
Squalene-Based Influenza Vaccine Adjuvants and Their Impact on the Hemagglutinin-Specific B Cell Response


Abstract
:1. Introduction
2. Squalene-Based Emulsion (SE) Adjuvant Overview
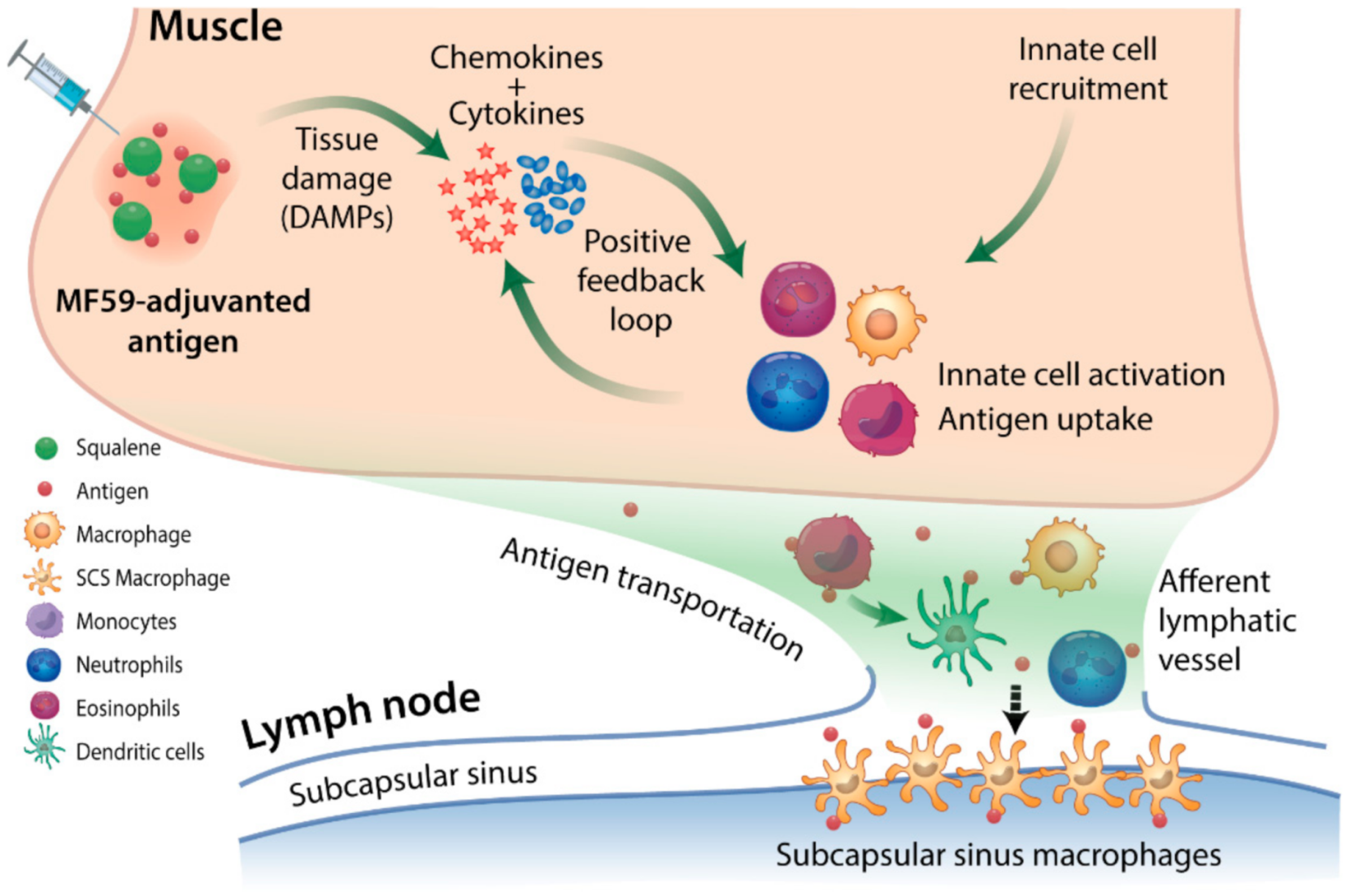
3. SE Adjuvants Induce an ‘Immunocompetent Environment’ Leading to Enhanced Antigen Uptake and Delivery

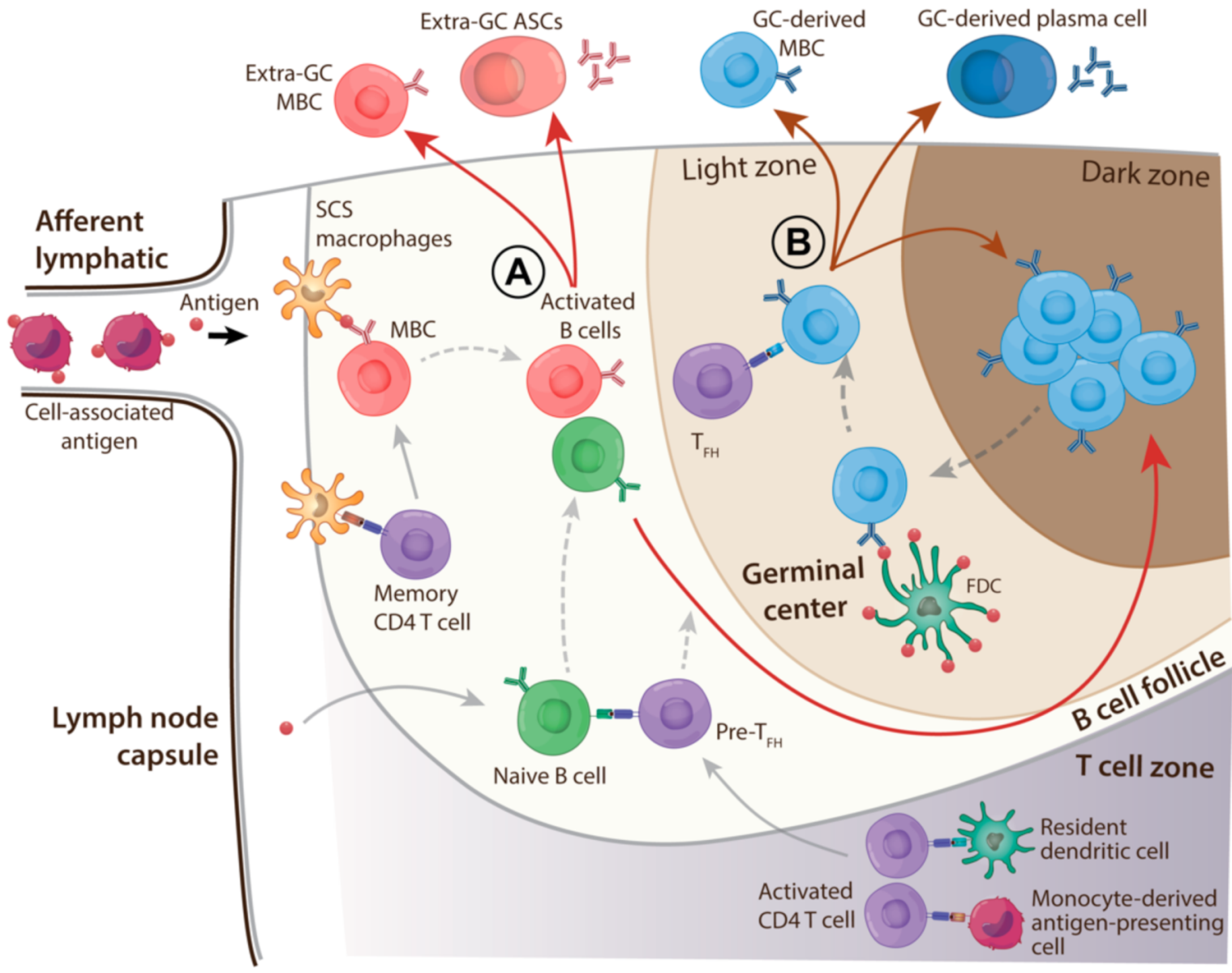
4. SE Adjuvants Strongly Promote Adaptive B Cell Responses
4.1. B Cell Competition
4.2. Mechanisms of Enhancement of B Cell Responses by SE Adjuvants
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gasparini, R.; Amicizia, D.; Lai, P.L.; Panatto, D. Clinical and Socioeconomic Impact of Seasonal and Pandemic Influenza in Adults and the Elderly. Hum. Vaccines Immunother. 2012, 8, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iuliano, A.D.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Gran, J.M.; Schanzer, D.; Cowling, B.J.; et al. Estimates of Global Seasonal Influenza-Associated Respiratory Mortality: A Modelling Study. Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef]
- Houser, K.; Subbarao, K. Influenza Vaccines: Challenges and Solutions. Cell Host Microbe 2015, 17, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Buisman, A.M.; de Rond, C.G.H.; Öztürk, K.; ten Hulscher, H.I.; van Binnendijk, R.S. Long-Term Presence of Memory B-Cells Specific for Different Vaccine Components. Vaccine 2009, 28, 179–186. [Google Scholar] [CrossRef]
- Reed, S.G.; Orr, M.T.; Fox, C.B. Key Roles of Adjuvants in Modern Vaccines. Nat. Med. 2013, 19, 1597–1608. [Google Scholar] [CrossRef]
- Dawood, F.S.; Chung, J.R.; Kim, S.S.; Zimmerman, R.K.; Nowalk, M.P.; Jackson, M.L.; Jackson, L.A.; Monto, A.S.; Martin, E.T.; Belongia, E.A.; et al. Interim Estimates of 2019–20 Seasonal Influenza Vaccine Effectiveness—United States, February 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Belongia, E.A.; Kieke, B.A.; Donahue, J.G.; Greenlee, R.T.; Balish, A.; Foust, A.; Lindstrom, S.; Shay, D.K. Marshfield Influenza Study Group Effectiveness of Inactivated Influenza Vaccines Varied Substantially with Antigenic Match from the 2004–2005 Season to the 2006–2007 Season. J. Infect. Dis. 2009, 199, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, R.K.; Nowalk, M.P.; Chung, J.; Jackson, M.L.; Jackson, L.A.; Petrie, J.G.; Monto, A.S.; McLean, H.Q.; Belongia, E.A.; Gaglani, M.; et al. 2014–2015 Influenza Vaccine Effectiveness in the United States by Vaccine Type. Clin. Infect. Dis. 2016, 63, 1564–1573. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhang, J.; Han, T.; Liu, C.; Li, X.; Yan, L.; Yang, B.; Yang, X. Effectiveness, Immunogenicity, and Safety of Influenza Vaccines with MF59 Adjuvant in Healthy People of Different Age Groups: A Systematic Review and Meta-Analysis. Medicine 2020, 99, e19095. [Google Scholar] [CrossRef] [PubMed]
- Tregoning, J.S.; Russell, R.F.; Kinnear, E. Adjuvanted Influenza Vaccines. Hum. Vaccines Immunother. 2018, 14, 550–564. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, A.; Omer, S.B. Why and How Vaccines Work. Cell 2020, 183, 290–295. [Google Scholar] [CrossRef]
