
Development of a Colloidal Gold Immunochromatographic Assay for Duck Enteritis Virus Detection Using Monoclonal Antibodies

Abstract
:1. Introduction
2. Results
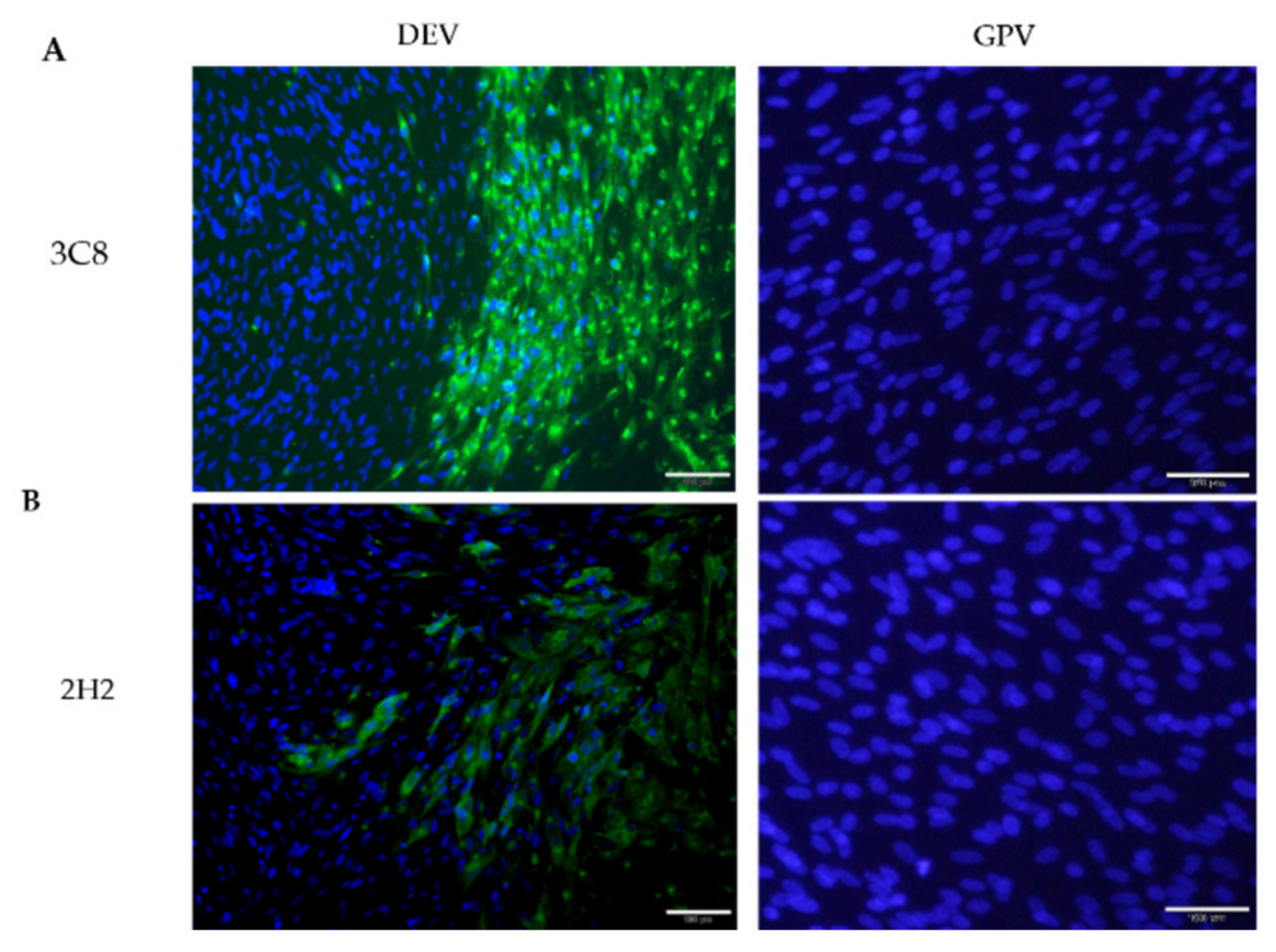
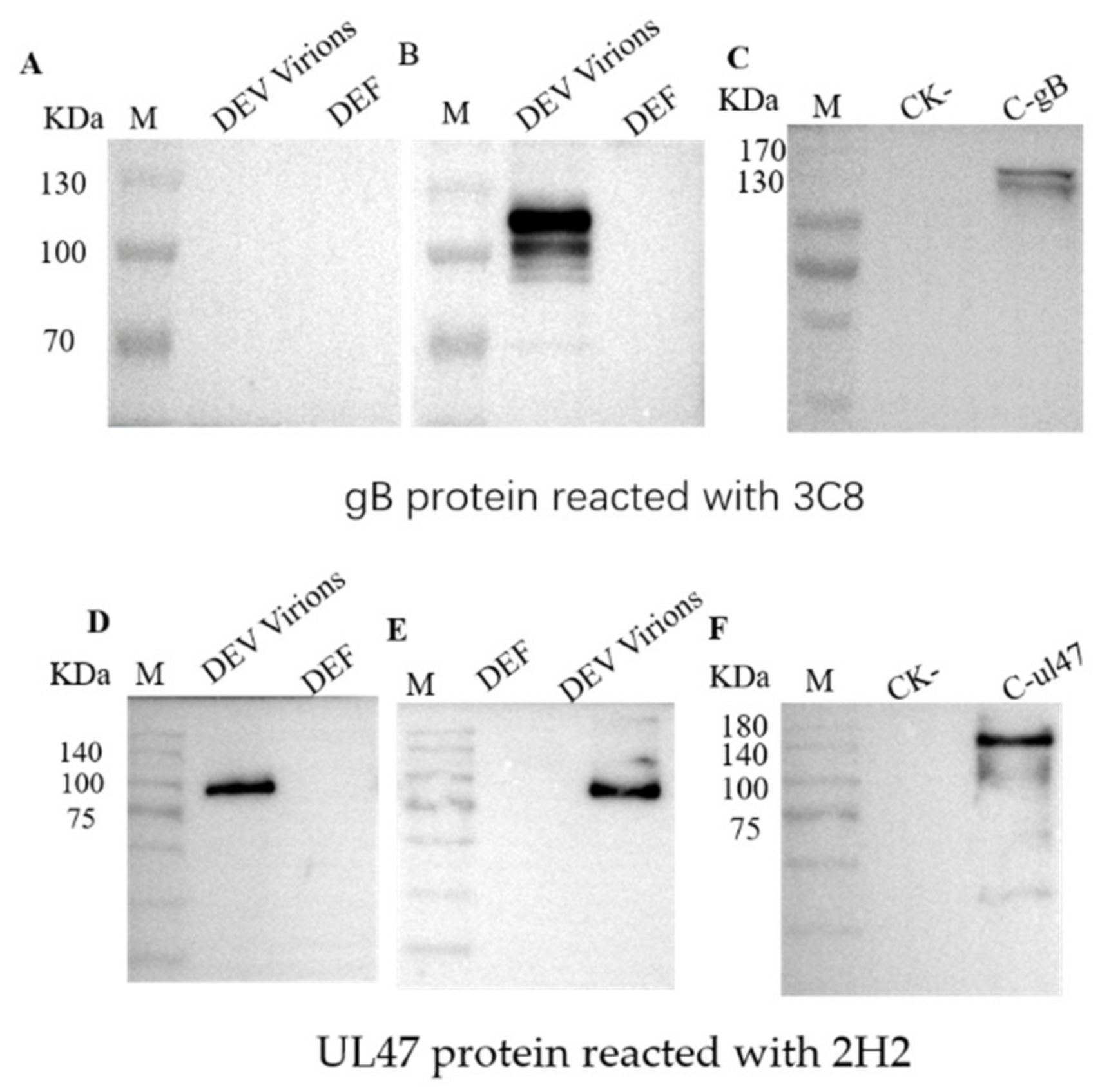
2.1. Production of Monoclonal Antibodies Against DEV
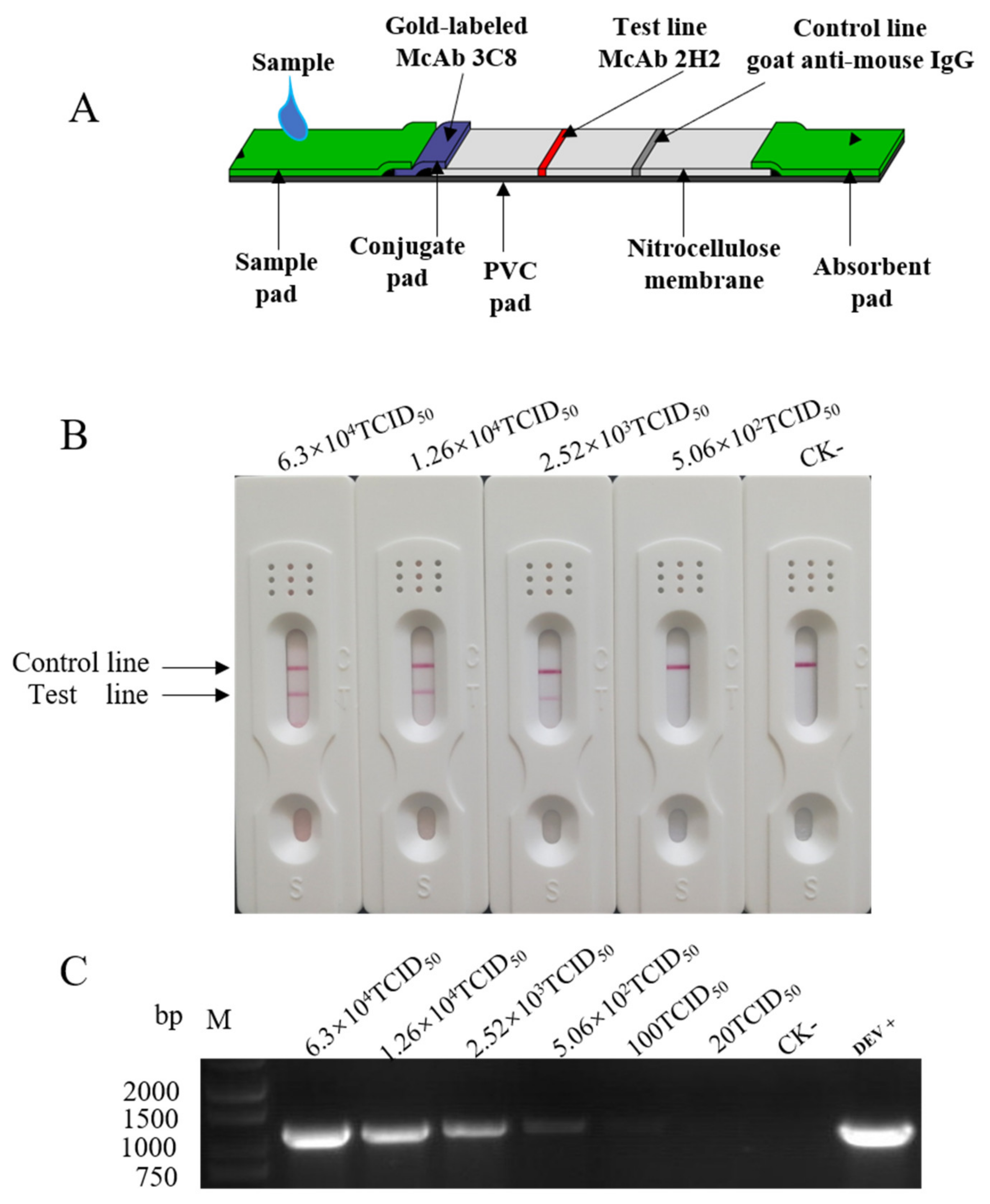
2.2. ICA Assembly, Sensitivity and Specificity Determination
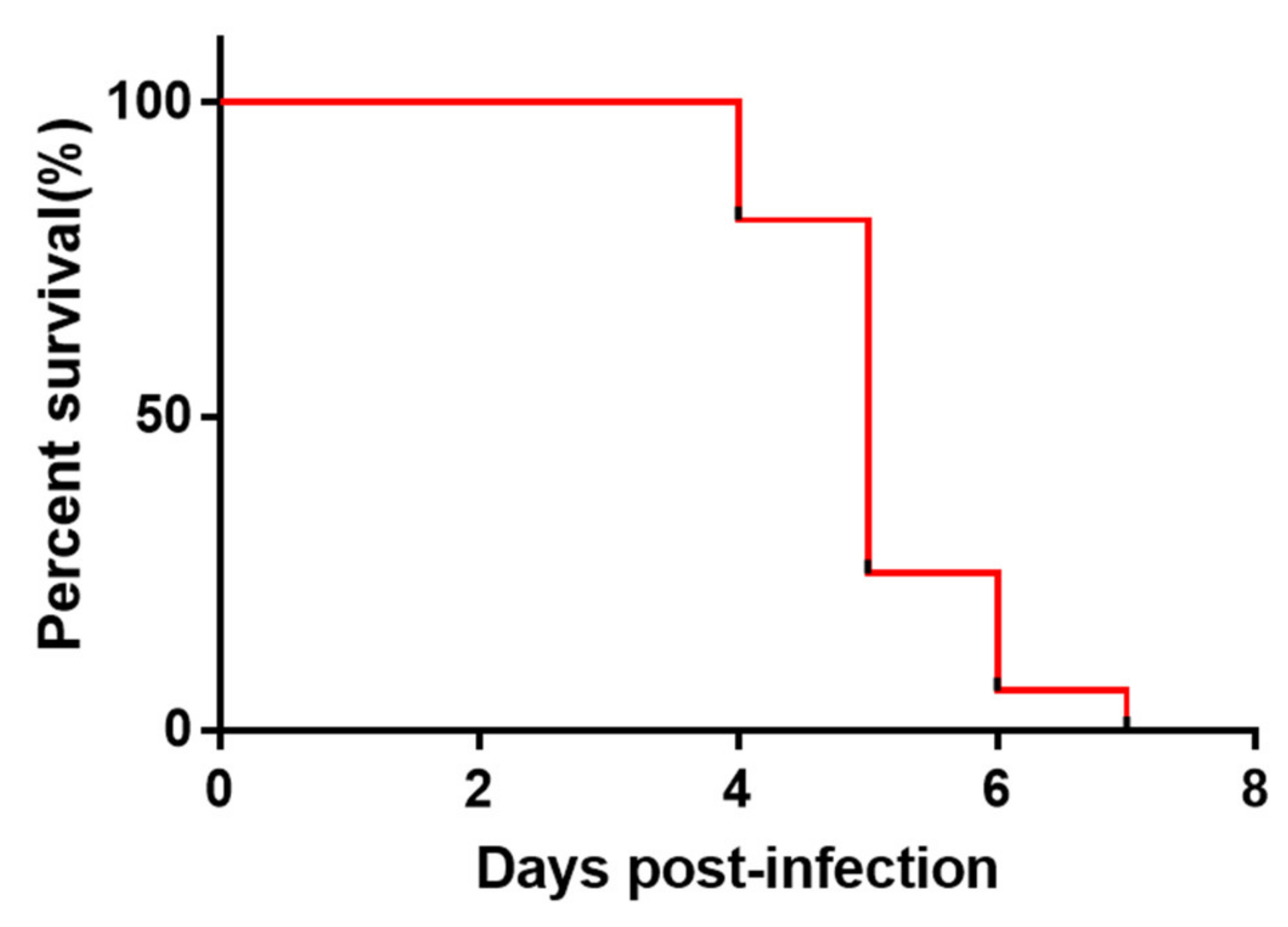
2.3. Validation of the ICA Strip Detection with Samples from Experimentally Inoculated Ducks
2.4. Stability of the ICA Strip
3. Discussion
4. Materials and Methods
4.1. Preparation and Titration of the DEV
4.2. Detection of the DEV Genome by PCR
4.3. Production of the Monoclonal Antibody
4.4. Immunofluorescence Assay
4.5. Western Blot and Mass Spectrometric Identification of the DEV Protein
4.6. Expression of the DEV Protein In Vitro
4.7. Double-Antibody Sandwich ELISA and the Conjugation of the MAb with Colloidal Gold
4.8. Determination of Specificity and Sensitivity of the Strip
4.9. Duck Infection and Sample Collection
4.10. Stability of the ICA Strip
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dhama, K.; Kumar, N.; Saminathan, M.; Tiwari, R.; Karthik, K.; Kumar, M.; Palanivelu, M.; Shabbir, M.; Malik, Y.; Singh, R. Duck virus enteritis (duck plague)—A comprehensive update. Vet. Q. 2017, 371, 57–80. [Google Scholar] [CrossRef]
- Li, Y.; Huang, B.; Ma, X.; Wu, J.; Li, F.; Ai, W.; Song, M.; Yang, H. Molecular characterization of the genome of duck enteritis virus. Virology 2009, 391, 151–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CHENG, A.C. Duck Plague, 1st ed.; China Agriculture Press: Beijing, China, 2015; pp. 3–4. [Google Scholar]
