
Molecular Characteristics of Carnivore protoparvovirus 1 with High Sequence Similarity between Wild and Domestic Carnivores in Taiwan

Abstract
:1. Introduction
2. Results
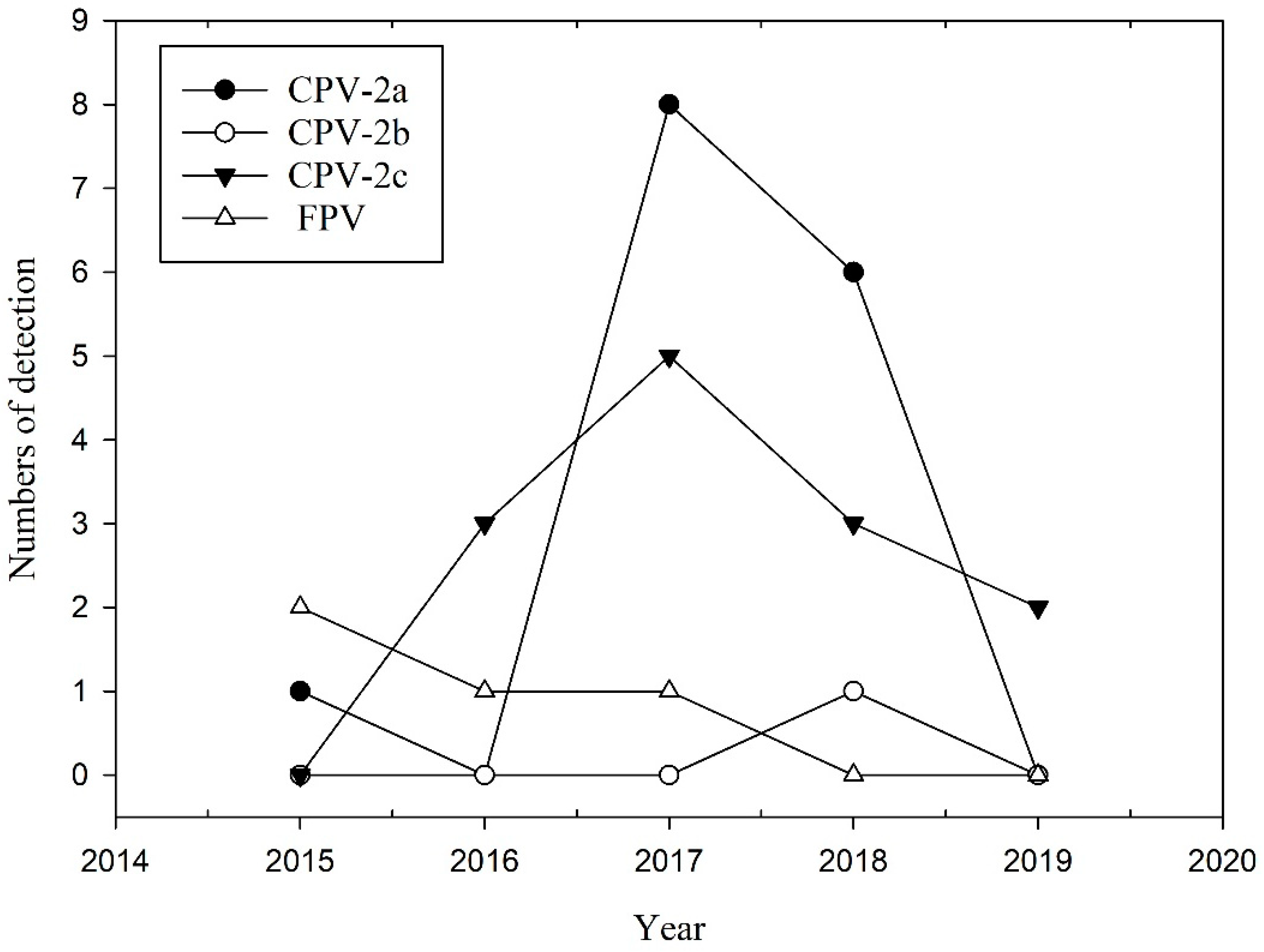
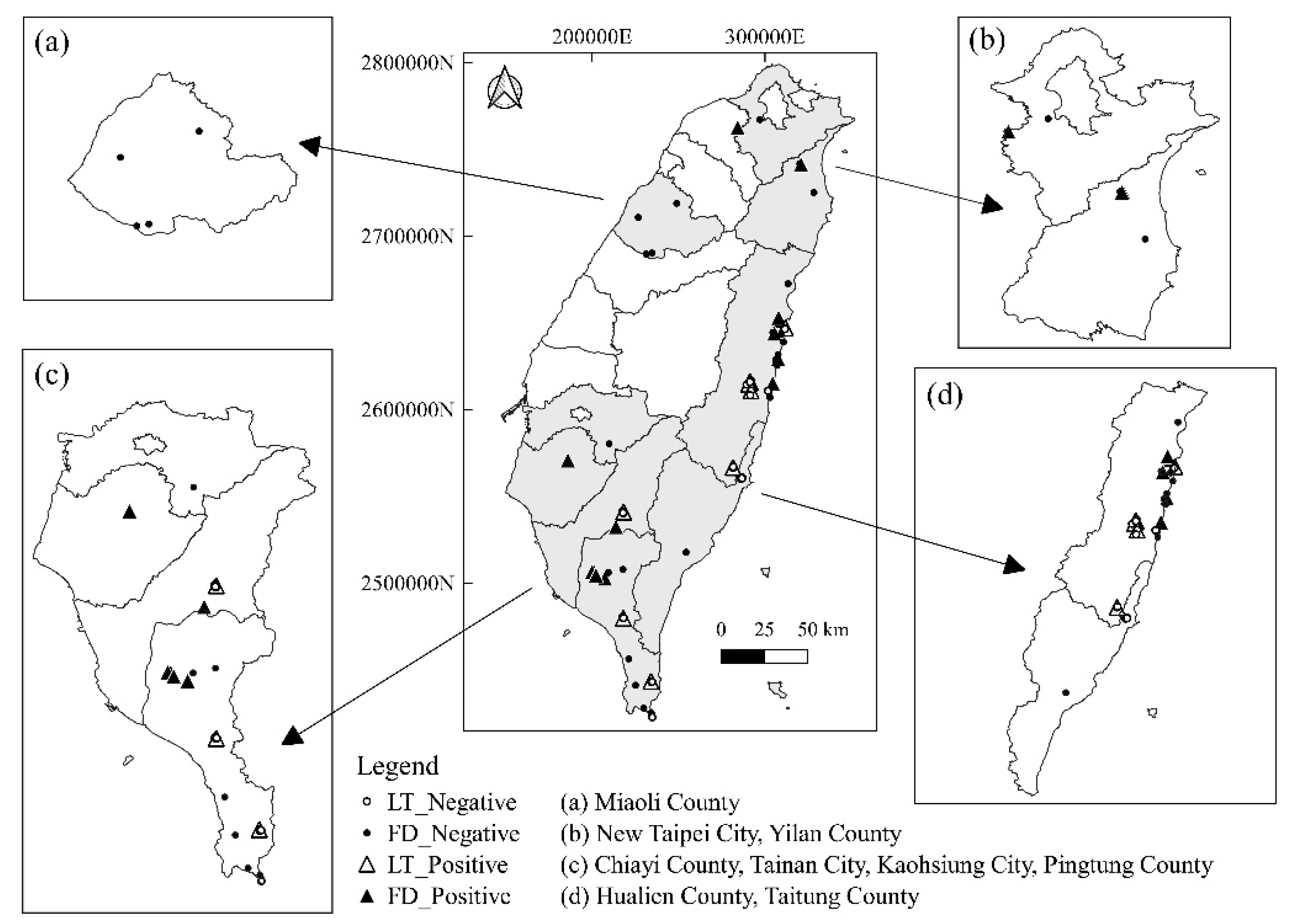
2.1. Prevalence of CPPV-1 in the Wild Carnivore Population
2.2. CPPV-1 Variants and Amino Acid Sequence Analysis in Wild Carnivores
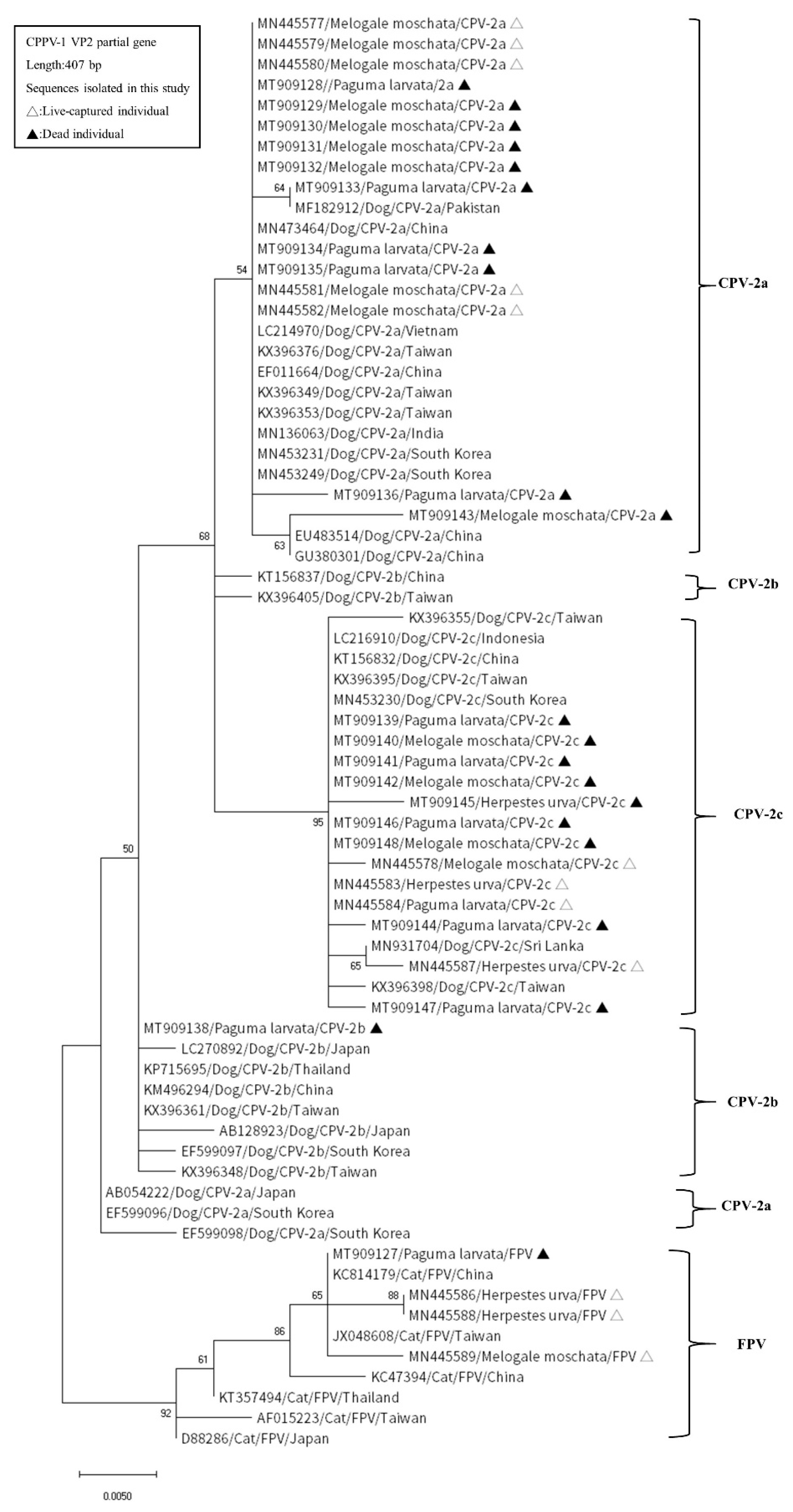
2.3. Phylogenetic Analysis of CPPV-1
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Sampling Area
4.3. Sample Collection
4.4. DNA Extraction, PCR Screening, and CPPV-1 Sequencing
4.5. Antigenic Type Characterization and Molecular Phylogenetic Analysis of CPPV-1
4.6. Genbank Sequence Submission
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Daniels, T.J.; Bekoff, M. Population and social biology of free-ranging dogs, Canis familiaris. J. Mammal. 1989, 70, 754–762. [Google Scholar] [CrossRef] [Green Version]
- Turner, D.C.; Bateson, P.; Bateson, P.P.G. The Domestic Cat: The Biology of Its Behaviour; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Allison, A.B.; Harbison, C.E.; Pagan, I.; Stucker, K.M.; Kaelber, J.T.; Brown, J.D.; Ruder, M.G.; Keel, M.K.; Dubovi, E.J.; Holmes, E.C. Role of multiple hosts in the cross-species transmission and emergence of a pandemic parvovirus. J. Virol. 2012, 86, 865–872. [Google Scholar] [CrossRef] [Green Version]
- Cleaveland, S. Epidemiology and control of rabies: The growing problem of rabies in Africa. Trans. R. Soc. Trop. Med. Hyg. 1998, 92, 131–134. [Google Scholar] [CrossRef]
- Cleaveland, S.; Appel, M.; Chalmers, W.; Chillingworth, C.; Kaare, M.; Dye, C. Serological and demographic evidence for domestic dogs as a source of canine distemper virus infection for Serengeti wildlife. Vet. Microbiol. 2000, 72, 217–227. [Google Scholar] [CrossRef]
- Battilani, M.; Scagliarini, A.; Tisato, E.; Turilli, C.; Jacoboni, I.; Casadio, R.; Prosperi, S. Analysis of canine parvovirus sequences from wolves and dogs isolated in Italy. J. Gen. Virol. 2001, 82, 1555–1560. [Google Scholar] [CrossRef]
- Ndiana, L.A.; Lanave, G.; Desario, C.; Berjaoui, S.; Alfano, F.; Puglia, I.; Fusco, G.; Colaianni, M.L.; Vincifori, G.; Camarda, A. Circulation of diverse protoparvoviruses in wild carnivores, Italy. