
Comparison of Chicken Cecal Microbiota after Metaphylactic Treatment or Following Administration of Feed Additives in a Broiler Farm with Enterococcal Spondylitis History

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. General Health and Performance Parameters
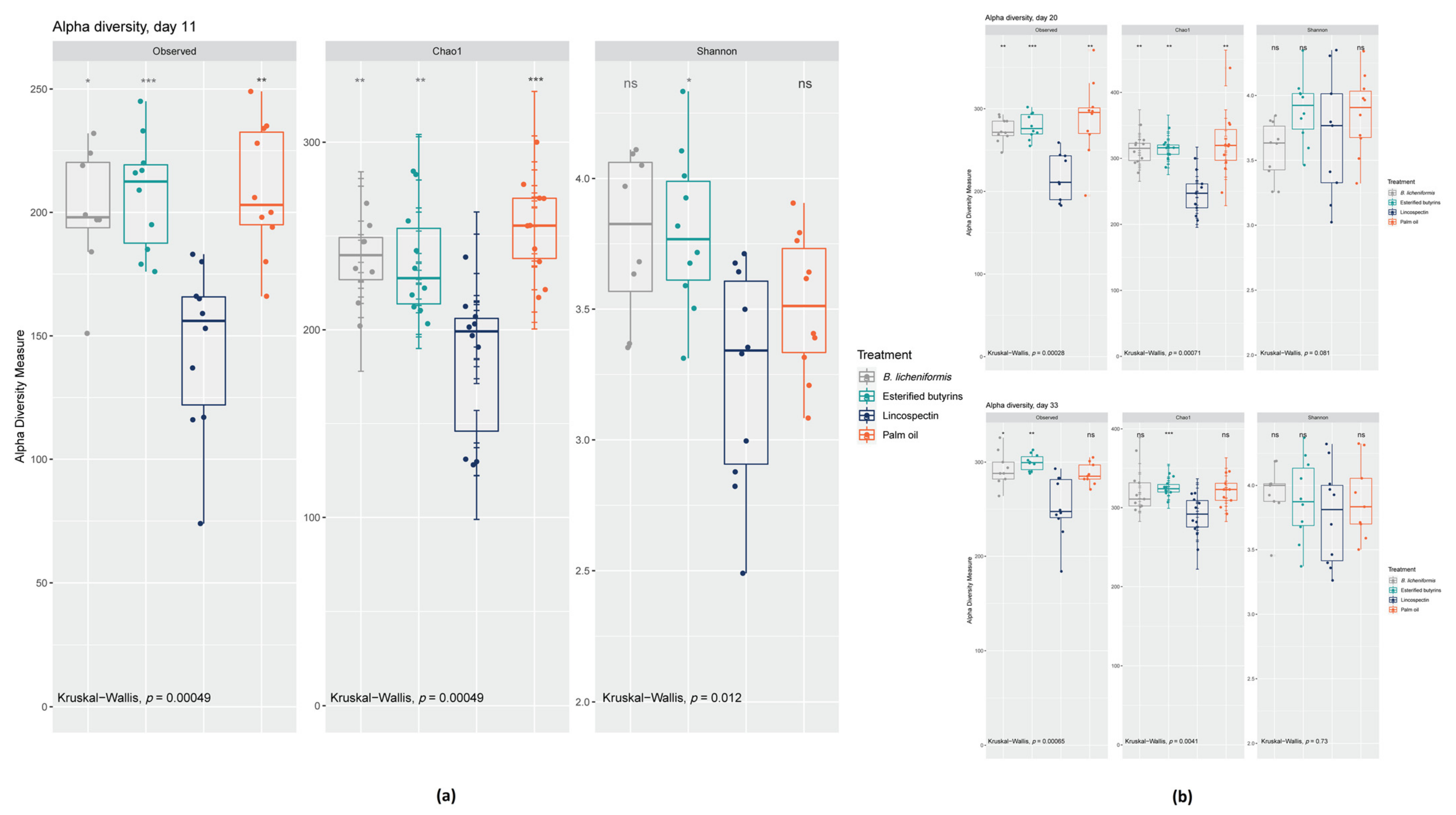
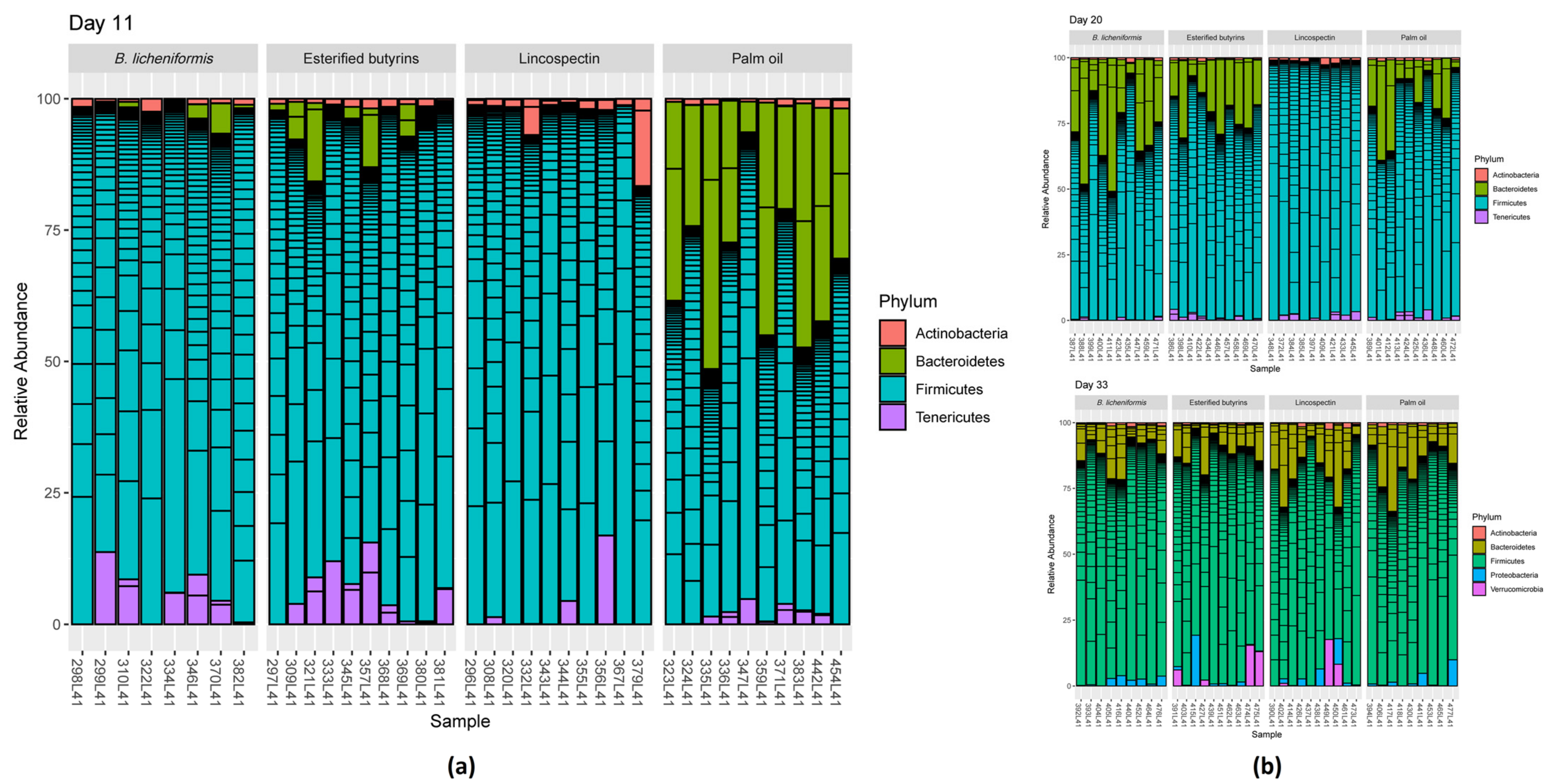
2.2. 16S rRNA Gene Analyses
3. Discussion
3.1. Microbiota Composition under Lincospectin Treatment
3.2. Microbiota Maturation under Administering of Palm Oil
3.3. Similar Microbiota Composition in Chickens Administered B. Licheniformis, Esterified Butyrins and Lincospectin
4. Materials and Methods
4.1. Experimental Design, Animals and Housing, Sampling
4.2. 16S rRNA Gene and Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dolka, B.; Chrobak-Chmiel, D.; Czopowicz, M.; Szeleszczuk, P. Characterization of pathogenic Enterococcus cecorum from different poultry groups: Broiler chickens, layers, turkeys, and waterfowl. PLoS ONE 2017, 12, e0185199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herdt, P.d.; Defoort, P.; Steelant, J.V.; Swam, H.; Tanghe, L.; Goethem, S.v.; Vanrobaeys, M. Enterococcus cecorum osteomyelitis and arthritis in broiler chickens. Vlaams Diergeneeskd. Tijdschr. 2009, 78, 44–48. [Google Scholar]
- Stalker, M.J.; Brash, M.L.; Weisz, A.; Ouckama, R.M.; Slavic, D. Arthritis and osteomyelitis associated with Enterococcus cecorum infection in broiler and broiler breeder chickens in Ontario, Canada. J. Vet. Diagn. Investig. 2010, 22, 643–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, A.; Chen, L.R.; Suyemoto, M.M.; Barnes, H.J.; Borst, L.B. A review of Enterococcus cecorum infection in poultry. Avian Dis. 2018, 62, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.T.; Martin, M.P.; Barnes, H.J. Experimental reproduction of enterococcal spondylitis in male broiler breeder chickens. Avian Dis. 2011, 55, 273–278. [Google Scholar] [CrossRef]