- Wang, G.; Qu, Y.; Wang, F.; Hu, D.; Liu, L.; Li, N.; Yue, R.; Li, C.; Liu, S. The comprehensive diagnosis and prevention of duck plague in northwest Shandong province of China. Poult. Sci. 2013, 9211, 2892–2898. [Google Scholar] [CrossRef]
- Diao, Y.; Lu, G.; Zheng, F.Y.; Yang, J.H.; Chen, Q.P.; Liu, Y.Z. Isolation and identification and characteristics of the pathogen of a new duck plague. Chin. J. Vet. Sci. 2006, 26, 136–139. [Google Scholar] [CrossRef]
- Plummer, P.; Alefantis, T.; Kaplan, S.; O’Connell, P.; Shawky, S.; Schat, K. Detection of duck enteritis virus by polymerase chain reaction. Avian Dis. 1998, 42, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Mandal, P.; Mukhopadhayay, S.; Pradhan, S.; Mondal, S.; Jana, C.; Patra, N.; Hansda, R. Development of nested polymerase chain reaction-based diagnosis of duck enteritis virus and detection of DNA polymerase gene from non-descriptive duck breeds of West Bengal, India. Vet. World 2017, 10, 336–341. [Google Scholar] [CrossRef] [Green Version]
- Jia, R.; Cheng, A.; Wang, M.; Qi, X.; Zhu, D.; Ge, H.; Luo, Q.; Liu, F.; Guo, Y.; Chen, X. Development and evaluation of an antigen-capture ELISA for detection of the UL24 antigen of the duck enteritis virus, based on a polyclonal antibody against the UL24 expression protein. J. Virol. Methods 2009, 161, 38–43. [Google Scholar] [CrossRef]
