample Size | Cases | MIR | JBI QAS | Study Ref. |
|---|---|---|---|---|---|---|---|---|
| 2005 | Austria | Europe | PCR | 864 | 441 | 51.04 | 5 | [2] |
| 2009 | Belarus | Europe | PCR | 453 | 5 | 1.10 | 7 | [48] |
| 2016–2017 | Denmark | Europe | qPCR | 1013 | 5 | 0.49 | 7 | [49] |
| 2015 | Finland | Europe | qPCR/PCR | 515 | 6 | 1.17 | 7 | [50] |
| 2012–2017 | Finland | Europe | qPCR | 7070 | 41 | 0.58 | 8 | [51] |
| 2009 | France | Europe | PCR/RLB | 495 | 4 | 0.81 | 7 | [52] |
| 2006–2007 | France | Europe | PCR | 572 | 35 | 6.12 | 8 | [53] |
| 2002 | France | Europe | PCR | 92 | 19 | 20.65 | 8 | [54] |
| 2012–2013 | France | Europe | PCR | 2620 | 15 | 0.57 | 8 | [55] |
| 2017 | France | Europe | qPCR | 60 | 8 | 13.33 | 8 | [56] |
| 2008 | France | Europe | PCR | 558 | 6 | 1.08 | 8 | [40] |
| 2009 | Germany | Europe | PCR | 226 | 8 | 3.54 | 8 | [57] |
| 2007 | Germany | Europe | PCR | 196 | 21 | 10.71 | 8 | [36] |
| 2008 | Germany | Europe | PCR | 293 | 26 | 8.87 | 8 | [58] |
| 1999–2001 | Germany | Europe | PCR | 3113 | 31 | 0.99 | 8 | [59] |
| 2006–2007 | Germany | Europe | PCR | 1000 | 50 | 5.00 | 8 | [60] |
| 2006 | Germany | Europe | PCR | 196 | 21 | 10.71 | 8 | [61] |
| 2011–2012 | Germany | Europe | PCR | 4381 | 45 | 1.03 | 8 | [46] |
| 2011 | Germany | Europe | PCR | 199 | 6 | 3.02 | 8 | [38] |
| 2009–2010 | Germany | Europe | PCR | 6593 | 28 | 0.42 | 8 | [47] |
| 2008–2010 | Germany | Europe | PCR | 1721 | 36 | 2.09 | 8 | [17] |
| 2010–2013 | Germany | Europe | PCR | 339 | 1 | 0.29 | 8 | [24] |
| 2011–2012 | Germany | Europe | PCR | 2000 | 0 | 0 | 8 | [62] |
| * 2010–2018 | Germany, Netherland, Belgium, and Great Britain | Europe | Microfluidic qPCR | 1486 | 16 | 1.08 | 6 | [25] |
| 2006–2008 | Hungary | Europe | PCR | 1800 | 15 | 0.83 | 5 | [63] |
| 2014–2015 | Hungary | Europe | PCR | 413 | 34 | 8.23 | 8 | [21] |
| 2006–2008 | Italy | Europe | PCR | 191 | 1 | 0.52 | 8 | [64] |
| 2006 | Italy | Europe | PCR | 356 | 3 | 0.84 | 6 | [65] |
| 2006–2007 | Italy | Europe | Nested PCR | 1861 | 19 | 1.02 | 7 | [66] |
| 2000–2001 | Italy | Europe | Multiplex PCR | 1931 | 31 | 1.61 | 5 | [67] |
| 2005–2007 | Latvia | Europe | PCR | 1125 | 19 | 1.69 | 8 | [68] |
| 2006 | Latvia and Lithuania | Europe | Nested PCR | 2810 | 40 | 1.42 | 8 | [69] |
| 2006–2008 | Norway | Europe | qPCR, nested PCR | 1908 | 17 | 0.89 | 8 | [70] |
| 2006 | Norway and Lithuania | Europe | qPCR | 364 | 5 | 1.37 | 8 | [71] |