- Borst, L.; Suyemoto, M.; Sarsour, A.; Harris, M.; Martin, M.; Strickland, J.; Oviedo, E.; Barnes, H. Pathogenesis of enterococcal spondylitis caused by Enterococcus cecorum in broiler chickens. Vet. Pathol. 2017, 54, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Kense, M.; Landman, W.J. Enterococcus cecorum infections in broiler breeders and their offspring: Molecular epidemiology. Avian Pathol. 2011, 40, 603–612. [Google Scholar] [CrossRef] [Green Version]
- Armour, N.K.; Collett, S.R.; Williams, S.M. Enterococcus cecorum-related arthritis and osteomyelitis in broilers and broiler breeders. Poult. Inf. Prof. 2011, 117, 1–7. [Google Scholar]
- Robbins, K.M.; Suyemoto, M.M.; Lyman, R.L.; Martin, M.P.; Barnes, H.J.; Borst, L.B. An Outbreak and Source Investigation of Enterococcal Spondylitis in Broilers Caused by Enterococcus cecorum. Avian Dis. 2012, 56, 768–773. [Google Scholar] [CrossRef]
- Walker, G.K.; Suyemoto, M.M.; Gall, S.; Chen, L.; Thakur, S.; Borst, L.B. The role of Enterococcus faecalis during co-infection with avian pathogenic Escherichia coli in avian colibacillosis. Avian Pathol. 2020, 49, 589–599. [Google Scholar] [CrossRef]
- Borst, L.; McLamb, K.; Suyemoto, M.; Chen, L.; Levy, M.; Sarsour, A.; Cordova, H.; Barnes, H.; Oviedo-Rondón, E. Coinfection with Eimeria spp. decreases bacteremia and spinal lesions caused by pathogenic Enterococcus cecorum. Anim. Feed Sci. Technol. 2019, 250, 59–68. [Google Scholar] [CrossRef]
- Sharma, P.; Gupta, S.K.; Barrett, J.B.; Hiott, L.M.; Woodley, T.A.; Kariyawasam, S.; Frye, J.G.; Jackson, C.R. Comparison of Antimicrobial Resistance and Pan-Genome of Clinical and Non-Clinical Enterococcus cecorum from Poultry Using Whole-Genome Sequencing. Foods 2020, 9, 686. [Google Scholar] [CrossRef] [PubMed]