- Wu, Y.; Cheng, A.; Wang, M.; Zhang, S.; Chen, X. Serologic Detection of Duck Enteritis Virus Using an Indirect ELISA Based on Recombinant UL55 Protein. Avian Dis. 2011, 55, 626–632. [Google Scholar] [CrossRef]
- Koczula, K.; Gallotta, A.A. Lateral flow assays. Essays Biochem. 2016, 60, 111–120. [Google Scholar] [CrossRef]
- Siu, L.; Tsai, Y.; Lin, J.; Chen, T.; Fung, C.; Chang, F. Development of a Colloidal Gold-Based Immunochromatographic Strip for Rapid Detection of Klebsiella pneumoniae Serotypes K1 and K2. J. Clin. Microbiol. 2016, 54, 3018–3021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, D.; Yang, H.; Liu, C.; Zhao, K.; Li, J.; Deng, A. Ultrasensitive detection of Sudan I in food samples by a quantitative immunochromatographic assay. Food Chem. 2019, 277, 595–603. [Google Scholar] [CrossRef]
- Gas, F.; Baus, B.; Queré, J.; Chapelle, A.; Dreanno, C. Rapid detection and quantification of the marine toxic algae, Alexandrium minutum, using a super-paramagnetic immunochromatographic strip test. Talanta 2016, 147, 581–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Yang, J.; Zhang, G.; Wang, X.; Qiao, S.; Zhao, D.; Zhi, Y.; Li, X.; Xing, G.; Luo, J.; et al. Development of an immunochromatographic strip for the detection of antibodies against foot-and-mouth disease virus serotype O—ScienceDirect. J. Virol. Methods 2010, 165, 139–144. [Google Scholar] [CrossRef]
- Sun, Z.; Shi, B.; Meng, F.; Ma, R.; Hu, Q.; Qin, T.; Chen, S.; Peng, D.; Liu, X. Development of a Colloidal Gold-Based Immunochromatographic Strip for Rapid Detection of H7N9 Influenza Viruses. Front. Microbiol. 2018, 9, 2069. [Google Scholar] [CrossRef]