- Leroux-Roels, I.; Borkowski, A.; Vanwolleghem, T.; Dramé, M.; Clement, F.; Hons, E.; Devaster, J.-M.; Leroux-Roels, G. Antigen Sparing and Cross-Reactive Immunity with an Adjuvanted RH5N1 Prototype Pandemic Influenza Vaccine: A Randomised Controlled Trial. Lancet Lond. Engl. 2007, 370, 580–589. [Google Scholar] [CrossRef]
- Moris, P.; van der Most, R.; Leroux-Roels, I.; Clement, F.; Dramé, M.; Hanon, E.; Leroux-Roels, G.G.; Van Mechelen, M. H5N1 Influenza Vaccine Formulated with AS03 A Induces Strong Cross-Reactive and Polyfunctional CD4 T-Cell Responses. J. Clin. Immunol. 2011, 31, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.S.; Bizjajeva, S.; Lindert, K.; Heijnen, E.; Oberye, J. Cumulative Clinical Experience with MF59-Adjuvanted Trivalent Seasonal Influenza Vaccine in Young Children. Int. J. Infect. Dis. 2019, 85, S26–S38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultze, V.; D’Agosto, V.; Wack, A.; Novicki, D.; Zorn, J.; Hennig, R. Safety of MF59TM Adjuvant. Vaccine 2008, 26, 3209–3222. [Google Scholar] [CrossRef] [PubMed]
- Langley, J.M.; Frenette, L.; Ferguson, L.; Riff, D.; Sheldon, E.; Risi, G.; Johnson, C.; Li, P.; Kenney, R.; Innis, B.; et al. Safety and Cross-Reactive Immunogenicity of Candidate AS03-Adjuvanted Prepandemic H5N1 Influenza Vaccines: A Randomized Controlled Phase 1/2 Trial in Adults. J. Infect. Dis. 2010, 201, 1644–1653. [Google Scholar] [CrossRef] [Green Version]
- Ott, G.; Barchfeld, G.L.; Chernoff, D.; Radhakrishnan, R.; van Hoogevest, P.; Van Nest, G. MF59 Design and Evaluation of a Safe and Potent Adjuvant for Human Vaccines. In Vaccine Design; Powell, M.F., Newman, M.J., Eds.; Pharmaceutical Biotechnology Springer US: Boston, MA, USA, 1995; Volume 6, pp. 277–296. ISBN 978-1-4613-5737-7. [Google Scholar]
- Levie, K.; Leroux-Roels, I.; Hoppenbrouwers, K.; Kervyn, A.; Vandermeulen, C.; Forgus, S.; Leroux-Roels, G.; Pichon, S.; Kusters, I. An Adjuvanted, Low-Dose, Pandemic Influenza A (H5N1) Vaccine Candidate Is Safe, Immunogenic, and Induces Cross-Reactive Immune Responses in Healthy Adults. J. Infect. Dis. 2008, 198, 642–649. [Google Scholar] [CrossRef]
- Hauser, M.I.; Muscatello, D.J.; Soh, A.C.Y.; Dwyer, D.E.; Turner, R.M. An Indirect Comparison Meta-Analysis of AS03 and MF59 Adjuvants in Pandemic Influenza A(H1N1)Pdm09 Vaccines. Vaccine 2019, 37, 4246–4255. [Google Scholar] [CrossRef]
- Vanderven, H.A.; Barr, I.; Reynaldi, A.; Wheatley, A.K.; Wines, B.D.; Davenport, M.P.; Hogarth, P.M.; Kent, S.J. Fc Functional Antibody Responses to Adjuvanted versus Unadjuvanted Seasonal Influenza Vaccination in Community-Dwelling Older Adults. Vaccine 2020, 38, 2368–2377. [Google Scholar] [CrossRef]
- Rudicell, R.S.; Garinot, M.; Kanekiyo, M.; Kamp, H.D.; Swanson, K.; Chou, T.-H.; Dai, S.; Bedel, O.; Simard, D.; Gillespie, R.A.; et al. Comparison of Adjuvants to Optimize Influenza Neutralizing Antibody Responses. Vaccine 2019, 37, 6208–6220. [Google Scholar] [CrossRef]