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Danner, R.M.; Goltz, D.M.; Hess, S.C.; Banko, P.C. Evidence of feline immunodeficiency virus, feline leukemia virus, and Toxoplasma gondii in feral cats on Mauna Kea, Hawaii. J. Wildl. Dis. 2007, 43, 315–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appel, M.; Scott, F.; Carmichael, L. Isolation and immunisation studies of a canine parco-like virus from dogs with haemorrhagic enteritis. Vet. Rec. 1979, 105, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Oğuzoğlu, T.; Muz, D.; Timurkan, M.; Maral, N.; Gurcan, I. Prevalences of feline coronavirus (FCoV), feline leukaemia virus (FeLV), feline immunodeficiency virus (FIV) and feline parvovirus (FPV) among domestic cats in Ankara, Turkey. Rev. Méd. Vét. 2013, 164, 511–516. [Google Scholar]
- Decaro, N.; Buonavoglia, C. Canine parvovirus—a review of epidemiological and diagnostic aspects, with emphasis on type 2c. Vet. Microbiol. 2012, 155, 1–12. [Google Scholar] [CrossRef]
- Allison, A.B.; Kohler, D.J.; Ortega, A.; Hoover, E.A.; Grove, D.M.; Holmes, E.C.; Parrish, C.R. Host-specific parvovirus evolution in nature is recapitulated by in vitro adaptation to different carnivore species. PLoS Pathog. 2014, 10, e1004475. [Google Scholar] [CrossRef]
- Maclachlan, N.J.; Dubovi, E.J. Fenner’s Veterinary Virology; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Woodroffe, R.; Prager, K.C.; Munson, L.; Conrad, P.A.; Dubovi, E.J.; Mazet, J.A. Contact with domestic dogs increases pathogen exposure in endangered African wild dogs (Lycaon pictus). PLoS ONE 2012, 7, e30099. [Google Scholar] [CrossRef]
- Yang, D.-K.; Yoon, S.-S.; Byun, J.-W.; Lee, K.-W.; Oh, Y.-I.; Song, J.-Y. Serological survey for canine parvovirus type 2a (CPV-2a) in the stray dogs in South Korea. J. Bacteriol. Virol. 2010, 40, 77–81. [Google Scholar] [CrossRef] [Green Version]
- DiGangi, B.A.; Levy, J.K.; Griffin, B.; McGorray, S.P.; Dubovi, E.J.; Dingman, P.A.; Tucker, S.J. Prevalence of serum antibody titers against feline panleukopenia virus, feline herpesvirus 1, and feline calicivirus in cats entering a Florida animal shelter. J. Am. Vet. Med Assoc. 2012, 241, 1320–1325. [Google Scholar] [CrossRef]
- Cotmore, S.F.; Agbandje-McKenna, M.; Chiorini, J.A.; Mukha, D.V.; Pintel, D.J.; Qiu, J.; Soderlund-Venermo, M.; Tattersall, P.; Tijssen, P.; Gatherer, D. The family parvoviridae. Arch. Virol. 2014, 159, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Steinel, A.; Parrish, C.R.; Bloom, M.E.; Truyen, U. Parvovirus infections in wild carnivores. J. Wildl. Dis. 2001, 37, 594–607. [Google Scholar] [CrossRef]
- Mech, L.D.; Kurtz, H.J.; Goyal, S. Death of a wild wolf from canine parvoviral enteritis. J. Wildl. Dis. 1997, 33, 321–322. [Google Scholar] [CrossRef] [PubMed]