- Cheng, S.; Sun, J.; Yang, J.; Lv, J.Q.; Wu, F.; Lin, Y.X.; Liao, L.S.; Ye, Y.Y.; Cao, C.F.; Fang, L.R.; et al. A new immunoassay of serum antibodies against Peste des petits ruminants virus using quantum dots and a lateral-flow test strip. Anal. Bioanal. Chem. 2017, 409, 133–141. [Google Scholar] [CrossRef]
- Cooper, R.; Heldwein, E. Herpesvirus gB: A Finely Tuned Fusion Machine. Viruses 2015, 7, 6552–6569. [Google Scholar] [CrossRef] [Green Version]
- Newcomb, W.; Mingo, R.; Han, J.; Brown, J. Replication of herpes simplex virus: Egress of progeny virus at specialized cell membrane sites. J. Virol. 2012, 86, 7084–7097. [Google Scholar] [CrossRef] [Green Version]
- He, T.; Wang, M.; Cheng, A.; Yang, Q.; Jia, R.; Wu, Y.; Huang, J.; Chen, S.; Zhao, X.; Liu, M.; et al. Duck enteritis virus pUL47, as a late structural protein localized in the nucleus, mainly depends on residues 40 to 50 and 768 to 777 and inhibits IFN-β signalling by interacting with STAT1. Vet. Res. 2020, 51, 135. [Google Scholar] [CrossRef] [PubMed]
- Maeda, K.; Horimoto, T.; Norimine, J.; Kawaguchi, Y.; Tomonaga, K.; Niikura, M.; Kai, C.; Takahashi, E.; Mikami, T. Identification and nucleotide sequence of a gene in feline herpesvirus type 1 homologous to the herpes simplex virus gene encoding the glycoprotein B. Arch. Virol. 1992, 127, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, K.; Fujii, S.; Takada, A.; Kida, H. The amino-terminal residue of glycoprotein B is critical for neutralization of bovine herpesvirus 1. Virus Res. 2006, 115, 105–111. [Google Scholar] [CrossRef]