- O’Hagan, D.T.; Friedland, L.R.; Hanon, E.; Didierlaurent, A.M. Towards an Evidence Based Approach for the Development of Adjuvanted Vaccines. Curr. Opin. Immunol. 2017, 47, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Vogel, F.R.; Caillet, C.; Kusters, I.C.; Haensler, J. Emulsion-Based Adjuvants for Influenza Vaccines. Expert Rev. Vaccines 2009, 8, 483–492. [Google Scholar] [CrossRef]
- O’Hagan, D.T.; Ott, G.S.; De Gregorio, E.; Seubert, A. The Mechanism of Action of MF59—An Innately Attractive Adjuvant Formulation. Vaccine 2012, 30, 4341–4348. [Google Scholar] [CrossRef]
- Del Giudice, G.; Rappuoli, R.; Didierlaurent, A.M. Correlates of Adjuvanticity: A Review on Adjuvants in Licensed Vaccines. Semin. Immunol. 2018, 39, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.-J.; Kang, S.-M. Immunology and Efficacy of MF59-Adjuvanted Vaccines. Hum. Vaccines Immunother. 2018, 14, 3041–3045. [Google Scholar] [CrossRef]
- Awate, S.; Babiuk, L.A.; Mutwiri, G. Mechanisms of Action of Adjuvants. Front. Immunol. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ott, G.; Barchfeld, G.L.; Van Nest, G. Enhancement of Humoral Response against Human Influenza Vaccine with the Simple Submicron Oil/Water Emulsion Adjuvant MF59. Vaccine 1995, 13, 1557–1562. [Google Scholar] [CrossRef]
- Klucker, M.; Dalençon, F.; Probeck, P.; Haensler, J. AF03, An Alternative Squalene Emulsion-Based Vaccine Adjuvant Prepared by a Phase Inversion Temperature Method. J. Pharm. Sci. 2012, 101, 4490–4500. [Google Scholar] [CrossRef]
- Baras, B.; Bouveret, N.; Devaster, J.-M.; Fries, L.; Gillard, P.; Sänger, R.; Hanon, E. A Vaccine Manufacturer’s Approach to Address Medical Needs Related to Seasonal and Pandemic Influenza Viruses: GSK Vaccine Portfolio against Influenza. Influenza Other Respir. Viruses 2008, 2, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Noe, S.M.; Green, M.A.; HogenEsch, H.; Hem, S.L. Mechanism of Immunopotentiation by Aluminum-Containing Adjuvants Elucidated by the Relationship between Antigen Retention at the Inoculation Site and the Immune Response. Vaccine 2010, 28, 3588–3594. [Google Scholar] [CrossRef]
- Hutchison, S.; Benson, R.A.; Gibson, V.B.; Pollock, A.H.; Garside, P.; Brewer, J.M. Antigen Depot Is Not Required for Alum Adjuvanticity. FASEB J. 2012, 26, 1272–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, G.K.; Wørzner, K.; Andersen, P.; Christensen, D. Vaccine Adjuvants Differentially Affect Kinetics of Antibody and Germinal Center Responses. Front. Immunol. 2020, 11, 579761. [Google Scholar] [CrossRef] [PubMed]
- Nakaya, H.I.; Clutterbuck, E.; Kazmin, D.; Wang, L.; Cortese, M.; Bosinger, S.E.; Patel, N.B.; Zak, D.E.; Aderem, A.; Dong, T.; et al. Systems Biology of Immunity to MF59-Adjuvanted versus Nonadjuvanted Trivalent Seasonal Influenza Vaccines in Early Childhood. Proc. Natl. Acad. Sci. USA 2016, 113, 1853–1858. [Google Scholar] [CrossRef] [Green Version]
