
Genomic Variability of Canine Parvoviruses from a Selected Population of Dogs and Cats in Sri Lanka

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Sampled Animals
2.2. CPV-2 PCR Positive Samples
2.3. CPV-2 Subtyping
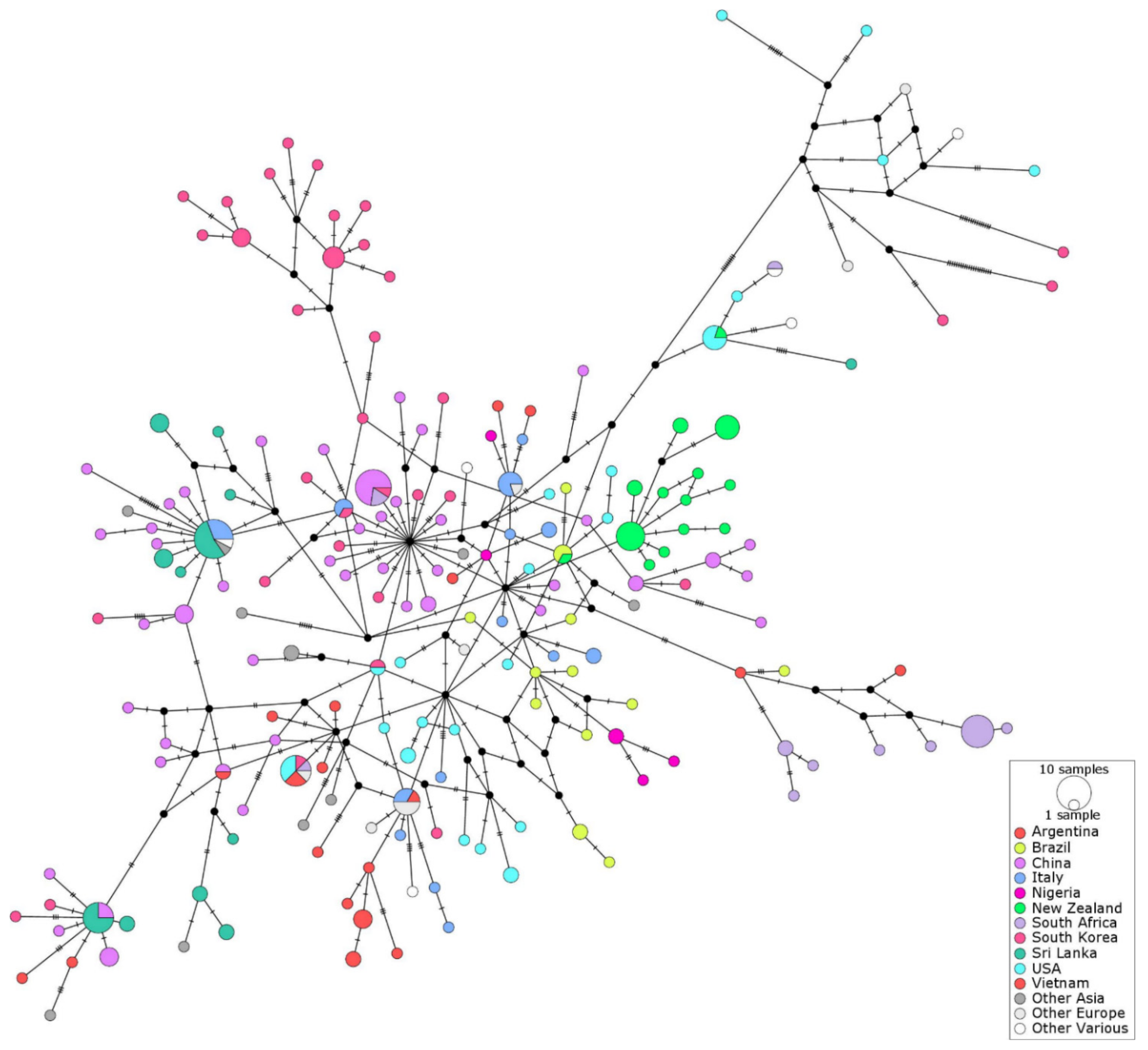
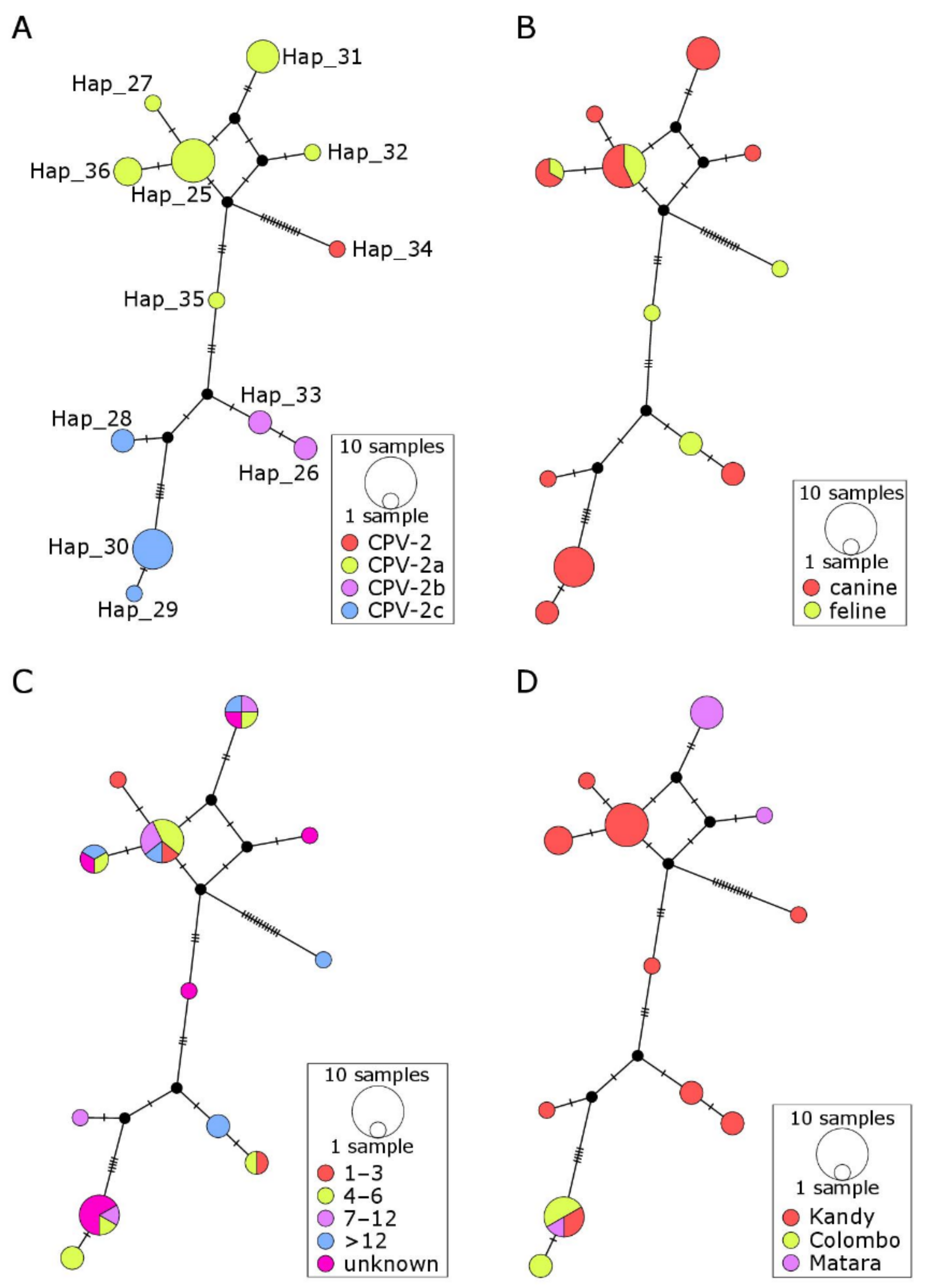
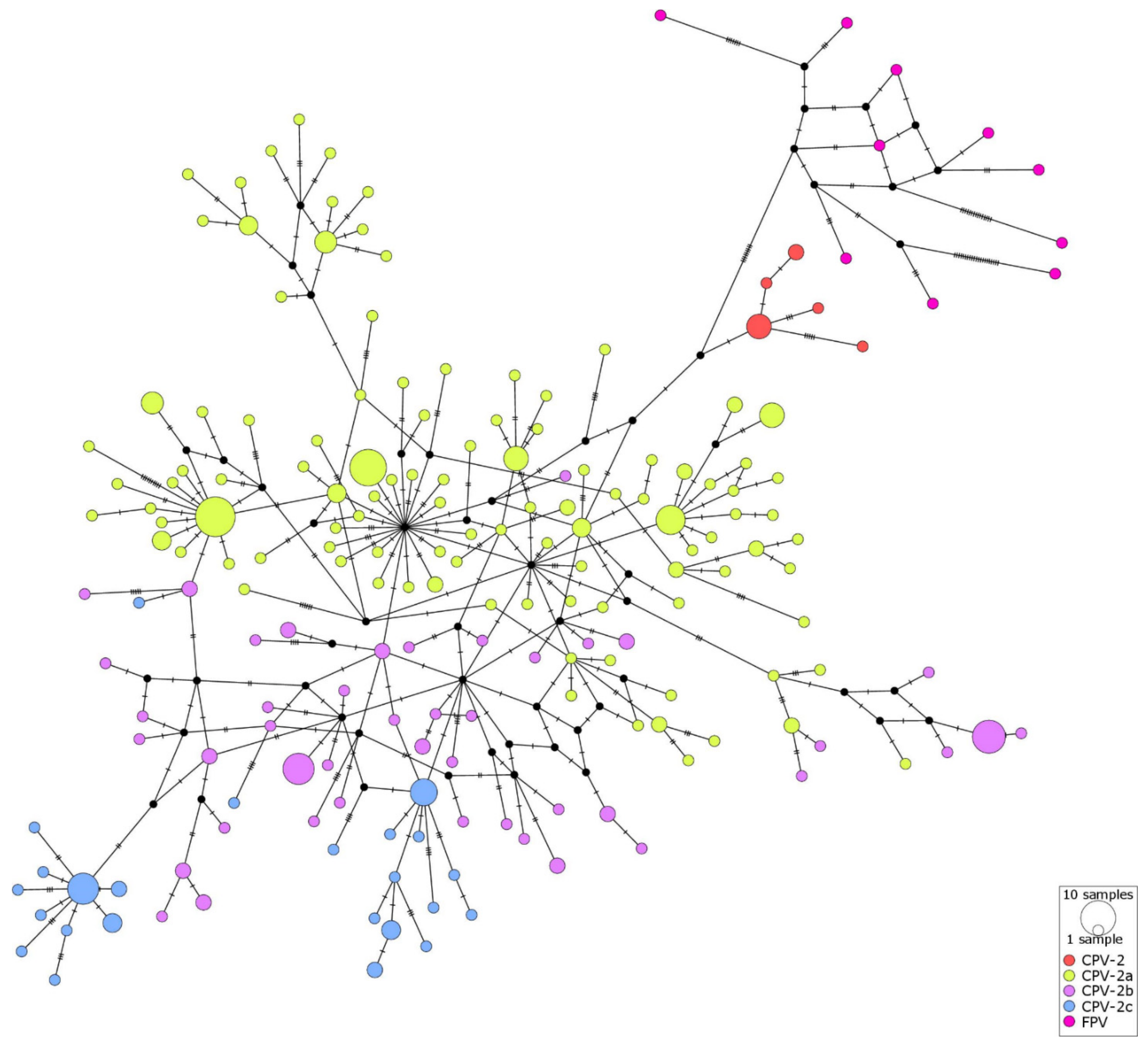
2.4. Haplotype Networks
3. Discussion
4. Materials and Methods
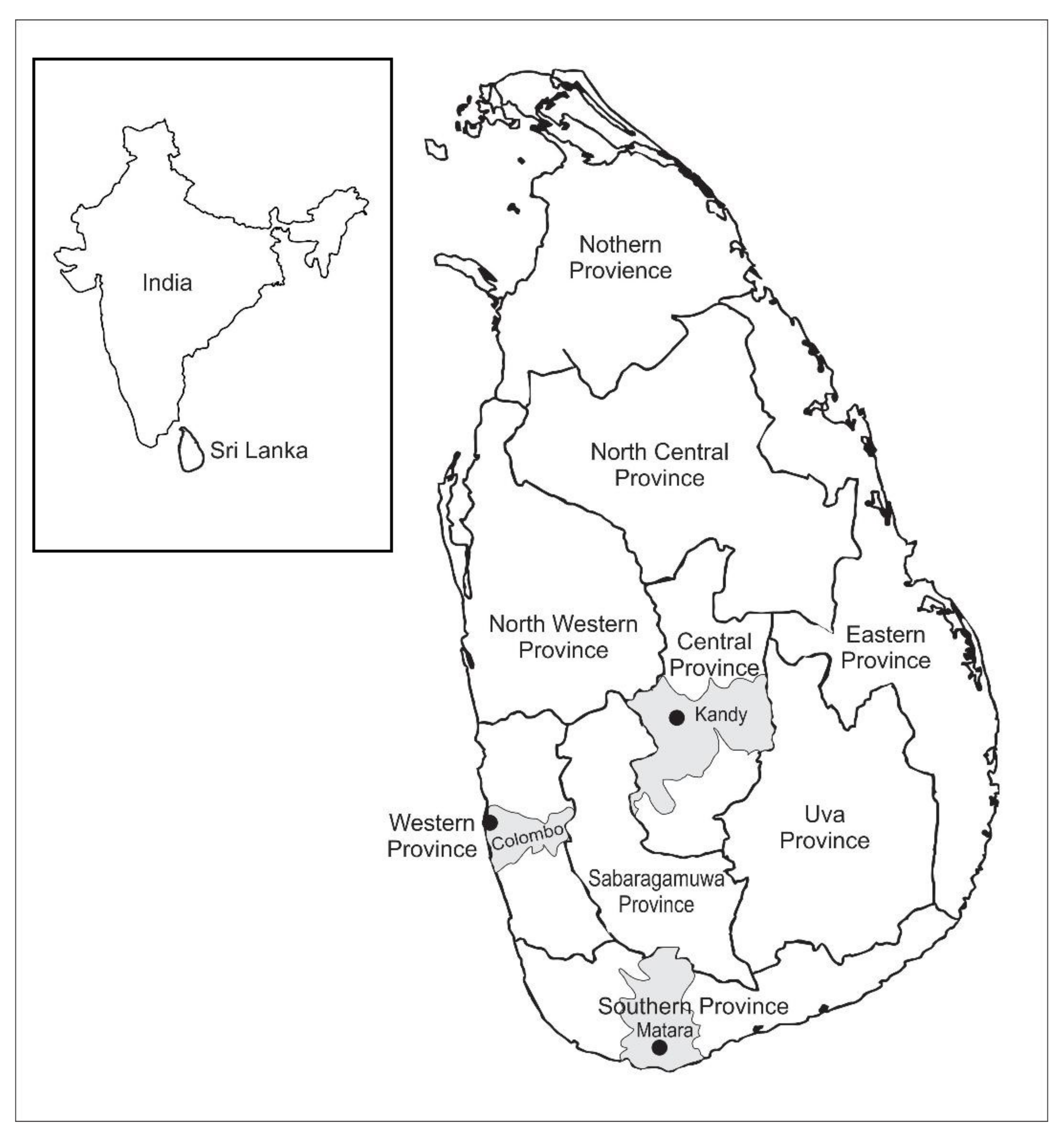
4.1. Sources of Samples
4.2. Processing of Samples
4.3. CPV-2 PCR
4.4. CPV Subtyping
4.5. Molecular Network Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cotmore, S.F.; Agbandje-McKenna, M.; Canuti, M.; Chiorini, J.A.; Eis-Hubinger, A.M.; Hughes, J.; Mietzsch, M.; Modha, S.; Ogliastro, M.; Penzes, J.J.; et al. ICTV Virus Taxonomy Profile: Parvoviridae. J. Gen. Virol. 2019, 100, 367–368. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.C.; Angrick, E.J. Canine parvovirus: Environmental effects on infectivity. Am. J. Vet. Res. 1986, 47, 1464–1467. [Google Scholar] [PubMed]