| 2005 | Lithuania | Europe | PCR | 62 | 0 | 0 | 7 | [72] |
| 2003–2007 | Netherlands | Europe | RLB/PCR | 1488 | 16 | 1.08 | 7 | [73] |
| NA | Poland | Europe | Nested PCR | 60 | 35 | 58.33 | 6 | [74] |
| 2008 | Poland | Europe | Nested PCR | 1392 | 22 | 1.58 | 8 | [75] |
| 2009–2012 | Poland | Europe | Nested PCR | 205 | 6 | 2.93 | 5 | [76] |
| 2001 | Poland | Europe | PCR | 1328 | 28 | 2.11 | 7 | [77] |
| 2000–2004 | Poland | Europe | PCR | 1513 | 5 | 0.33 | 8 | [78] |
| 1999 | Poland | Europe | PCR | 2095 | 130 | 6.21 | 8 | [79] |
| 2009–2010 | Poland | Europe | qPCR | 1875 | 47 | 2.51 | 8 | [80] |
| 2009–2010 | Poland | Europe | PCR | 3165 | 50 | 1.58 | 6 | [81] |
| 2008–2009 | Poland | Europe | PCR, nested PCR | 468 | 21 | 4.49 | 8 | [82] |
| 2011–2012 | Poland | Europe | PCR, nested PCR | 1435 | 55 | 3.83 | 8 | [83] |
| 2011 | Poland | Europe | PCR | 634 | 26 | 4.10 | 7 | [84] |
| 2004–2006 | Poland | Europe | Nested PCR | 1620 | 57 | 3.52 | 7 | [85] |
| 2001 | Poland | Europe | Nested PCR | 701 | 16 | 2.28 | 7 | [86] |
| 2006–2008 | Estonia | Europe | RLB, nested PCR | 2603 | 36 | 1.38 | 6 | [87] |
| 2012 | Portugal | Europe | PCR | 263 | 0 | 0.0 | 8 | [88] |
| 2012–2013 | Portugal | Europe | PCR | 175 | 0 | 0.0 | 8 | [89] |
| 2007 | Luxembourg | Europe | PCR | 1394 | 36 | 2.58 | 7 | [90] |
| 2010 | Romania | Europe | PCR | 40 | 0 | 0 | 8 | [91] |
| 2013–2014 | Sweden | Europe | PCR | 519 | 23 | 4.43 | 7 | [92] |
| 2015–2016 | Sweden | Europe | PCR | 277 | 0 | 0 | 8 | [93] |
| 2000 | Russia | Europe | PCR | 738 | 7 | 0.95 | 6 | [94] |
| 2009 | Russia | Europe | PCR | 481 | 5 | 1.04 | 6 | [95] |
| 2003–2004 | Russia | Europe | Nested PCR | 209 | 3 | 1.44 | 6 | [22] |
| 2008–2009 | Russia | Europe | Nested PCR | 922 | 24 | 2.60 | 6 | [96] |
| 2010–2015 | Russia | Europe | Nested PCR | 911 | 4 | 0.44 | 6 | [31] |
| 2002 | Slovakia | Europe | PCR | 100 | 1 | 1.0 | 8 | [97] |
| 2011 | Slovakia | Europe | PCR | 5148 | 78 | 1.63 | 8 | [98] |
| 2011–2012 | Slovakia | Europe | PCR | 886 | 12 | 1.35 | 7 | [99] |
| 1997 | Slovenia | Europe | PCR | 135 | 13 | 9.63 | 7 | [100] |
| 2003 | Czech Republic | Europe | PCR | 350 | 5 | 1.43 | 8 | [101] |
| 2011–2014 | Czech Republic | Europe | PCR | 2473 | 32 | 1.29 | 8 | [102] |
| 1997 | Belgium | Europe | PCR | 230 | 0 | 0 | 6 | [103] |
| 2011–2013 | Netherlands and Belgium | Europe | RLB/PCR | 855 | 17 | 1.99 | 7 | [41] |