- Stępień-Pyśniak, D.; Marek, A.; Banach, T.; Adaszek, Ł.; Pyzik, E.; Wilczyński, J.; Winiarczyk, S. Prevalence and antibiotic resistance of Enterococcus strains isolated from poultry. Acta Vet. Hung. 2016, 64, 148–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, H.K.; Levine, U.Y.; Looft, T.; Bandrick, M.; Casey, T.A. Treatment, promotion, commotion: Antibiotic alternatives in food-producing animals. Trends Microbiol. 2013, 21, 114–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofacre, C.L.; Mathis, G.F.; Lumpkins, B.S.; Sygall, R.; Vaessen, S.; Hofacre, C.S.; Smith, J.A.; Clanton, E. Efficacy of Butyric and Valeric Acid Esters in a Necrotic Enteritis Challenge Model. Avian Dis. 2020, 64, 407–414. [Google Scholar] [CrossRef]
- Timbermont, L.; Lanckriet, A.; Dewulf, J.; Nollet, N.; Schwarzer, K.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Control of Clostridium perfringens-induced necrotic enteritis in broilers by target-released butyric acid, fatty acids and essential oils. Avian Pathol. 2010, 39, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Clausen, M.R.; Mortensen, P.B. Kinetic studies on the metabolism of short-chain fatty acids and glucose by isolated rat colonocytes. Gastroenterology 1994, 106, 423–432. [Google Scholar] [CrossRef]
- McIntyre, A.; Gibson, P.; Young, G. Butyrate production from dietary fibre and protection against large bowel cancer in a rat model. Gut 1993, 34, 386–391. [Google Scholar] [CrossRef] [Green Version]
- Dalmasso, G.; Nguyen, H.T.T.; Yan, Y.; Charrier-Hisamuddin, L.; Sitaraman, S.V.; Merlin, D. Butyrate transcriptionally enhances peptide transporter PepT1 expression and activity. PLoS ONE 2008, 3, e2476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inan, M.S.; Rasoulpour, R.J.; Yin, L.; Hubbard, A.K.; Rosenberg, D.W.; Giardina, C. The luminal short-chain fatty acid butyrate modulates NF-κB activity in a human colonic epithelial cell line. Gastroenterology 2000, 118, 724–734. [Google Scholar] [CrossRef]
- Peng, L.; He, Z.; Chen, W.; Holzman, I.R.; Lin, J. Effects of Butyrate on Intestinal Barrier Function in a Caco-2 Cell Monolayer Model of Intestinal Barrier. Pediatr. Res. 2007, 61, 37. [Google Scholar] [CrossRef] [Green Version]