- Bastianello, S.S. Canine parvovirus myocarditis: Clinical signs and pathological lesions encountered in natural cases. J. South Afr. Vet. Assoc. 1981, 52, 105–108. [Google Scholar]
- Nho, W.-G.; Sur, J.-H.; Doster, A.R.; Kim, S.-B. Detection of canine parvovirus in naturally infected dogs with enteritis and myocarditis by in situ hybridization. J. Vet. Diagn. Investig. 1997, 9, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Creel, S.; Creel, N.M.; Munson, L.; Sanderlin, D.; Appel, M.J. Serosurvey for selected viral diseases and demography of African wild dogs in Tanzania. J. Wildl. Dis. 1997, 33, 823–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtis Pei, M.-T.C. Present Status and Conservation of Small Carnivores at Low Elevation Mountains in Shinchu County and Miaoli County (3/3). For. Bur. Cons. Res. 2008, 9, 1–88. [Google Scholar]
- Hsu, Y.; Liu Severinghaus, L.; Serpell, J.A. Dog keeping in Taiwan: Its contribution to the problem of free-roaming dogs. J. Appl. Anim. Welf. Sci. 2003, 6, 1–23. [Google Scholar] [CrossRef]
- Chen, C.-C.; Chang, A.-M.; Wada, T.; Chen, M.-T.; Tu, Y.-S. Distribution of Carnivore protoparvovirus 1 in free-living leopard cats (Prionailurus bengalensis chinensis) and its association with domestic carnivores in Taiwan. PLoS ONE 2019, 14, e0221990. [Google Scholar] [CrossRef] [Green Version]
- Chiang, S.-Y.; Wu, H.-Y.; Chiou, M.-T.; Chang, M.-C.; Lin, C.-N. Identification of a novel canine parvovirus type 2c in Taiwan. Virol. J. 2016, 13, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.-C.; Chen, W.-D.; Lin, S.-L.; Chan, J.P.-W.; Wong, M.-L. Phylogenetic analysis of canine parvovirus VP2 gene in Taiwan. Virus Genes 2005, 31, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Decaro, N.; Buonavoglia, C. Evolution of CPV-2 and implicance for antigenic/genetic characterization. Virus Genes 2006, 33, 11–13. [Google Scholar] [CrossRef]
- Bagshaw, C.; Isdell, A.E.; Thiruvaiyaru, D.S.; Brisbin Jr, I.L.; Sanchez, S. Molecular detection of canine parvovirus in flies (Diptera) at open and closed canine facilities in the eastern United States. Prev. Vet. Med. 2014, 114, 276–284. [Google Scholar] [CrossRef]
- Behdenna, A.; Lembo, T.; Calatayud, O.; Cleaveland, S.; Halliday, J.E.; Packer, C.; Lankester, F.; Hampson, K.; Craft, M.E.; Czupryna, A. Transmission ecology of canine parvovirus in a multi-host, multi-pathogen system. Proc. R. Soc. B 2019, 286, 20182772. [Google Scholar] [CrossRef] [Green Version]