| 2003–2005 | Spain | Europe | RLB/PCR | 562 | 17 | 3.03 | 8 | [44] |
| 2002–2003 | Switzerland | Europe | PCR | 865 | 4 | 0.46 | 7 | [104] |
| 2006 | Switzerland | Europe | RLB/PCR | 2568 | 44 | 1.71 | 8 | [105] |
| 2009–2010 | Switzerland | Europe | RLB/PCR | 1476 | 28 | 1.89 | 7 | [39] |
| 2015–2016 | Switzerland | Europe | qPCR | 1079 | 6 | 0.56 | 8 | [106] |
| 2012 | Switzerland | Europe | PCR | 261 | 16 | 6.13 | 8 | [23] |
| 2013–2014 | Ukraine | Europe | PCR | 767 | 13 | 1.69 | 7 | [107] |
| 2011–2013 | Turkey | Europe-Asia | NGS | 205 | 1 | 0.49 | 7 | [33] |
| 2014–2018 | Turkey | Europe-Asia | PCR | 1019 | 27 | 2.65 | 8 | [108] |
| 2013–2014 | China | Asia | RLB/PCR | 450 | 37 | 8.22 | 8 | [43] |
| 2013–2014 | China | Asia | Nested PCR | 558 | 2 | 0.36 | 8 | [8] |
| 2013–2014 | China | Asia | Nested PCR | 797 | 51 | 6.39 | 7 | [28] |
| 2013–2014 | Israel | Asia | PCR | 1196 | 3 | 0.25 | 6 | [109] |
| 2013–2015 | Japan | Asia | Nested PCR | 624 | 5 | 0.80 | 8 | [110] |
| 2000–2003 | Japan | Asia | Nested PCR | 1656 | 40 | 2.42 | 8 | [37] |
| 2008 | Japan | Asia | PCR | 1459 | 18 | 1.23 | 8 | [29] |
| 2000–2003 | Japan | Asia | PCR | 294 | 17 | 5.78 | 8 | [30] |
| NA | Mongolia | Asia | Nested PCR | 108 | 7 | 6.48 | 6 | [45] |
| 2009 | Mongolia | Asia | PCR | 400 | 9 | 2.25 | 8 | [111] |
| 2012–2013 | Mongolia | Asia | Nested PCR | 219 | 19 | 8.68 | 7 | [32] |
| 2015 | Thailand | Asia | PCR | 12,184 | 1 | 0.01 | 8 | [112] |
| 2009 | Nigeria | Africa | PCR | 700 | 0 | 0 | 8 | [34] |
| 2001 | United States of America | North America | PCR | 107 | 9 | 8.41 | 6 | [113] |
| 2013–2014 | United States of America | North America | PCR | 423 | 3 | 0.71 | 6 | [114] |
| 1985 | United States of America | North America | PCR | 395 | 48 | 12.15 | 8 | [115] |
| 1996 | United States of America | North America | PCR | 100 | 5 | 5.0 | 6 | [116] |
| 2003–2006 | United States of America | North America | PCR | 394 | 41 | 10.41 | 7 | [117] |
| 2003 | United States of America | North America | PCR | 68 | 7 | 10.29 | 6 | [42] |
| 2015–2017 | United States of America | North America | HRM | 1721 | 62 | 3.60 | 8 | [118] |
| 2010 | United States of America | North America | PCR | 191 | 0 | 0 | 8 | [119] |
| 2012–2014 | United States of America | North America | qPCR | 1855 | 54 | 2.91 | 8 | [120] |
| 2003–2004 | United States of America | North America | Multiplex PCR | 11,184 | 283 | 2.53 | 8 | [27] |