- Carmichael, L.E. An annotated historical account of canine parvovirus. J. Vet. Med. B Infect. Dis. Vet. Public Health 2005, 52, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Parrish, C.R.; Holmes, E.C.; Morens, D.M.; Park, E.C.; Burke, D.S.; Calisher, C.H.; Laughlin, C.A.; Saif, L.J.; Daszak, P. Cross-species virus transmission and the emergence of new epidemic diseases. Microbiol. Mol. Biol. Rev. 2008, 72, 457–470. [Google Scholar] [CrossRef] [Green Version]
- Wilson, N.D. Origin of canine parvovirus. Vet. Rec. 1980, 106, 392. [Google Scholar] [CrossRef] [PubMed]
- Hoelzer, K.; Parrish, C.R. The emergence of parvoviruses of carnivores. Vet. Res. 2010, 41, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, R.D.; Thiel, B.; Mukhtar, E.; Sharp, P.; Larson, L.J. Age and long-term protective immunity in dogs and cats. J. Comp. Pathol. 2010, 142 (Suppl. 1), S102–S108. [Google Scholar] [CrossRef]
- Decaro, N.; Buonavoglia, C. Canine parvovirus–a review of epidemiological and diagnostic aspects, with emphasis on type 2c. Vet. Microbiol. 2012, 155, 1–12. [Google Scholar] [CrossRef]