- Murphy, F.A.; Gibbs, E.P.J.; Horzinek, M.C.; Studdert, M.J. Veterinary Virology; Elsevier: Amsterdam, The Netherlands, 1999. [Google Scholar]
- Belsare, A.; Vanak, A.T.; Gompper, M. Epidemiology of viral pathogens of free-ranging dogs and Indian foxes in a human-dominated landscape in central India. Transbound. Emerg. Dis. 2014, 61, 78–86. [Google Scholar] [CrossRef]
- de Almeida Curi, N.H.; Massara, R.L.; de Oliveira Paschoal, A.M.; Soriano-Araújo, A.; Lobato, Z.I.P.; Demétrio, G.R.; Chiarello, A.G.; Passamani, M. Prevalence and risk factors for viral exposure in rural dogs around protected areas of the Atlantic forest. Bmc Vet. Res. 2016, 12, 21. [Google Scholar]
- Holmala, K.; Kauhala, K. Habitat use of medium-sized carnivores in southeast Finland—Key habitats for rabies spread? Ann. Zool. Fenn. 2009, 46, 233–246. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Zhu, Q.; Diao, F.; Teng, C.; Peng, H.; Shang, Y.; Zhao, Y.; Wang, J.; Jiang, S.; Xie, Z. Emergence of novel canine parvovirus type 2 and its pathogenesis in raccoon dogs. Vet. Microbiol. 2018, 216, 7–12. [Google Scholar]
- Calatayud, O.; Esperón, F.; Velarde, R.; Oleaga, Á.; Llaneza, L.; Ribas, A.; Negre, N.; de la Torre, A.; Rodríguez, A.; Millán, J. Genetic characterization of Carnivore Parvoviruses in Spanish wildlife reveals domestic dog and cat-related sequences. Transbound. Emerg. Dis. 2020, 67, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.-L. The Study of Dog Diseases in Public Shelter Yuan; Council of Agriculture of the Executive Yuan: Taiwan, 2006. [Google Scholar]
- Chaiyasak, S.; Piewbang, C.; Banlunara, W.; Techangamsuwan, S. Carnivore protoparvovirus-1 associated with an outbreak of hemorrhagic gastroenteritis in small Indian civets. Vet. Pathol. 2020, 57, 706–713. [Google Scholar] [CrossRef]
- Gjeltema, J.; Murphy, H.; Rivera, S. Clinical canine parvovirus type 2C infection in a group of Asian small-clawed otters (Aonyx cinerea). J. Zoo Wildl. Med. 2015, 46, 120–123. [Google Scholar] [CrossRef]
- Brockie, R.E.; Sadleir, R.M.; Linklater, W.L. Long-term wildlife road-kill counts in New Zealand. N. Z. J. Zool. 2009, 36, 123–134. [Google Scholar] [CrossRef]
- Ikeda, Y.; Miyazawa, T.; Nakamura, K.; Naito, R.; Inoshima, Y.; Tung, K.C.; Lee, W.M.; Chen, M.C.; Kuo, T.F.; Lin, J.A.; et al. Serosurvey for selected virus infections of wild carnivores in Taiwan and Vietnam. J. Wildl. Dis. 1999, 35, 578–581. [Google Scholar] [CrossRef] [Green Version]