- Vono, M.; Taccone, M.; Caccin, P.; Gallotta, M.; Donvito, G.; Falzoni, S.; Palmieri, E.; Pallaoro, M.; Rappuoli, R.; Di Virgilio, F.; et al. The Adjuvant MF59 Induces ATP Release from Muscle That Potentiates Response to Vaccination. Proc. Natl. Acad. Sci. USA 2013, 110, 21095–21100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabro, S.; Tritto, E.; Pezzotti, A.; Taccone, M.; Muzzi, A.; Bertholet, S.; De Gregorio, E.; O’Hagan, D.T.; Baudner, B.; Seubert, A. The Adjuvant Effect of MF59 Is Due to the Oil-in-Water Emulsion Formulation, None of the Individual Components Induce a Comparable Adjuvant Effect. Vaccine 2013, 31, 3363–3369. [Google Scholar] [CrossRef]
- Morel, S.; Didierlaurent, A.; Bourguignon, P.; Delhaye, S.; Baras, B.; Jacob, V.; Planty, C.; Elouahabi, A.; Harvengt, P.; Carlsen, H. Adjuvant System AS03 Containing α-Tocopherol Modulates Innate Immune Response and Leads to Improved Adaptive Immunity. Vaccine 2011, 29, 2461–2473. [Google Scholar] [CrossRef]
- Liang, F.; Lindgren, G.; Sandgren, K.J.; Thompson, E.A.; Francica, J.R.; Seubert, A.; De Gregorio, E.; Barnett, S.; O’Hagan, D.T.; Sullivan, N.J.; et al. Vaccine Priming Is Restricted to Draining Lymph Nodes and Controlled by Adjuvant-Mediated Antigen Uptake. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef]
- Tegenge, M.A.; Von Tungeln, L.S.; Anderson, S.A.; Mitkus, R.J.; Vanlandingham, M.M.; Forshee, R.A.; Beland, F.A. Comparative Pharmacokinetic and Biodistribution Study of Two Distinct Squalene-Containing Oil-in-Water Emulsion Adjuvants in H5N1 Influenza Vaccines. Regul. Toxicol. Pharmacol. 2019, 108, 104436. [Google Scholar] [CrossRef]
- Calabro, S.; Tortoli, M.; Baudner, B.C.; Pacitto, A.; Cortese, M.; O’Hagan, D.T.; De Gregorio, E.; Seubert, A.; Wack, A. Vaccine Adjuvants Alum and MF59 Induce Rapid Recruitment of Neutrophils and Monocytes That Participate in Antigen Transport to Draining Lymph Nodes. Vaccine 2011, 29, 1812–1823. [Google Scholar] [CrossRef]
- Moyer, T.J.; Kato, Y.; Abraham, W.; Chang, J.Y.H.; Kulp, D.W.; Watson, N.; Turner, H.L.; Menis, S.; Abbott, R.K.; Bhiman, J.N.; et al. Engineered Immunogen Binding to Alum Adjuvant Enhances Humoral Immunity. Nat. Med. 2020, 26, 430–440. [Google Scholar] [CrossRef]
- Mosca, F.; Tritto, E.; Muzzi, A.; Monaci, E.; Bagnoli, F.; Iavarone, C.; O’Hagan, D.; Rappuoli, R.; De Gregorio, E. Molecular and Cellular Signatures of Human Vaccine Adjuvants. Proc. Natl. Acad. Sci. USA 2008, 105, 10501–10506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tizard, I.R. Adjuvants and adjuvanticity. In Vaccines for Veterinarians; Elsevier: Amsterdam, The Netherlands, 2021; pp. 75.e1–86.e1. ISBN 978-0-323-68299-2. [Google Scholar]
- Kim, E.H.; Woodruff, M.C.; Grigoryan, L.; Maier, B.; Lee, S.H.; Mandal, P.; Cortese, M.; Natrajan, M.S.; Ravindran, R.; Ma, H.; et al. Squalene Emulsion-Based Vaccine Adjuvants Stimulate CD8 T Cell, but Not Antibody Responses, through a RIPK3-Dependent Pathway. eLife 2020, 9, e52687. [Google Scholar] [CrossRef] [PubMed]
- Seubert, A.; Monaci, E.; Pizza, M.; O’Hagan, D.T.; Wack, A. The Adjuvants Aluminum Hydroxide and MF59 Induce Monocyte and Granulocyte Chemoattractants and Enhance Monocyte Differentiation toward Dendritic Cells. J. Immunol. 2008, 180, 5402–5412. [Google Scholar] [CrossRef] [PubMed]