- Gamage, B.; Dissanayake, D.R.A.; Prasada, D.V.P.; Silva, I.D. Risk, prognosis and causality of parvo viral enteritis in dogs in Sri Lanka. Comp. Immunol. Microbiol. Infect. Dis. 2020, 72, 101496. [Google Scholar] [CrossRef]
- Reed, A.P.; Jones, E.V.; Miller, T.J. Nucleotide sequence and genome organization of canine parvovirus. J. Virol. 1988, 62, 266–276. [Google Scholar] [CrossRef] [Green Version]
- Parrish, C.R.; Aquadro, C.F.; Strassheim, M.L.; Evermann, J.F.; Sgro, J.Y.; Mohammed, H.O. Rapid antigenic-type replacement and DNA sequence evolution of canine parvovirus. J. Virol. 1991, 65, 6544–6552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrish, C.R. Host range relationships and the evolution of canine parvovirus. Vet. Microbiol. 1999, 69, 29–40. [Google Scholar] [CrossRef]
- Hueffer, K.; Parrish, C.R. Parvovirus host range, cell tropism and evolution. Curr. Opin. Microbiol. 2003, 6, 392–398. [Google Scholar] [CrossRef]
- Buonavoglia, C.; Martella, V.; Pratelli, A.; Tempesta, M.; Cavalli, A.; Buonavoglia, D.; Bozzo, G.; Elia, G.; Decaro, N.; Carmichael, L. Evidence for evolution of canine parvovirus type 2 in Italy. J. Gen. Virol. 2001, 82, 3021–3025. [Google Scholar] [CrossRef]
- Nakamura, M.; Tohya, Y.; Miyazawa, T.; Mochizuki, M.; Phung, H.T.; Nguyen, N.H.; Huynh, L.M.; Nguyen, L.T.; Nguyen, P.N.; Nguyen, P.V.; et al. A novel antigenic variant of Canine parvovirus from a Vietnamese dog. Arch. Virol. 2004, 149, 2261–2269. [Google Scholar] [CrossRef]
- Hong, C.; Decaro, N.; Desario, C.; Tanner, P.; Pardo, M.C.; Sanchez, S.; Buonavoglia, C.; Saliki, J.T. Occurrence of canine parvovirus type 2c in the United States. J. Vet. Diagn. Investig. 2007, 19, 535–539. [Google Scholar] [CrossRef] [Green Version]
- Perez, R.; Francia, L.; Romero, V.; Maya, L.; Lopez, I.; Hernandez, M. First detection of canine parvovirus type 2c in South America. Vet. Microbiol. 2007, 124, 147–152. [Google Scholar] [CrossRef]