- Xinyu, T.; Min, C.S.; Yifan, W.; Lien, S.M.; Chan, A.; Hui, T.X.; Lee, B.; Yelin, W.; Chia-Da, H.; Oh, S. Canine parvovirus-2c (CPV-2c) infection in wild Asian Palm Civets (Paradoxurus hermaphroditus) in Singapore. J. Wildl. Dis. 2019, 55, 965–969. [Google Scholar] [CrossRef] [PubMed]
- Calatayud, O.; Esperón, F.; Cleaveland, S.; Biek, R.; Keyyu, J.; Eblate, E.; Neves, E.; Lembo, T.; Lankester, F. Carnivore parvovirus ecology in the Serengeti ecosystem: Vaccine strains circulating and new host species identified. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Kaelber, J.T.; Demogines, A.; Harbison, C.E.; Allison, A.B.; Goodman, L.B.; Ortega, A.N.; Sawyer, S.L.; Parrish, C.R. Evolutionary reconstructions of the transferrin receptor of Caniforms supports canine parvovirus being a re-emerged and not a novel pathogen in dogs. PLoS Pathog 2012, 8, e1002666. [Google Scholar] [CrossRef] [Green Version]
- Allison, A.B.; Parrish, C.R. Parvoviruses of carnivores: Their transmission and the variation of viral host range. In The Role of Animals in Emerging Viral Diseases; Elsevier: Amsterdam, The Netherlands, 2014; pp. 39–61. [Google Scholar]
- Barr, M.C. Feline Parvovirus. Clin. Small Anim. Intern. Med. 2020, 869–871. [Google Scholar]
- Ikeda, Y.; Nakamura, K.; Miyazawa, T.; Tohya, Y.; Takahashi, E.; Mochizuki, M. Feline host range of canine parvovirus: Recent emergence of new antigenic types in cats. Emerg. Infect. Dis. 2002, 8, 341. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.F.; Sgro, J.Y.; Parrish, C.R. Multiple amino acids in the capsid structure of canine parvovirus coordinately determine the canine host range and specific antigenic and hemagglutination properties. J. Virol. 1992, 66, 6858–6867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, K.; Lee, S.-M.; Kim, Y.; Kim, Y.-K.; Lee, S.-Y.; Jheong, W.-H.; Oem, J.-K. Genetic Characterization of Canine Parvovirus Type 2 Detected in Wild Raccoon Dogs (Nyctereutes procyonoides) in the Republic of Korea. J. Wildl. Dis. 2019, 55, 512–515. [Google Scholar]
- Chang, W.; Chang, A.; Pan, M. Antigenic Types of Canine Parvoviruses Prevailing in Taiwan. Vet. Rec. 1996, 138, 447. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-C.; Chiang, S.-Y.; Wu, H.-Y.; Lin, J.-H.; Chiou, M.-T.; Liu, H.-F.; Lin, C.-N. Phylodynamic and Genetic Diversity of Canine Parvovirus Type 2c in Taiwan. Int. J. Mol. Sci. 2017, 18, 2703. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lin, P.; Zhao, H.; Cheng, Y.; Jiang, Z.; Zhu, H.; Wu, H.; Cheng, S. Continuing evolution of canine parvovirus in China: Isolation of novel variants with an Ala5Gly mutation in the VP2 protein. Infect. Genet. Evol. 2016, 38, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Jeoung, S.-Y.; Ahn, S.-J.; Kim, D. Genetic analysis of VP2 gene of canine parvovirus isolates in Korea. J. Vet. Med. Sci. 2008, 70, 719–722. [Google Scholar] [CrossRef] [Green Version]