| 2011 | United States of America | North America | PCR | 1245 | 35 | 2.81 | 7 | [35] |
| 2011 | United States of America | North America | qPCR | 4368 | 255 | 5.84 | 8 | [121] |
| 2016–2017 | Canada | North America | PCR | 249 | 4 | 1.61 | 8 | [26] |
| Subgroup | Number of Studies | Pooled Prevalence Estimates | Measure of Heterogeneity | ||||
|---|---|---|---|---|---|---|---|
| Sample Size | No of Positives | Weighted MIR95% CI (%) | Q Value | I2 | Q−p | ||
| All studies | 104 | 137,364 | 3069 | 2.10 (1.60–2.70) | 4438.97 | 97.65 | p < 0.0001 |
| Tick species | |||||||
| Ixodes ricinus | 57 | 74,802 | 1756 | 2.40 (1.50–3.60) | 3737.86 | 98.50 | p < 0.0001 |
| I. persulcatus | 14 | 5823 | 102 | 1.50 (0.70–3.20) | 154.44 | 91.58 | p < 0.0001 |
| I. ovatus | 3 | 1420 | 39 | 0.60 (0.00–9.20) | 17.23 | 88.39 | p < 0.0001 |
| I. scapularis | 14 | 22,694 | 786 | 4.10 (2.70–6.20) | 296.36 | 95.95 | p < 0.0001 |
| I. pavlovskyi | 1 | 577 | 2 | 0.30 (0.01–1.40) | − | − | − |
| Dermacentor reticulatus | 20 | 11,802 | 197 | 2.10 (1.30–3.50) | 174.89 | 89.14 | p < 0.0001 |
| D. marginatus | 2 | 390 | 1 | 0.80 (0.10–9.4) | 2.26 | 55.65 | p < 0.0001 |
| D. nuttalli | 3 | 389 | 7 | 1.30 (0.10–12.10) | 7.60 | 73.76 | p = 0.022 |
| D. silvarum | 2 | 223 | 4 | 1.80 (0.20–18.50) | 3.06 | 67.23 | p = 0.080 |
| R. bursa | 4 | 120 | 2 | 2.90 (0.90–8.50) | 0.99 | 0.00 | p = 0.802 |
| R. sanguineus s.l. | 5 | 1668 | 3 | 0.60 (0.10–2.60) | 8.77 | 54.39 | p < 0.001 |
| R. (Boophilus) microplus | 3 | 1498 | 2 | 0.30 (0.10–1.90) | 1.63 | 0.00 | p = 0.443 |
| R. turanicus | 1 | 9 | 1 | 11.1 (1.50–50.00) | 0.00 | 0.00 | p = 1.000 |
| Hemaphysalis longicornis | 5 | 626 | 28 | 4.30 (1.60–10.90) | 13.17 | 69.62 | p = 0.010 |
| H. concinna | 4 | 130 | 6 | 6.10 (3.00–11.90) | 0.760 | 0.00 | p = 0.825 |
| H. qinghaiensis | 2 | 430 | 73 | 17.20 (10.90–26.0) | 4.32 | 76.86 | p = 0.038 |
| H. punctata | 1 | 111 | 4 | 3.60 (1.40–9.20) | 0.00 | 0.00 | p = 1.000 |
| H. parva | 1 | 793 | 13 | 1.60 (1.00–2.80) | 0.00 | 0.00 | p = 1.000 |
| H. inermis | 1 | 87 | 1 | 1.10 (0.20–7.70) | 0.00 | 0.00 | p = 1.000 |
| H. flava | 2 | 282 | 3 | 1.30 (0.50–3.80) | 0.49 | - | p = 0.484 |
| H. formosensis | 1 | 159 | 2 | 1.30 (0.30–4.90) | 0.00 | 0.00 | p = 1.000 |
| H. lagrangei | 1 | 11,309 | 1 | 0.00 (0.00–0.01) | 0.00 | 0.00 | p = 1.000 |
| Hyalomma marginatum | 1 | 105 | 13 | 12.38 (7.30–20.20) | 0.00 | 0.00 | p = 1.000 |
| Life stages | |||||||