- Touihri, L.; Bouzid, I.; Daoud, R.; Desario, C.; El Goulli, A.F.; Decaro, N.; Ghorbel, A.; Buonavoglia, C.; Bahloul, C. Molecular characterization of canine parvovirus-2 variants circulating in Tunisia. Virus Genes 2009, 38, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Desario, C.; Addie, D.D.; Martella, V.; Vieira, M.J.; Elia, G.; Zicola, A.; Davis, C.; Thompson, G.; Thiry, E.; et al. The study molecular epidemiology of canine parvovirus, Europe. Emerg. Infect. Dis. 2007, 13, 1222–1224. [Google Scholar] [CrossRef]
- Miranda, C.; Carvalheira, J.; Parrish, C.R.; Thompson, G. Factors affecting the occurrence of canine parvovirus in dogs. Vet. Microbiol. 2015, 180, 59–64. [Google Scholar] [CrossRef]
- Miranda, C.; Thompson, G. Canine parvovirus: The worldwide occurrence of antigenic variants. J. Gen. Virol. 2016, 97, 2043–2057. [Google Scholar] [CrossRef] [PubMed]
- Import/Export–Procedure for Import/Export. Available online: http://www.daph.gov.lk/web/index.php?option=com_content&view=article&id=24&Itemid=126&lang=en (accessed on 20 June 2021).
- Ubeyratne, K.; Srikitjakarn, L.; Pfeiffer, D.; Kong, F.; Sunil-Chandra, N.; Chaisowwong, W.; Hemwan, P. A Knowledge, Attitudes and Practices (KAP) survey on canine rabies prevention and control in four rural areas of Sri Lanka. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Sanchez-Soriano, C.; Gibson, A.D.; Gamble, L.; Burdon Bailey, J.L.; Green, S.; Green, M.; Bronsvoort, B.M.D.; Handel, I.G.; Mellanby, R.J.; Mazeri, S. Development of a high number, high coverage dog rabies vaccination programme in Sri Lanka. BMC Infect. Dis. 2019, 19, 977. [Google Scholar] [CrossRef] [Green Version]
- Yapa, A.; Ratnavira, G. The Mammals of Sri Lanka; Field Ornithology Group Sri Lanka: Colombo, Sri Lanka, 2013. [Google Scholar]
- Veterinary Drug Control Authority of Sri Lanka. Registered Veterinary Medical Products for Import and Free Sale. 2021. Available online: http://www.daph.gov.lk/web/images/content_image/vra/2021/VDCAregisteredVMPs_for_import_and_free_sale_updated_to_31.12.2020.pdf (accessed on 9 July 2021).