- Phromnoi, S.; Sirinarumitr, K.; Sirinarumitr, T. Sequence analysis of VP2 gene of canine parvovirus isolates in Thailand. Virus Genes 2010, 41, 23–29. [Google Scholar] [CrossRef]
- Soma, T.; Taharaguchi, S.; Ohinata, T.; Ishii, H.; Hara, M. Analysis of the VP2 protein gene of canine parvovirus strains from affected dogs in Japan. Res. Vet. Sci. 2013, 94, 368–371. [Google Scholar] [CrossRef]
- Lin, C.-N.; Chien, C.-H.; Chiou, M.-T.; Chueh, L.-L.; Hung, M.-Y.; Hsu, H.-S. Genetic characterization of type 2a canine parvoviruses from Taiwan reveals the emergence of an Ile324 mutation in VP2. Virol. J. 2014, 11, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, H.; Matta, S.L.; Amsaveni, S.; Antony, P.; Thanislass, J.; Pillai, R. Phylogenetic analysis of canine parvovirus partial VP2 gene in India. Virus Genes 2014, 48, 89–95. [Google Scholar] [CrossRef]
- Mira, F.; Purpari, G.; Lorusso, E.; Di Bella, S.; Gucciardi, F.; Desario, C.; Macaluso, G.; Decaro, N.; Guercio, A. Introduction of Asian canine parvovirus in Europe through dog importation. Transbound. Emerg. Dis. 2018, 65, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Cságola, A.; Varga, S.; Lőrincz, M.; Tuboly, T. Analysis of the full-length VP2 protein of canine parvoviruses circulating in Hungary. Arch. Virol. 2014, 159, 2441–2444. [Google Scholar] [CrossRef] [PubMed]
- Mira, F.; Purpari, G.; Di Bella, S.; Colaianni, M.L.; Schirò, G.; Chiaramonte, G.; Gucciardi, F.; Pisano, P.; Lastra, A.; Decaro, N. Spreading of canine parvovirus type 2c mutants of Asian origin in southern Italy. Transbound. Emerg. Dis. 2019, 66, 2297–2304. [Google Scholar] [CrossRef]
- Steinel, A.; Munson, L.; Van Vuuren, M.; Truyen, U. Truyen. Genetic characterization of feline parvovirus sequences from various carnivores. J. Gen. Virol. 2000, 81, 345–350. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucl. Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+ C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Family, Species | Common Name | Sample Type | Age | Sex | Total | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Adult | Subadult | Juvenile | ND 1 | Female | Male | ND 1 | ||||
| Herpestidae, Herpestes urva | Crab-eating mongoose | LT 2 | 53 | 25 | 0 | 2 | 43 | 36 | 1 | 80 |