| Adult | 79 | 55,411 | 1484 | 2.60 (2.00–3.40) | 1693.34 | 95.34 | p < 0.0001 |
| Nymphs | 53 | 44,746 | 1066 | 1.70 (1.10–2.50) | 1578.82 | 96.77 | p < 0.0001 |
| Larvae | 13 | 20,866 | 174 | 0.60 (0.10–3.60) | 699.77 | 98.29 | p < 0.0001 |
| Sex | |||||||
| Male | 26 | 7534 | 199 | 3.60 (3.10–4.20) | 145.53 | 82.82 | p < 0.0001 |
| Female | 26 | 8395 | 275 | 4.90 (4.40–5.60) | 256.98 | 90.27 | p < 0.0001 |
| Diagnostic technique | |||||||
| Conventional PCR | 66 | 76,021 | 1663 | 1.90 (1.30–2.90) | 3339.99 | 98.05 | p < 0.0001 |
| qPCR | 12 | 23,314 | 522 | 1.70 (1.00–3.00) | 332.86 | 96.69 | p < 0.0001 |
| Nested PCR | 16 | 14,653 | 376 | 2.80 (1.70–4.70) | 339.97 | 95.59 | p < 0.0001 |
| RLB | 7 | 10,002 | 195 | 2.20 (1.30–3.80) | 85.88 | 92.99 | p < 0.0001 |
| Multiplex PCR | 2 | 13,115 | 246 | 1.90 (1.70–2.10) | 0.89 | 0.00 | p = 0.344 |
| NGS | 1 | 205 | 2 | 1.00 (0.20–3.80) | 0.00 | 0.00 | p = 1.000 |
| Subgroup | Number of Studies | Pooled Prevalence Estimates | Measure of Heterogeneity | ||||
|---|---|---|---|---|---|---|---|
| Sample Size | No of Positives | Weighted MIR95% CI (%) | Q Value | I2 | Q−p | ||
| All studies | 104 | 137,364 | 3069 | 2.10 (1.60–2.70) | 4438.41 | 97.68 | p < 0.0001 |
| Babesiaspecies | |||||||
| Babesia microti | 46 | 68,537 | 1425 | 1.90 (1.40–2.50) | 1071.94 | 95.80 | p < 0.0001 |
| B. venatorum | 31 | 50,611 | 370 | 0.90 (0.70–1.10) | 163.47 | 81.65 | p < 0.0001 |
| B. divergens | 20 | 33,517 | 141 | 0.40 (0.20–0.70) | 161.75 | 88.47 | p < 0.0001 |
| B. spp. | 19 | 38,125 | 183 | 0.50 (0.20–1.10) | 363.19 | 95.59 | p < 0.0001 |
| B. capreoli | 6 | 15,927 | 13 | 0.10 (0.10–0.20) | 7.31 | 31.61 | p = 0.199 |
| B. canis | 15 | 14,938 | 132 | 1.10 (0.50–2.40) | 235.83 | 94.06 | p < 0.0001 |
| B. odocoilei | 6 | 8002 | 52 | 0.90 (0.20–4.50) | 102.06 | 95.10 | p < 0.0001 |
| B. caballi | 3 | 1525 | 17 | 1.40 (0.30–6.90) | 20.02 | 90.01 | p < 0.0001 |
| B. bovis | 2 | 1012 | 3 | 0.30 (0.10–0.90) | 0.15 | 0.00 | p = 0.700 |
| B. bigemina | 3 | 1570 | 7 | 0.50 (0.20–1.40) | 2.75 | 27.16 | p = 0.253 |
| B. ovata | 2 | 1909 | 18 | 0.60 (0.10–5.00) | 2.85 | 64.87 | p = 0.092 |
| B. spp. Xinjiang | 2 | 1247 | 84 | 6.70 (5.50–8.30) | 0.39 | 0.00 | p = 0.528 |
| B. gibsoni | 1 | 6593 | 1 | 0.00 (0.00–0.10) | 0.00 | 0.00 | p = 1.000 |
| B. ovis | 1 | 205 | 1 | 0.50 (0.10–3.40) | 0.00 | 0.00 | p = 1.000 |
| B. occultans | 1 | 1019 | 12 | 1.20 (0.70–2.10) | − | − | − |