- Mariadason, J.M.; Barkla, D.H.; Gibson, P.R. Effect of short-chain fatty acids on paracellular permeability in Caco-2 intestinal epithelium model. Am. J. Physiol. Gastrointest. Liver Physiol. 1997, 272, G705–G712. [Google Scholar] [CrossRef]
- Hankel, J.; Jung, K.; Kuder, H.; Keller, B.; Keller, C.; Galvez, E.; Strowig, T.; Visscher, C. Caecal Microbiota of Experimentally Camyplobacter jejuni-Infected Chickens at Different Ages. Front. Microbiol. 2019, 10, 2303. [Google Scholar] [CrossRef]
- Onrust, L.; Baeyen, S.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Effect of in feed administration of different butyrate formulations on Salmonella Enteritidis colonization and cecal microbiota in broilers. Vet. Res. 2020, 51, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Fernández, S.M.; Cretenet, M.; Bernardeau, M. In vitro inhibition of avian pathogenic Enterococcus cecorum isolates by probiotic Bacillus strains. Poult. Sci. 2019, 98, 2338–2346. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Lin, Y.; Zeng, D.; Zhou, M.; Zeng, Y.; Wang, H.; Zhou, Y.; Zhu, H.; Pan, K.; Jing, B. Bacillus licheniformis normalize the ileum microbiota of chickens infected with necrotic enteritis. Sci. Rep. 2018, 8, 1744. [Google Scholar] [CrossRef] [PubMed]
- Hovorková, P.; Laloučková, K.; Skřivanová, E. Determination of In Vitro Antibacterial Activity of Plant Oils Containing Medium-Chain Fatty Acids against Gram-Positive Pathogenic and Gut Commensal Bacteria. Czech. J. Anim. Sci. 2018, 63, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Aviagen. Ross 308 Broiler: Performance Objectives. Available online: https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&ved=2ahUKEwi-1dHHjpzwAhVJgf0HHZzyD3QQFjAAegQIBhAD&url=https%3A%2F%2Fwww.winmixsoft.com%2Ffiles%2Finfo%2FRoss-308-Broiler-PO-2014-EN.pdf&usg=AOvVaw2FfKFdmg8PopdgG2t-Dl51 (accessed on 26 April 2021).