- Ellebedy, A.H.; Lupfer, C.; Ghoneim, H.E.; DeBeauchamp, J.; Kanneganti, T.-D.; Webby, R.J. Inflammasome-Independent Role of the Apoptosis-Associated Speck-like Protein Containing CARD (ASC) in the Adjuvant Effect of MF59. Proc. Natl. Acad. Sci. USA 2011, 108, 2927–2932. [Google Scholar] [CrossRef] [Green Version]
- Dupuis, M.; Denis-Mize, K.; LaBarbara, A.; Peters, W.; Charo, I.F.; McDonald, D.M.; Ott, G. Immunization with the Adjuvant MF59 Induces Macrophage Trafficking and Apoptosis. Eur. J. Immunol. 2001, 31, 2910–2918. [Google Scholar] [CrossRef]
- Fanger, N.A.; Liu, C.; Guyre, P.M.; Wardwell, K.; O’Neil, J.; Guo, T.L.; Christian, T.P.; Mudzinski, S.P.; Gosselin, E.J. Activation of Human T Cells by Major Histocompatability Complex Class II Expressing Neutrophils: Proliferation in the Presence of Superantigen, but Not Tetanus Toxoid. Blood 1997, 89, 4128–4135. [Google Scholar] [CrossRef] [PubMed]
- Radsak, M.; Iking-Konert, C.; Stegmaier, S.; Andrassy, K.; Hansch, G.M. Polymorphonuclear Neutrophils as Accessory Cells for T-Cell Activation: Major Histocompatibility Complex Class II Restricted Antigen-Dependent Induction of T-Cell Proliferation. Immunology 2000, 101, 521–530. [Google Scholar] [CrossRef]
- Reinisch, W.; Tillinger, W.; Lichtenberger, C.; Gangl, A.; Willheim, M.; Scheiner, O.; Steger, G. In Vivo Induction of HLA-DR on Human Neutrophils in Patients Treated with Interferon-Gamma. Blood 1996, 87, 3068. [Google Scholar] [CrossRef]
- Mesin, L.; Ersching, J.; Victora, G.D. Germinal Center B Cell Dynamics. Immunity 2016, 45, 471–482. [Google Scholar] [CrossRef] [Green Version]
- Cyster, J.G.; Allen, C.D.C. B Cell Responses: Cell Interaction Dynamics and Decisions. Cell 2019, 177, 524–540. [Google Scholar] [CrossRef] [Green Version]
- Li, G.-M.; Chiu, C.; Wrammert, J.; McCausland, M.; Andrews, S.F.; Zheng, N.-Y.; Lee, J.-H.; Huang, M.; Qu, X.; Edupuganti, S.; et al. Pandemic H1N1 Influenza Vaccine Induces a Recall Response in Humans That Favors Broadly Cross-Reactive Memory B Cells. Proc. Natl. Acad. Sci. USA 2012, 109, 9047–9052. [Google Scholar] [CrossRef] [Green Version]
- Henry, C.; Palm, A.-K.E.; Krammer, F.; Wilson, P.C. From Original Antigenic Sin to the Universal Influenza Virus Vaccine. Trends Immunol. 2018, 39, 70–79. [Google Scholar] [CrossRef]
- Sangster, M.Y.; Nguyen, P.Q.T.; Topham, D.J. Role of Memory B Cells in Hemagglutinin-Specific Antibody Production Following Human Influenza A Virus Infection. Pathogens 2019, 8, 167. [Google Scholar] [CrossRef] [Green Version]
- Margine, I.; Hai, R.; Albrecht, R.A.; Obermoser, G.; Harrod, A.C.; Banchereau, J.; Palucka, K.; Garcia-Sastre, A.; Palese, P.; Treanor, J.J.; et al. H3N2 Influenza Virus Infection Induces Broadly Reactive Hemagglutinin Stalk Antibodies in Humans and Mice. J. Virol. 2013, 87, 4728–4737. [Google Scholar] [CrossRef] [Green Version]
- Ellebedy, A.H.; Krammer, F.; Li, G.-M.; Miller, M.S.; Chiu, C.; Wrammert, J.; Chang, C.Y.; Davis, C.W.; McCausland, M.; Elbein, R.; et al. Induction of Broadly Cross-Reactive Antibody Responses to the Influenza HA Stem Region Following H5N1 Vaccination in Humans. Proc. Natl. Acad. Sci. USA 2014, 111, 13133–13138. [Google Scholar] [CrossRef] [Green Version]