| B. rossi | 1 | 1019 | 4 | 0.40 (0.10–1.00) | 0.00 | 0.00 | p = 1.000 |
| B. vogeli | 1 | 1196 | 3 | 1.50 (0.00–32.40) | |||
| B. crassa | 1 | 1019 | 8 | 0.80 (0.40–1.60) | 0.00 | 0.00 | p = 1.000 |
| B. motasi like | 1 | 450 | 3 | 0.70 (0.20–2.00) | 0.00 | 0.00 | p = 1.000 |
| Region | |||||||
| Europe | 78 | 94,376 | 2056 | 1.90 (1.30–2.70) | 3964.12 | 98.06 | p < 0.0001 |
| Asia | 12 | 19,945 | 209 | 2.00 (1.10–3.50) | 174.67.69 | 93.70 | p < 0.0001 |
| North America | 13 | 22,299 | 806 | 4.30 (3.00–6.20) | 237.73 | 94.95 | p < 0.0001 |
| Sampling period | |||||||
| 1992–1997 (period 1) | 3 | 465 | 18 | 4.30 (1.30–13.90) | 8.28 | 75.85 | p = 0.016 |
| 1998–2002 (period 2) | 9 | 10,205 | 269 | 2.90 (1.40–5.70) | 205.79 | 96.11 | p < 0.0001 |
| 2003–2008 (period 3) | 29 | 39,266 | 1326 | 2.60 (1.40–4.80) | 2628.50 | 98.94 | p < 0.0001 |
| 2009–2014 (period 4) | 38 | 52,571 | 950 | 1.60 (1.20–2.20) | 627.33 | 94.10 | p < 0.0001 |
| 2015–2020 (period 5) | 10 | 20,722 | 103 | 0.90 (0.40–2.10) | 112.84 | 92.91 | p < 0.0001 |
| Subgroup | Number of Studies | Pooled Prevalence Estimates | Measure of Heterogeneity | ||||
|---|---|---|---|---|---|---|---|
| Sample Size | No of Positives | Weighted MIR95% CI (%) | Q Value | I2 | Q−p | ||
| Austria | 1 | 864 | 441 | 51.00 (47.70–54.40) | − | − | − |
| Belarus | 1 | 453 | 5 | 1.10 (0.50–2.60) | − | − | − |
| Denmark | 1 | 1013 | 5 | 0.50 (0.20–1.20) | − | − | − |
| Finland | 2 | 7585 | 47 | 0.70 (0.40–1.40) | 2.56 | 60.97 | p = 0.109 |
| France | 6 | 4397 | 87 | 3.30 (0.90–10.80) | 148.22 | 96.62 | p < 0.0001 |
| Germany | 12 | 20,257 | 273 | 2.20 (1.10–4.40) | 326.82 | 96.63 | p < 0.0001 |
| Hungary | 2 | 2213 | 49 | 2.70 (0.30–22.0) | 56.48 | 98.23 | p < 0.0001 |
| Italy | 4 | 4339 | 54 | 1.20 (0.90–1.70) | 3.93 | 23.64 | p = 0.269 |
| Latvia | 2 | 1306 | 24 | 1.90 (1.30–2.80) | 0.98 | 0.00 | p = 0.323 |
| Norway | 2 | 2132 | 19 | 0.90 (0.60–1.40) | 0.00 | 0.00 | p = 0.998 |
| Lithuania | 3 | 2831 | 64 | 2.30 (1.80–2.90) | 0.59 | 0.00 | p = 0.042 |
| Netherland | 3 | 2893 | 32 | 1.20 (0.40–3.50) | 13.34 | 85.01 | p = 0.000 |
| Poland | 13 | 16,491 | 498 | 3.40 (2.10–5.50) | 330.43 | 96.37 | p < 0.0001 |
| Estonia | 1 | 2603 | 36 | 1.40 (1.00–1.90) | − | − | − |
| Portugal | 2 | 438 | 0 | 0.02 (0.00–1.60) | 0.041 | 0.00 | p = 0.839 |
| Luxembourg | 1 | 1394 | 36 | 2.60 (1.90–3.60) | 0.00 | 0.00 | − |
| Romania | 1 | 40 | 0 | 1.20 (0.10–16.70) | − | − | − |