- Ward, T.L.; Weber, B.P.; Mendoza, K.M.; Danzeisen, J.L.; Llop, K.; Lang, K.; Clayton, J.B.; Grace, E.; Brannon, J.; Radovic, I. Antibiotics and Host-Tailored Probiotics Similarly Modulate Effects on the Developing Avian Microbiome, Mycobiome, and Host Gene Expression. MBio 2019, 10, e02171-19. [Google Scholar] [CrossRef] [Green Version]
- Zwirzitz, B.; Pinior, B.; Metzler-Zebeli, B.; Handler, M.; Gense, K.; Knecht, C.; Ladinig, A.; Dzieciol, M.; Wetzels, S.U.; Wagner, M. Microbiota of the Gut-Lymph Node Axis: Depletion of Mucosa-Associated Segmented Filamentous Bacteria and Enrichment of Methanobrevibacter by Colistin Sulfate and Linco-Spectin in Pigs. Front. Microbiol. 2019, 10, 599. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, V.E.; Battaglia, T.; Kurtz, Z.D.; Bijnens, L.; Ou, A.; Engstrand, I.; Zheng, X.; Iizumi, T.; Mullins, B.J.; Müller, C.L. A single early-in-life macrolide course has lasting effects on murine microbial network topology and immunity. Nat. Commun. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Tonks, A.A. Exploring the Effects of Management Strategies on the Gut Microbiome and Metabolome of Growing Broiler Chickens: An Integrated Metagenomic and Metabolomic Approach. Ph.D. Thesis, University of Reading, Reading, UK, 2018. [Google Scholar]
- Pereira, R.; Bortoluzzi, C.; Durrer, A.; Fagundes, N.S.; Pedroso, A.A.; Rafael, J.M.; Perim, J.E.d.L.; Zavarize, K.C.; Napty, G.S.; Andreote, F.D. Performance and intestinal microbiota of chickens receiving probiotic in the feed and submitted to antibiotic therapy. J. Anim. Physiol. Anim. Nutr. 2019, 103, 72–86. [Google Scholar] [CrossRef]
- Fjalstad, J.W.; Esaiassen, E.; Juvet, L.K.; van den Anker, J.N.; Klingenberg, C. Antibiotic therapy in neonates and impact on gut microbiota and antibiotic resistance development: A systematic review. J. Antimicrob. Chemother. 2018, 73, 569–580. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, P.; Wigley, P.; Fothergill, J.; Bernardeau, M. Development of the Caecal Microbiota in Three Broiler Breeds. Front. Vet. Sci. 2019, 6, 201. [Google Scholar] [CrossRef] [PubMed]
- Oakley, B.B.; Buhr, R.J.; Ritz, C.W.; Kiepper, B.H.; Berrang, M.E.; Seal, B.S.; Cox, N.A. Successional changes in the chicken cecal microbiome during 42 days of growth are independent of organic acid feed additives. BMC Vet. Res. 2014, 10, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubasova, T.; Kollarcikova, M.; Crhanova, M.; Karasova, D.; Cejkova, D.; Sebkova, A.; Matiasovicova, J.; Faldynova, M.; Pokorna, A.; Cizek, A. Contact with adult hen affects development of caecal microbiota in newly hatched chicks. PLoS ONE 2019, 14, e0212446. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Li, L.; Willer, T.; Baumgärtner, W.; Rautenschlein, S. Adhesion and invasion of Campylobacter jejuni in chickens with a modified gut microbiota due to antibiotic treatment. Vet. Microbiol. 2020, 240, 108504. [Google Scholar] [CrossRef]
- Jacobs-Reitsma, W.; Van de Giessen, A.; Bolder, N.; Mulder, R. Epidemiology of Campylobacter spp. at two Dutch broiler farms. Epidemiol. Infect. 1995, 114, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Stern, N.J.; Cox, N.A.; Musgrove, M.T.; Park, C. Incidence and Levels of Campylobacter in Broilers After Exposure to an Inoculated Seeder Bird. J. Appl. Poult. Res. 2001, 10, 315–318. [Google Scholar] [CrossRef]