| FD 3 | 5 | 0 | 1 | 0 | 1 | 2 | 1 | 6 | ||
| Mustelidae, Melogale moschata | Chinese ferret badger | LT | 25 | 5 | 0 | 2 | 11 | 18 | 3 | 32 |
| FD | 29 | 1 | 0 | 1 | 15 | 16 | 0 | 31 | ||
| Viverridae, Paguma larvata | Masked palm civet | LT | 4 | 1 | 0 | 1 | 2 | 3 | 1 | 6 |
| FD | 13 | 4 | 0 | 9 | 8 | 10 | 8 | 26 | ||
| Species | Sample Type | No. Individuals | No. Positive | Prevalence | 95% CI | Variants | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Lower | Upper | FPV | CPV-2a | CPV-2b | CPV-2c | |||||
| Crab-eating mongoose | LT 1 | 80 | 4 | 5.0% | 0.2% | 9.8% | 2 | 0 | 0 | 2 |
| FD 2 | 6 | 1 | 16.7% | 0.0% | 46.5% | 0 | 0 | 0 | 1 | |
| Sum | 86 | 5 | 5.8% | 0.9% | 10.8% | |||||
| Masked palm civet | LT | 6 | 1 | 16.7% | 0.0% | 46.5% | 0 | 0 | 0 | 1 |
| FD | 26 | 11 3 | 42.3% | 23.3% | 61.3% | 1 | 5 | 1 | 5 | |
| Sum | 32 | 12 | 37.5% | 20.7% | 54.3% | |||||
| Chinese ferret badger | LT | 32 | 7 | 21.9% | 0.2% | 9.8% | 1 | 4 | 0 | 1 |
| FD | 31 | 8 | 25.8% | 10.4% | 41.2% | 0 | 5 | 0 | 3 | |
| Sum | 63 | 15 | 23.8% | 13.3% | 34.3% | |||||
| Total | 181 | 32 | 17.7% | 12.1% | 23.2% | 4 | 15 | 1 | 13 | |
| aaSTs 1 | Viral Variant | n2 | aa Position (Nucleotide Position) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 303 (907–909) | 305 (913–915) | 308 (923–924) | 309 (925–927) | 310 (928–930) | 323 (967–969) | 335 (1003–1005) | 360 (1078–1080) | 373 (1117–1119) | 400 (1198–1200) | 411 (1231–1233) | 426 (1276–1278) | 429 (1285–1287) | |||
| A | CPV-2a | 12 | Phe (TTT) | Tyr (TAT) | Val (GTT) | Gln (CAA) | Gln (CAA) | Asn (AAC) | Glu (GAG) | Gly (GGA) | Asp (GAT) | Tyr (TAT) | Glu (GAA) | Asn (AAT) | Val (GTA) |
| B | CPV-2a | 1 | Phe (TTT) | Tyr (TAT) | Val (GTT) | Gln (CAA) | Gln (CAA) | Asn (AAC) | Glu (GAG) | Gly (GGA) | Gly (GGT) | Tyr (TAT) | Glu (GAA) | Asn (AAT) | Val (GTA) |
| C | CPV-2a | 1 | Phe (TTT) | Tyr (TAT) | Val (GTT) | Arg (CGA) | Gln (CAA) | Asn (AAC) | Glu (GAG) | Arg (AGA) | Asp (GAT) | Tyr (TAT) | Glu (GAA) | Asn (AAT) | Val (GTA) |
| D | CPV-2b | 1 | Phe (TTT) | Tyr (TAT) | Val (GTT) | Gln (CAA) | Gln (CAA) | Asn (AAC) | Glu (GAG) | Gly (GGA) | Asp (GAT) | Tyr (TAT) | Glu (GAA) | Asp (GAT) | Val (GTA) |
| E | CPV-2c | 11 | Phe (TTT) | Tyr (TAT) | Val (GTT) | Gln (CAA) | Gln (CAA) | Asn (AAC) | Glu (GAG) | Gly (GGA) | Asp (GAT) | Tyr (TAT) | Glu (GAA) | Glu (GAA) | Val (GTA) |
| F | CPV-2c | 1 | Phe (TTT) | Tyr (TAT) | Val (GTT) | Gln (CAA) | His (CAT) | Asn (AAC) | Glu (GAG) | Gly (GGA) | Asp (GAT) | Tyr (TAT) | Glu (GAA) | Glu (GAA) | Val (GTA) |