- Meers, J.; Kyaw-Tanner, M.; Bensink, Z.; Zwijnenberg, R. Genetic analysis of canine parvovirus from dogs in Australia. Aust. Vet. J. 2007, 85, 392–396. [Google Scholar] [CrossRef]
- Ohneiser, S.A.; Hills, S.F.; Cave, N.J.; Passmore, D.; Dunowska, M. Canine parvoviruses in New Zealand form a monophyletic group distinct from the viruses circulating in other parts of the world. Vet. Microbiol. 2015, 178, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Houston, D.M.; Ribble, C.S.; Head, L.L. Risk factors associated with parvovirus enteritis in dogs: 283 cases (1982–1991). J. Am. Vet. Med. Assoc. 1996, 208, 542–546. [Google Scholar]
- Giraldo-Ramirez, S.; Rendon-Marin, S.; Ruiz-Saenz, J. Phylogenetic, Evolutionary and Structural Analysis of Canine Parvovirus (CPV-2) Antigenic Variants Circulating in Colombia. Viruses 2020, 12, 500. [Google Scholar] [CrossRef]
- Allison, A.B.; Harbison, C.E.; Pagan, I.; Stucker, K.M.; Kaelber, J.T.; Brown, J.D.; Ruder, M.G.; Keel, M.K.; Dubovi, E.J.; Holmes, E.C.; et al. Role of multiple hosts in the cross-species transmission and emergence of a pandemic parvovirus. J. Virol. 2012, 86, 865–872. [Google Scholar] [CrossRef] [Green Version]
- Steinel, A.; Parrish, C.R.; Bloom, M.E.; Truyen, U. Parvovirus infections in wild carnivores. J. Wildl. Dis. 2001, 37, 594–607. [Google Scholar] [CrossRef]
- Truyen, U.; Evermann, J.F.; Vieler, E.; Parrish, C.R. Evolution of canine parvovirus involved loss and gain of feline host range. Virology 1996, 215, 186–189. [Google Scholar] [CrossRef] [Green Version]
- Martella, V.; Decaro, N.; Elia, G.; Buonavoglia, C. Surveillance activity for canine parvovirus in Italy. J. Vet. Med. B Infect. Dis. Vet. Public Health 2005, 52, 312–315. [Google Scholar] [CrossRef]
- Castro, T.X.; Costa, E.M.; Leite, J.P.; Labarthe, N.V.; Cubel Garcia, R.C. Monitoring of canine parvovirus (CPV) strains detected in vaccinated puppies in Brazil. Res. Vet. Sci. 2011, 90, 336–340. [Google Scholar] [CrossRef]
- Inthong, N.; Kaewmongkol, S.; Meekhanon, N.; Sirinarumitr, K.; Sirinarumitr, T. Dynamic evolution of canine parvovirus in Thailand. Vet. World 2020, 13, 245–255. [Google Scholar] [CrossRef] [Green Version]
- Clark, N.J.; Seddon, J.M.; Kyaw-Tanner, M.; Al-Alawneh, J.; Harper, G.; McDonagh, P.; Meers, J. Emergence of canine parvovirus subtype 2b (CPV-2b) infections in Australian dogs. Infect. Genet. Evol. 2018, 58, 50–55. [Google Scholar] [CrossRef]
- Woolford, L.; Crocker, P.; Bobrowski, H.; Baker, T.; Hemmatzadeh, F. Detection of the Canine Parvovirus 2c Subtype in Australian Dogs. Viral. Immunol. 2017, 30, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Franzo, G.; Tucciarone, C.M.; Casagrande, S.; Caldin, M.; Cortey, M.; Furlanello, T.; Legnardi, M.; Cecchinato, M.; Drigo, M. Canine parvovirus (CPV) phylogeny is associated with disease severity. Sci. Rep. 2019, 9, 11266. [Google Scholar] [CrossRef]