- Halliley, J.L.; Tipton, C.M.; Liesveld, J.; Rosenberg, A.F.; Darce, J.; Gregoretti, I.V.; Popova, L.; Kaminiski, D.; Fucile, C.F.; Albizua, I.; et al. Long-Lived Plasma Cells Are Contained within the CD19−CD38hiCD138+ Subset in Human Bone Marrow. Immunity 2015, 43, 132–145. [Google Scholar] [CrossRef] [Green Version]
- Stadlbauer, D.; Rajabhathor, A.; Amanat, F.; Kaplan, D.; Masud, A.; Treanor, J.J.; Izikson, R.; Cox, M.M.; Nachbagauer, R.; Krammer, F. Vaccination with a Recombinant H7 Hemagglutinin-Based Influenza Virus Vaccine Induces Broadly Reactive Antibodies in Humans. mSphere 2017, 2, e00502–e00517. [Google Scholar] [CrossRef] [Green Version]
- Nachbagauer, R.; Feser, J.; Naficy, A.; Bernstein, D.I.; Guptill, J.; Walter, E.B.; Berlanda-Scorza, F.; Stadlbauer, D.; Wilson, P.C.; Aydillo, T.; et al. A Chimeric Hemagglutinin-Based Universal Influenza Virus Vaccine Approach Induces Broad and Long-Lasting Immunity in a Randomized, Placebo-Controlled Phase I Trial. Nat. Med. 2021, 27, 106–114. [Google Scholar] [CrossRef]
- Cirelli, K.M.; Crotty, S. Germinal Center Enhancement by Extended Antigen Availability. Curr. Opin. Immunol. 2017, 47, 64–69. [Google Scholar] [CrossRef]
- Tam, H.H.; Melo, M.B.; Kang, M.; Pelet, J.M.; Ruda, V.M.; Foley, M.H.; Hu, J.K.; Kumari, S.; Crampton, J.; Baldeon, A.D.; et al. Sustained Antigen Availability during Germinal Center Initiation Enhances Antibody Responses to Vaccination. Proc. Natl. Acad. Sci. USA 2016, 113, E6639–E6648. [Google Scholar] [CrossRef] [Green Version]
- Kemp, J.M.; Kajihara, M.; Nagahara, S.; Sano, A.; Brandon, M.; Lofthouse, S. Continuous Antigen Delivery from Controlled Release Implants Induces Significant and Anamnestic Immune Responses. Vaccine 2002, 20, 1089–1098. [Google Scholar] [CrossRef]
- Martin, J.T.; Cottrell, C.A.; Antanasijevic, A.; Carnathan, D.G.; Cossette, B.J.; Enemuo, C.A.; Gebru, E.H.; Choe, Y.; Viviano, F.; Fischinger, S.; et al. Targeting HIV Env Immunogens to B Cell Follicles in Nonhuman Primates through Immune Complex or Protein Nanoparticle Formulations. NPJ Vaccines 2020, 5, 72. [Google Scholar] [CrossRef]
- Schudel, A.; Francis, D.M.; Thomas, S.N. Material Design for Lymph Node Drug Delivery. Nat. Rev. Mater. 2019, 4, 415–428. [Google Scholar] [CrossRef]
- Topham, D.J.; Nguyen, P.; Sangster, M.Y. Pandemic Influenza Vaccines: What They Have Taught Us about B Cell Immunology. Curr. Opin. Immunol. 2018, 53, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, M.J.; Bernstein, D.I.; Winokur, P.; Rupp, R.; Anderson, E.; Rouphael, N.; Dickey, M.; Stapleton, J.T.; Edupuganti, S.; Spearman, P.; et al. Serological Responses to an Avian Influenza A/H7N9 Vaccine Mixed at the Point-of-Use with MF59 Adjuvant: A Randomized Clinical Trial. JAMA 2014, 312, 1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belshe, R.B.; Frey, S.E.; Graham, I.L.; Anderson, E.L.; Jackson, L.A.; Spearman, P.; Edupuganti, S.; Mulligan, M.J.; Rouphael, N.; Winokur, P.; et al. Immunogenicity of Avian