- C Vallbracht, M.; Brun, D.; Tassinari, M.; Vaney, M.; Pehau-Arnaudet, G.; Guardado-Calvo, P.; Haouz, A.; Klupp, B.; Mettenleiter, T.; Rey, F.; et al. Structure-Function Dissection of Pseudorabies Virus Glycoprotein B Fusion Loops. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen-de, W.; Min, L.; Ming, C.; Li-Ping, L.; Rui, W.; Hai-Lan, C.; Fu-Yan, C.; Qiang, M.; Wan-Wen, L.; Han-Zhong, C. Development of a colloidal gold immunochromatographic strip for rapid detection of Streptococcus agalactiae in tilapia. Biosens. Bioelectron. 2017, 91, 66–69. [Google Scholar] [CrossRef]
- Fenner, E.; Gibbs, E.; Murph, F.; Rott, R.; Studdert, M.; White, D. Veterinary Virology, 2nd ed.; Publisher: New York, NY, USA, 1993; pp. 339–373. [Google Scholar]
- Wang, H.H.; Hu, X.Y.; Su, J.L.; He, S.H.; Cheng, Y.Y.; Guo, Y. Isolation and identification of duck plague viruses from commercial meat-type ducks. Prev. Vet. Med. 2006, 28, 105–108. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method for estimating 50% endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Z.; Jin, H.; Hu, X.; Su, J. Development and application of a monoclonal antibody-based blocking ELISA for detection of antibodies to Tembusu virus in multiple poultry species. BMC Vet. Res. 2018, 14, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Grodzki, A.; Berenstein, E. Antibody purification: Ammonium sulfate fractionation or gel filtration. Methods Mol. Biol. 2010, 588, 15–26. [Google Scholar] [CrossRef]
- Asturias, J.A.; Arilla, M.C.; Aguirre, M.; Gómez-Bayón, N.; Martínez, J. Quantification of profilins by a monoclonal antibody-based sandwich ELISA. J. Immunol. Methods 1999, 229, 61–71. [Google Scholar] [CrossRef]