- Rychlik, I. Composition and Function of Chicken Gut Microbiota. Animals 2020, 10, 103. [Google Scholar] [CrossRef] [Green Version]
- Ocejo, M.; Oporto, B.; Hurtado, A. 16S rRNA amplicon sequencing characterization of caecal microbiome composition of broilers and free-range slow-growing chickens throughout their productive lifespan. Sci. Rep. 2019, 9, 2506. [Google Scholar] [CrossRef] [Green Version]
- Videnska, P.; Sedlar, K.; Lukac, M.; Faldynova, M.; Gerzova, L.; Cejkova, D.; Sisak, F.; Rychlik, I. Succession and replacement of bacterial populations in the caecum of egg laying hens over their whole life. PLoS ONE 2014, 9, e115142. [Google Scholar] [CrossRef] [Green Version]
- Stackebrandt, E. The family Lachnospiraceae. In The Prokaryotes, 4th ed.; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 197–201. [Google Scholar]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef]
- Biddle, A.; Stewart, L.; Blanchard, J.; Leschine, S. Untangling the Genetic Basis of Fibrolytic Specialization by Lachnospiraceae and Ruminococcaceae in Diverse Gut Communities. Diversity 2013, 5, 627–640. [Google Scholar] [CrossRef]
- Guilloteau, P.; Martin, L.; Eeckhaut, V.; Ducatelle, R.; Zabielski, R.; Van Immerseel, F. From the gut to the peripheral tissues: The multiple effects of butyrate. Nutr. Res. Rev. 2010, 23, 366–384. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhong, R.; Zhang, L.; Zhang, H. The Chronic Effect of Transgenic Maize Line with mCry1Ac or maroACC gene on Ileal Microbiota Using a Hen Model. Microorganisms 2019, 7, 92. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Wang, L.; Khan, A.; Zhao, R.; Wei, S.; Jing, X. Fermented wheat bran by xylanase-producing Bacillus cereus boosts the intestinal microflora of broiler chickens. Poult. Sci. 2020, 99, 263–271. [Google Scholar] [CrossRef]
- Patterson, J.; Burkholder, K. Application of prebiotics and probiotics in poultry production. Poult. Sci. 2003, 82, 627–631. [Google Scholar] [CrossRef] [PubMed]
- Abd El, A.E.-M.E.; El-Wardany, I.; Abu-Taleb, A.M.; Wakwak, M.M.; Ebeid, T.A.; Saleh, A.A. Assessment of in ovo administration of Bifidobacterium bifidum and Bifidobacterium longum on performance, ileal histomorphometry, blood hematological, and biochemical parameters of broilers. Probiotics. Antimicro. Prot. 2020, 12, 439–450. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Yu, Y.-H. Bacillus licheniformis–fermented products improve growth performance and the fecal microbiota community in broilers. Poult. Sci. 2020, 99, 1432–1443. [Google Scholar] [CrossRef] [PubMed]
- Trela, J.; Kierończyk, B.; Hautekiet, V.; Józefiak, D. Combination of Bacillus licheniformis and salinomycin: Effect on the growth performance and git microbial populations of broiler chickens. Animals 2020, 10, 889. [Google Scholar] [CrossRef] [PubMed]