| G | CPV-2c | 1 | Phe (TTT) | Tyr (TAT) | Val (GTT) | Gln (CAA) | Gln (CAA) | Asn (AAC) | Glu (GAG) | Gly (GGA) | Asp (GAT) | Asn (AAT) | Glu (GAA) | Glu (GAA) | Val (GTA) |
| H | FPV | 2 | Phe (TTT) | Asp (GAT) | Val (GTT) | Gln (CAA) | Gln (CAA) | Asp (GAC) | Glu (GAG) | Gly (GGA) | Asp (GAT) | Tyr (TAT) | Glu (GAA) | Asn (AAT) | Val (GTA) |
| I | FPV | 1 | Phe (TTT) | Asp (AAT) | Val (GTT) | Gln (CAA) | Gln (CAA) | Asp (GAC) | Gly (GGG) | Gly (GGA) | Asp (GAT) | Tyr (TAT) | Glu (GAA) | Asn (AAT) | Val (GTA) |
| J | FPV | 1 | Phe (TTT) | Asn (GAT) | Val (GTT) | Gln (CAA) | Gln (CAA) | Asp (GAC) | Glu (GAG) | Gly (GGA) | Asp (GAT) | Tyr (TAT) | Lys (AAA) | Asn (AAT) | Val (GTA) |
| aaSTs 1 | Viral Variant | aaSTs | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | D | E | F | G | H | I | J | ||
| A | CPV-2a | ||||||||||
| B | CPV-2a | 0.00738 | |||||||||
| C | CPV-2a | 0.01474 | 0.02224 | ||||||||
| D | CPV-2b | 0.01485 | 0.02241 | 0.02985 | |||||||
| E | CPV-2c | 0.02263 | 0.03035 | 0.03789 | 0.02997 | ||||||
| F | CPV-2c | 0.02258 | 0.03027 | 0.03780 | 0.02989 | 0.01480 | |||||
| G | CPV-2c | 0.01503 | 0.02267 | 0.03019 | 0.02238 | 0.00738 | 0.00737 | ||||
| H | FPV | 0.02235 | 0.02997 | 0.03743 | 0.02231 | 0.04595 | 0.04584 | 0.03814 | |||
| I | FPV | 0.02995 | 0.03766 | 0.04515 | 0.02990 | 0.05389 | 0.05376 | 0.04601 | 0.00740 | ||
| J | FPV | 0.02993 | 0.03762 | 0.04511 | 0.02987 | 0.05384 | 0.05370 | 0.04596 | 0.01479 | 0.02230 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, A.-M.; Chen, C.-C. Molecular Characteristics of Carnivore protoparvovirus 1 with High Sequence Similarity between Wild and Domestic Carnivores in Taiwan. Pathogens 2021, 10, 671. https://doi.org/10.3390/pathogens10060671
Chang A-M, Chen C-C. Molecular Characteristics of Carnivore protoparvovirus 1 with High Sequence Similarity between Wild and Domestic Carnivores in Taiwan. Pathogens. 2021; 10(6):671. https://doi.org/10.3390/pathogens10060671
Chicago/Turabian StyleChang, Ai-Mei, and Chen-Chih Chen. 2021. "Molecular Characteristics of Carnivore protoparvovirus 1 with High Sequence Similarity between Wild and Domestic Carnivores in Taiwan" Pathogens 10, no. 6: 671. https://doi.org/10.3390/pathogens10060671
APA StyleChang, A. -M., & Chen, C. -C. (2021). Molecular Characteristics of Carnivore protoparvovirus 1 with High Sequence Similarity between Wild and Domestic Carnivores in Taiwan. Pathogens, 10(6), 671. https://doi.org/10.3390/pathogens10060671