- Roth, J. The preparation of protein A-gold complexes with 3 nm and 15 nm gold particles and their use in labelling multiple antigens on ultra-thin sections. Histochem. J. 1982, 14, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Wang, X.; Zhi, A.; Bao, Y.; Yang, Y.; Qu, M.; Luo, J.; Li, Q.; Guo, J.; Wang, Z.; et al. Development of a lateral flow immunoassay strip for screening of sulfamonomethoxine residues. Food Addit. Contam. 2008, 25, 413–423. [Google Scholar] [CrossRef]
- Zeng, Y.; Liang, D.; Zheng, P.; Peng, T.; Sun, S.; Mari, G.; Jiang, H. Immunochromatographic fluorometric determination of clenbuterol with enhanced sensitivity. Mikrochim. Acta 2019, 186, 225. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days Post-Infectionš(dpi) | Control | Infected | |||
|---|---|---|---|---|---|
| PCR | ICA | PCR | ICA | Coincidence (%) | |
| 2 | 0/6 | 0/6 | 1/10 * | 0/10 | 93.7 |
| 3 | 0/6 | 0/6 | 10/10 | 8/10 | 87.5 |
| 4 | 0/6 | 0/6 | 10/10 | 10/10 | 100 |
| 5 | 0/6 | 0/6 | 5/5 | 5/5 | 100 |
| 6 | 0/6 | 0/6 | 3/3 | 3/3 | 100 |
| 7 | 0/6 | 0/6 | 1/1 | 1/1 | 100 |
| Total | 0/36 | 0/36 | 30/39 | 27/39 | 92.3 |
| Tissues | Sample Number | Positive Number | Coincidence (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| PCR | RICA | ||||||||
| Control | Infected | Control | Infected | Total * | Control | Infected | Total * | ||
| Heart | 5 | 5 | 0 | 5 | 5/10 | 0 | 2 | 2/10 | 70 |
| Liver | 5 | 10 | 0 | 10 | 10/15 | 2 | 10 | 12/15 | 86.7 |
| Spleen | 4 | 10 | 0 | 10 | 10/14 | 0 | 10 | 10/14 | 100 |