- Zaghari, M.; Sarani, P.; Hajati, H. Comparison of two probiotic preparations on growth performance, intestinal microbiota, nutrient digestibility and cytokine gene expression in broiler chickens. J. Appl. Anim. Res. 2020, 48, 166–175. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Nat. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P. Vegan: Community Ecology Package, R Package Version 2.5-6; 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 8 August 2021).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Day of Life | Chicken House 1 | Chicken House 2 | Chicken House 3 | Chicken House 4 |
|---|---|---|---|---|
| 28 | 1538 (n = 7285) | 1559 (n = 7113) | 1457 (n = 7025) | |
| 33 | 1977 (n = 4958) | 1984 (n = 4954) | 1920 (n = 5006) | 1902 (n = 8255) |
| 37 | 2350 (n = 25,025) | 2424 (n = 23,854) | ||
| 38 | 2444 (n = 24,677) | 2467 (n = 28,224) |
| Day 11 | Day 20 | Day 33 | ||||
|---|---|---|---|---|---|---|
| R2 | Pr(>F) | R2 | Pr(>F) | R2 | Pr(>F) | |
| Treatment | 0.2520 | 0.001 | 0.2430 | 0.001 | 0.1667 | 0.001 |
| B. licheniformis | Esterified Butyrins | Palm Oil | ||
|---|---|---|---|---|
| Lincospectin | Day 11 | 0.4110 *** | 0.4778 *** | 0.7518 *** |
| Day 20 | 0.7355 *** | 0.6055 *** | 0.5668 *** | |
| Day 33 | 0.1854 *** | 0.1770 *** | 0.3005 *** |
| Day 11 | ||||
|---|---|---|---|---|
| Family | B. licheniformis | Esterified Butyrins | Lincospectin | Palm Oil |
| Bacteroidaceae | 0.955 ± 1.41 | 3.15 ± 3.96 | 0.010 ± 0.007 | 24.7 ± 11.1 |
| Ruminococcaceae | 57.0 ± 13.5 | 57.5 ± 6.10 | 41.1 ± 10.7 | 45.9 ± 9.60 |
| Lachnospiraceae | 17.8 ± 14.7 | 14.2 ± 4.77 | 29.5 ± 10.8 | 12.5 ± 4.22 |
| Erysipelotrichaceae | 3.38 ± 1.89 | 4.64 ± 3.60 | 13.2 ± 13.0 | 3.29 ± 1.77 |
| Bifidobacteriaceae | 0.005 ± 0.011 | 0.005 ± 0.009 | 1.94 ± 3.54 | 0.026 ± 0.063 |
| Staphylococcaceae | 0 | 0.002 ± 0.004 | 0.179 ± 0.209 | 0.011 ± 0.017 |
| Enterococcaceae | 0.031 ± 0.035 | 0.158 ± 0.157 | 0.207 ± 0.172 | 0.110 ± 0.231 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hankel, J.; Bodmann, B.; Todte, M.; Galvez, E.; Strowig, T.; Radko, D.; Antakli, A.; Visscher, C. Comparison of Chicken Cecal Microbiota after Metaphylactic Treatment or Following Administration of Feed Additives in a Broiler Farm with Enterococcal Spondylitis History. Pathogens 2021, 10, 1068. https://doi.org/10.3390/pathogens10081068
Hankel J, Bodmann B, Todte M, Galvez E, Strowig T, Radko D, Antakli A, Visscher C. Comparison of Chicken Cecal Microbiota after Metaphylactic Treatment or Following Administration of Feed Additives in a Broiler Farm with Enterococcal Spondylitis History. Pathogens. 2021; 10(8):1068. https://doi.org/10.3390/pathogens10081068
Chicago/Turabian StyleHankel, Julia, Björn Bodmann, Matthias Todte, Eric Galvez, Till Strowig, Dimitri Radko, Ali Antakli, and Christian Visscher. 2021. "Comparison of Chicken Cecal Microbiota after Metaphylactic Treatment or Following Administration of Feed Additives in a Broiler Farm with Enterococcal Spondylitis History" Pathogens 10, no. 8: 1068. https://doi.org/10.3390/pathogens10081068
APA StyleHankel, J., Bodmann, B., Todte, M., Galvez, E., Strowig, T., Radko, D., Antakli, A., & Visscher, C. (2021). Comparison of Chicken Cecal Microbiota after Metaphylactic Treatment or Following Administration of Feed Additives in a Broiler Farm with Enterococcal Spondylitis History. Pathogens, 10(8), 1068. https://doi.org/10.3390/pathogens10081068