- Moon, H.-S.; Lee, S.-A.; Lee, S.-G.; Choi, S.-Y.; Kim, D.; Hyun, C. Comparison of the pathogenicity in three different Korean canine parvovirus 2 (CPV-2) isolates. Vet. Microbiol. 2008, 131, 47–56. [Google Scholar] [CrossRef]
- Kalli, I.; Leontides, L.S.; Mylonakis, M.E.; Adamama-Moraitou, K.; Rallis, T.; Koutinas, A.F. Factors affecting the occurrence, duration of hospitalization and final outcome in canine parvovirus infection. Res. Vet. Sci. 2010, 89, 174–178. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Zakrajsek, B.A. Effect of experimental treatment on housekeeping gene expression: Validation by real-time, quantitative RT-PCR. J. Biochem. Biophys. Methods 2000, 46, 69–81. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.; Bryant, D. Popart: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test | Variation within Populations | Variation between Populations | PhiST | p |
|---|---|---|---|---|
| International network | ||||
| Geographical region 1 | 77.1% | 22.9% | 0.23 | <0.001 |
| Decade of collection 2 | 83.8% | 16.2% | 0.16 | <0.001 |
| Subtype 3 | 51.7% | 48.3% | 0.48 | <0.001 |
| Sri Lankan network | ||||
| Location 4 | 76.5% | 23.5% | 0.23 | 0.001 |
| Age-group 5 | 106.2% | −6.2% | −0.06 | 0.879 |
| Host 6 | 95.6% | 4.4% | 0.04 | 0.080 |
| Subtypes 7 | 16.6% | 83.4% | 0.83 | <0.001 |
| Subtype | Nucleotide (Amino Acid) Position 1 | |||||
|---|---|---|---|---|---|---|
| 3045 (87) | 3087 (101) | 3684 (300) | 3699 (305) | 4062 (426) | 4449 (555) | |
| CPV-2 | ATG (Met) | ATT (Ile) | GCT (Ala) | GAT (Asp) | AAT (Asn) | GTA (Val) |
| CPV2a | TTG (Leu) | ACT (Thr) | GGT (Gly) | TAT (Tyr) | AAT (Asn) | ATA (Ile) |
| CPV2b | TTG (Leu) | ACT (Thr) | GGT (Gly) | TAT (Tyr) | GAT (Asp) | ATA (Ile) |
| CPV2c | TTG (Leu) | ACT (Thr) | GGT (Gly) | TAT (Tyr) | GAA (Glu) | ATA (Ile) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jinadasa, R.; Ghosh, S.; Hills, S.; Premalal, T.; Atapattu, U.; Fuward, M.; Kalupahana, W.; Dunowska, M. Genomic Variability of Canine Parvoviruses from a Selected Population of Dogs and Cats in Sri Lanka. Pathogens 2021, 10, 1102. https://doi.org/10.3390/pathogens10091102
Jinadasa R, Ghosh S, Hills S, Premalal T, Atapattu U, Fuward M, Kalupahana W, Dunowska M. Genomic Variability of Canine Parvoviruses from a Selected Population of Dogs and Cats in Sri Lanka. Pathogens. 2021; 10(9):1102. https://doi.org/10.3390/pathogens10091102
Chicago/Turabian StyleJinadasa, Rasika, Sayani Ghosh, Simon Hills, Thushini Premalal, Ushani Atapattu, Manohari Fuward, Wasantha Kalupahana, and Magdalena Dunowska. 2021. "Genomic Variability of Canine Parvoviruses from a Selected Population of Dogs and Cats in Sri Lanka" Pathogens 10, no. 9: 1102. https://doi.org/10.3390/pathogens10091102
APA StyleJinadasa, R., Ghosh, S., Hills, S., Premalal, T., Atapattu, U., Fuward, M., Kalupahana, W., & Dunowska, M. (2021). Genomic Variability of Canine Parvoviruses from a Selected Population of Dogs and Cats in Sri Lanka. Pathogens, 10(9), 1102. https://doi.org/10.3390/pathogens10091102