| Lung | 5 | 8 | 0 | 8 | 8/13 | 0 | 8 | 8/13 | 100 |
| Kidney | 4 | 10 | 0 | 10 | 10/14 | 0 | 10 | 14/14 | 100 |
| Brain | 4 | 5 | 0 | 5 | 5/9 | 0 | 2 | 2/9 | 66.7 |
| Bursa | 5 | 10 | 0 | 10 | 10/15 | 0 | 10 | 10/15 | 100 |
| Intestine | 4 | 10 | 0 | 10 | 10/14 | 0 | 10 | 10/14 | 100 |
| Total | 36 | 68 | 68/104 | 64/104 | 96.2 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.; Cao, Y.; Yan, M.; Sun, M.; Zhang, Q.; Wang, J.; Fu, G.; Liu, R.; Huang, Y.; Su, J. Development of a Colloidal Gold Immunochromatographic Assay for Duck Enteritis Virus Detection Using Monoclonal Antibodies. Pathogens 2021, 10, 365. https://doi.org/10.3390/pathogens10030365
Liu F, Cao Y, Yan M, Sun M, Zhang Q, Wang J, Fu G, Liu R, Huang Y, Su J. Development of a Colloidal Gold Immunochromatographic Assay for Duck Enteritis Virus Detection Using Monoclonal Antibodies. Pathogens. 2021; 10(3):365. https://doi.org/10.3390/pathogens10030365
Chicago/Turabian StyleLiu, Fengli, Yanxin Cao, Maokai Yan, Mengxu Sun, Qingshui Zhang, Jun Wang, Guanghua Fu, Rongchang Liu, Yu Huang, and Jingliang Su. 2021. "Development of a Colloidal Gold Immunochromatographic Assay for Duck Enteritis Virus Detection Using Monoclonal Antibodies" Pathogens 10, no. 3: 365. https://doi.org/10.3390/pathogens10030365
APA StyleLiu, F., Cao, Y., Yan, M., Sun, M., Zhang, Q., Wang, J., Fu, G., Liu, R., Huang, Y., & Su, J. (2021). Development of a Colloidal Gold Immunochromatographic Assay for Duck Enteritis Virus Detection Using Monoclonal Antibodies. Pathogens, 10(3), 365. https://doi.org/10.3390/pathogens10030365