Influenza A/Anhui/01/2005(H5N1) Vaccine with MF59 Adjuvant: A Randomized Clinical Trial. JAMA 2014, 312, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Jackson, L.A.; Campbell, J.D.; Frey, S.E.; Edwards, K.M.; Keitel, W.A.; Kotloff, K.L.; Berry, A.A.; Graham, I.; Atmar, R.L.; Creech, C.B.; et al. Effect of Varying Doses of a Monovalent H7N9 Influenza Vaccine with and Without AS03 and MF59 Adjuvants on Immune Response: A Randomized Clinical Trial. JAMA 2015, 314, 237. [Google Scholar] [CrossRef] [Green Version]
- Goji, N.A.; Nolan, C.; Hill, H.; Wolff, M.; Noah, D.L.; Williams, T.B.; Rowe, T.; Treanor, J.J. Immune Responses of Healthy Subjects to a Single Dose of Intramuscular Inactivated Influenza A/Vietnam/1203/2004 (H5N1) Vaccine after Priming with an Antigenic Variant. J. Infect. Dis. 2008, 198, 635–641. [Google Scholar] [CrossRef]
- Baer, J.; Santiago, F.; Yang, H.; Wu, H.; Holden-Wiltse, J.; Treanor, J.; Topham, D.J. B Cell Responses to H5 Influenza HA in Human Subjects Vaccinated with a Drifted Variant. Vaccine 2010, 28, 907–915. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S.F.; Chambers, M.J.; Schramm, C.A.; Plyler, J.; Raab, J.E.; Kanekiyo, M.; Gillespie, R.A.; Ransier, A.; Darko, S.; Hu, J.; et al. Activation Dynamics and Immunoglobulin Evolution of Pre-Existing and Newly Generated Human Memory B Cell Responses to Influenza Hemagglutinin. Immunity 2019, 51, 398–410.e5. [Google Scholar] [CrossRef]
- Khurana, S.; Verma, N.; Yewdell, J.W.; Hilbert, A.K.; Castellino, F.; Lattanzi, M.; Del Giudice, G.; Rappuoli, R.; Golding, H. MF59 Adjuvant Enhances Diversity and Affinity of Antibody-Mediated Immune Response to Pandemic Influenza Vaccines. Sci. Transl. Med. 2011, 3, 85ra48. [Google Scholar] [CrossRef] [Green Version]
- Khurana, S.; Coyle, E.M.; Manischewitz, J.; King, L.R.; Gao, J.; Germain, R.N.; Schwartzberg, P.L.; Tsang, J.S.; Golding, H.; The CHI Consortium. AS03-Adjuvanted H5N1 Vaccine Promotes Antibody Diversity and Affinity Maturation, NAI Titers, Cross-Clade H5N1 Neutralization, but Not H1N1 Cross-Subtype Neutralization. NPJ Vaccines 2018, 3, 40. [Google Scholar] [CrossRef] [Green Version]
- Galson, J.D.; Trück, J.; Kelly, D.F.; van der Most, R. Investigating the Effect of AS03 Adjuvant on the Plasma Cell Repertoire Following PH1N1 Influenza Vaccination. Sci. Rep. 2016, 6, 37229. [Google Scholar] [CrossRef]
- Mastelic Gavillet, B.; Eberhardt, C.S.; Auderset, F.; Castellino, F.; Seubert, A.; Tregoning, J.S.; Lambert, P.-H.; de Gregorio, E.; Del Giudice, G.; Siegrist, C.-A. MF59 Mediates Its B Cell Adjuvanticity by Promoting T Follicular Helper Cells and Thus Germinal Center Responses in Adult and Early Life. J. Immunol. 2015, 194, 4836–4845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, S.M.; Lou, M.; Yao, L.; Germain, R.N.; Radtke, A.J. The Lymph Node at a Glance—How Spatial Organization Optimizes the Immune Response. J. Cell Sci. 2020, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heesters, B.A.; van der Poel, C.E.; Das, A.; Carroll, M.C. Antigen Presentation to B Cells. Trends Immunol. 2016, 37, 844–854. [Google Scholar] [CrossRef]