| Sweden | 2 | 796 | 23 | 1.20 (0.10–22.0) | 5.14 | 80.53 | p = 0.023 |
| Russia | 5 | 3261 | 43 | 1.20 (0.60–2.30) | 15.48 | 74.15 | p = 0.004 |
| Slovakia | 3 | 6130 | 97 | 1.60 (1.30–1.90) | 0.57 | 0.00 | p = 0.751 |
| Slovenia | 1 | 135 | 13 | 7.40 (4.00–13.20) | − | − | − |
| Czech Republic | 2 | 2823 | 37 | 1.30 (1.00–1.80) | 0.04 | 0.00 | p = 0.836 |
| Belgium | 3 | 1053 | 1 | 0.20 (0.10–0.90) | 0.54 | 0.00 | p = 0.761 |
| Britain | 1 | 113 | 16 | 14.20 (8.90–21.90) | − | − | − |
| Turkey | 2 | 1224 | 28 | 2.00 (0.80–4.80) | 1.90 | 47.41 | p = 0.168 |
| Spain | 1 | 562 | 17 | 3.00 (1.90–4.80) | − | − | − |
| Switzerland | 5 | 6259 | 98 | 1.50 (0.80–3.00) | 40.02 | 90.00 | p < 0.0001 |
| Ukraine | 1 | 767 | 13 | 1.90 (1.10–3.20) | − | − | − |
| China | 3 | 1805 | 90 | 4.10 (1.90–9.0) | 19.63 | 89.81 | p < 0.0001 |
| Israel | 1 | 1196 | 3 | 0.30 (0.10–0.80) | − | − | − |
| Japan | 4 | 4033 | 80 | 2.00 (1.00–4.20) | 27.61 | 89.14 | p < 0.0001 |
| Mongolia | 3 | 727 | 35 | 5.10 (2.20–11.50) | 11.87 | 83.15 | p = 0.003 |
| Thailand | 1 | 12,184 | 1 | 0.00 (0.00–0.10) | − | − | − |
| Nigeria | 1 | 700 | 0 | 0.00 (0.00–0.00) | − | − | − |
| United States | 12 | 22,300 | 806 | 4.30 (3.00–6.20) | 237.33 | 94.95 | p < 0.0001 |
| Canada | 1 | 248 | 4 | 1.60 (0.60–4.20) | − | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Onyiche, T.E.; Răileanu, C.; Fischer, S.; Silaghi, C. Global Distribution of Babesia Species in Questing Ticks: A Systematic Review and Meta-Analysis Based on Published Literature. Pathogens 2021, 10, 230. https://doi.org/10.3390/pathogens10020230
Onyiche TE, Răileanu C, Fischer S, Silaghi C. Global Distribution of Babesia Species in Questing Ticks: A Systematic Review and Meta-Analysis Based on Published Literature. Pathogens. 2021; 10(2):230. https://doi.org/10.3390/pathogens10020230
Chicago/Turabian StyleOnyiche, ThankGod E., Cristian Răileanu, Susanne Fischer, and Cornelia Silaghi. 2021. "Global Distribution of Babesia Species in Questing Ticks: A Systematic Review and Meta-Analysis Based on Published Literature" Pathogens 10, no. 2: 230. https://doi.org/10.3390/pathogens10020230
APA StyleOnyiche, T. E., Răileanu, C., Fischer, S., & Silaghi, C. (2021). Global Distribution of Babesia Species in Questing Ticks: A Systematic Review and Meta-Analysis Based on Published Literature. Pathogens, 10(2), 230. https://doi.org/10.3390/pathogens10020230