- Mueller, S.N. Spreading the Load: Antigen Transfer between Migratory and Lymph Node-resident Dendritic Cells Promotes T-cell Priming. Eur. J. Immunol. 2017, 47, 1798–1801. [Google Scholar] [CrossRef] [Green Version]
- Crotty, S. T Follicular Helper Cell Biology: A Decade of Discovery and Diseases. Immunity 2019, 50, 1132–1148. [Google Scholar] [CrossRef]
- Tesini, B.L.; Kanagaiah, P.; Wang, J.; Hahn, M.; Halliley, J.L.; Chaves, F.A.; Nguyen, P.Q.T.; Nogales, A.; DeDiego, M.L.; Anderson, C.S.; et al. Broad Hemagglutinin-Specific Memory B Cell Expansion by Seasonal Influenza Virus Infection Reflects Early-Life Imprinting and Adaptation to the Infecting Virus. J. Virol. 2019, 93, e00169-19. [Google Scholar] [CrossRef] [Green Version]
- Moran, I.; Grootveld, A.K.; Nguyen, A.; Phan, T.G. Subcapsular Sinus Macrophages: The Seat of Innate and Adaptive Memory in Murine Lymph Nodes. Trends Immunol. 2019, 40, 35–48. [Google Scholar] [CrossRef]
- Angeletti, D.; Yewdell, J.W. Understanding and Manipulating Viral Immunity: Antibody Immunodominance Enters Center Stage. Trends Immunol. 2018, 39, 549–561. [Google Scholar] [CrossRef]
- Cobey, S.; Hensley, S.E. Immune History and Influenza Virus Susceptibility. Curr. Opin. Virol. 2017, 22, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. MRNA Vaccines—A New Era in Vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
| Adjuvants | Component | Vaccines | Trade Name | Use (Age Group) | Manufacturer |
|---|---|---|---|---|---|
| MF59 | Squalene; polysorbate 80; sorbitan trioleate | Seasonal influenza vaccine | FLUAD FLUAD Quadrivalent | 65 years and older | Novartis |
| A/H1N1 pandemic influenza vaccine | Forcetria Celtura | 6 months and older | Novartis | ||
| AS03 | Squalene; α-tocopherol; polysorbate 80 | A/H1N1 pandemic influenza vaccine | Pandemrix Prepandrix | 6 months and older 18 years and older | GlaxoSmithKline (GSK) |
| AF03 | Squalene; polyoxyethylene cetostearyl ether; mannitol; sorbitan oleate | A/H1N1 pandemic influenza vaccine | Humenza | 6 months and older | Sanofi |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen-Contant, P.; Sangster, M.Y.; Topham, D.J. Squalene-Based Influenza Vaccine Adjuvants and Their Impact on the Hemagglutinin-Specific B Cell Response. Pathogens 2021, 10, 355. https://doi.org/10.3390/pathogens10030355
Nguyen-Contant P, Sangster MY, Topham DJ. Squalene-Based Influenza Vaccine Adjuvants and Their Impact on the Hemagglutinin-Specific B Cell Response. Pathogens. 2021; 10(3):355. https://doi.org/10.3390/pathogens10030355
Chicago/Turabian StyleNguyen-Contant, Phuong, Mark Y. Sangster, and David J. Topham. 2021. "Squalene-Based Influenza Vaccine Adjuvants and Their Impact on the Hemagglutinin-Specific B Cell Response" Pathogens 10, no. 3: 355. https://doi.org/10.3390/pathogens10030355
APA StyleNguyen-Contant, P., Sangster, M. Y., & Topham, D. J. (2021). Squalene-Based Influenza Vaccine Adjuvants and Their Impact on the Hemagglutinin-Specific B Cell Response. Pathogens, 10(3), 355. https://doi.org/10.3390/pathogens10030355