
HERV-W and Mycobacterium avium subspecies paratuberculosis Are at Play in Pediatric Patients at Onset of Type 1 Diabetes

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Blood Samples
2.3. Peptides
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
2.5. Statistical Analysis
3. Results
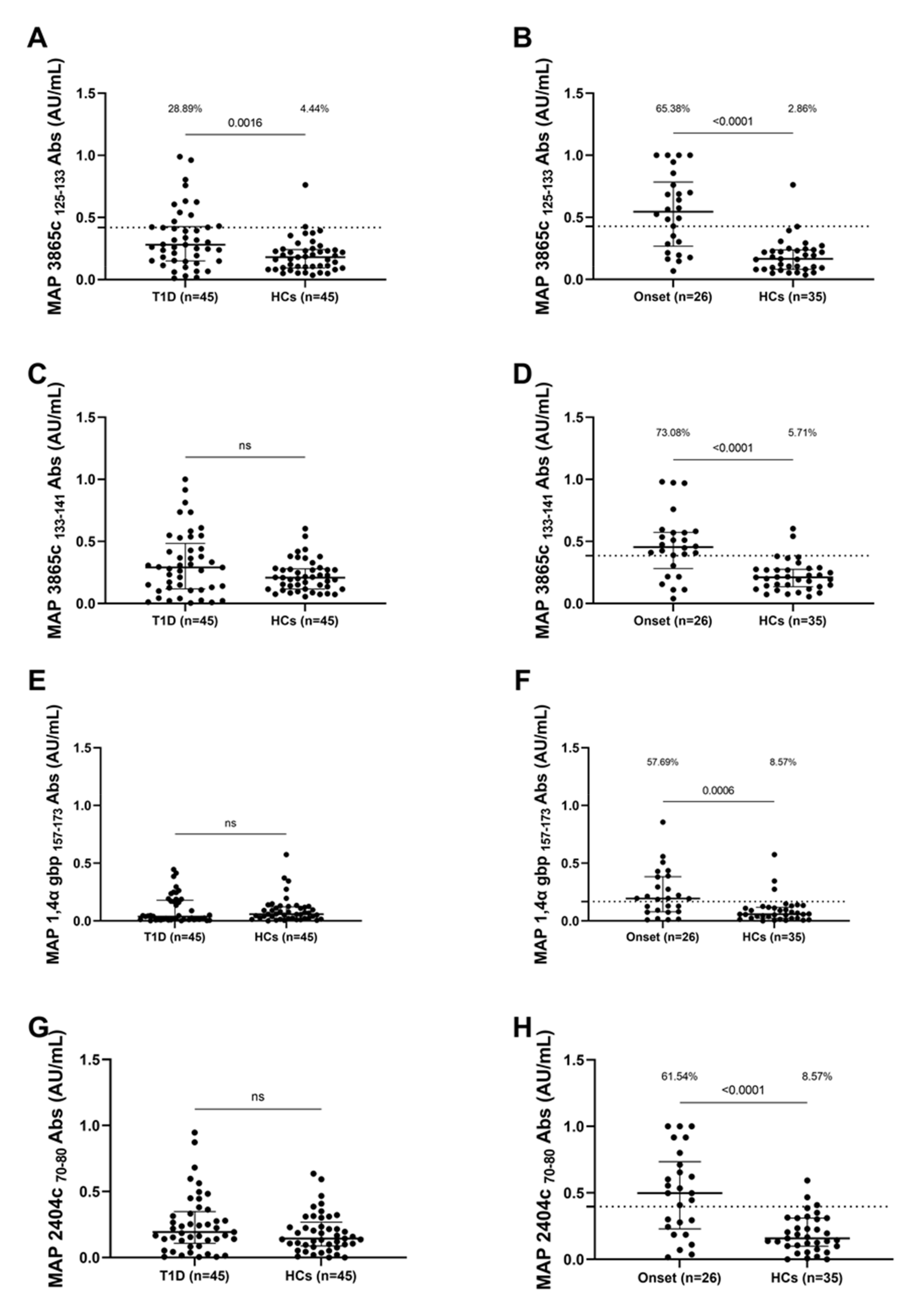
3.1. Seroreactivity against MAP Antigens
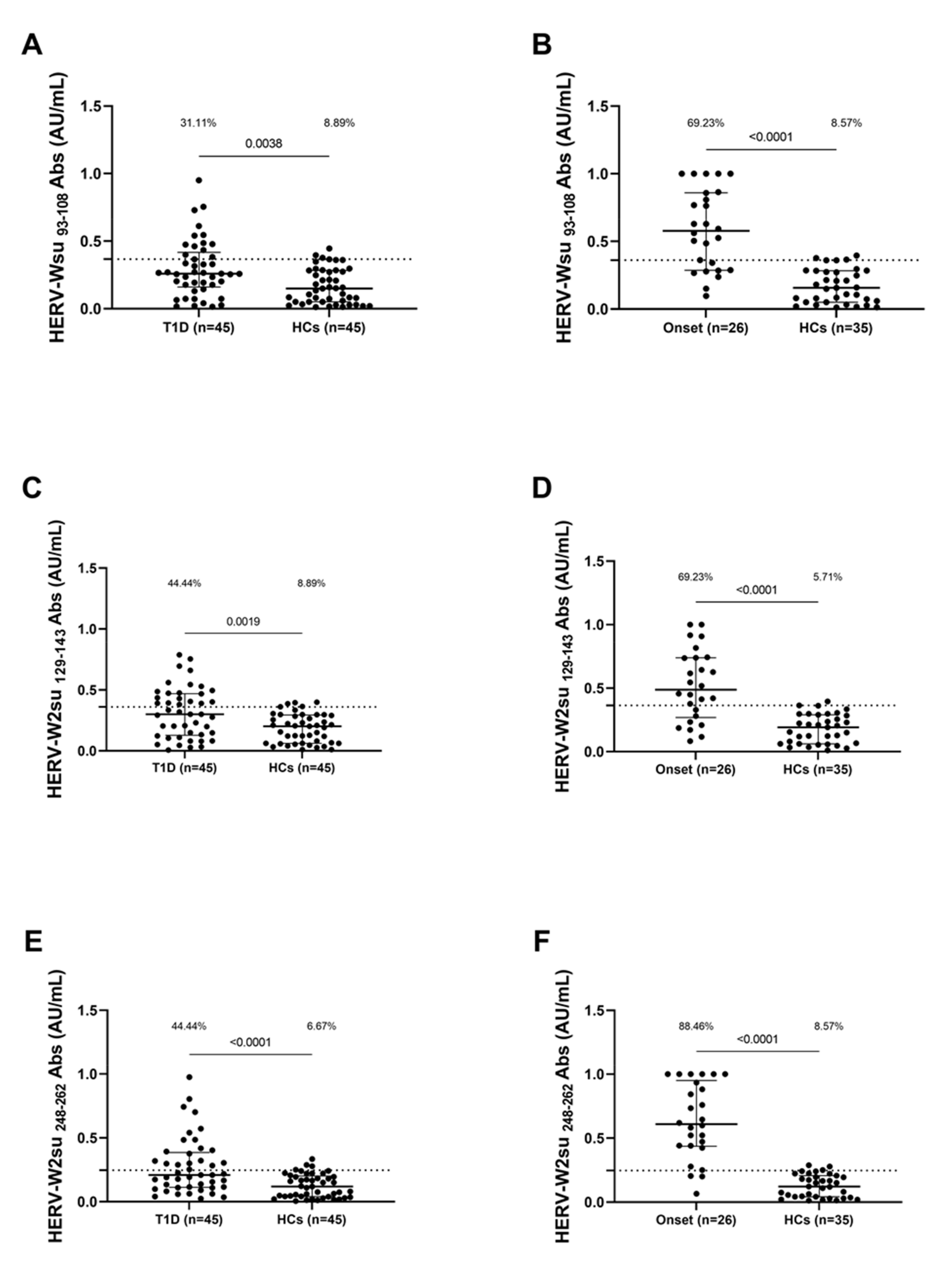
3.2. Seroreactivity against HERV-W Antigen
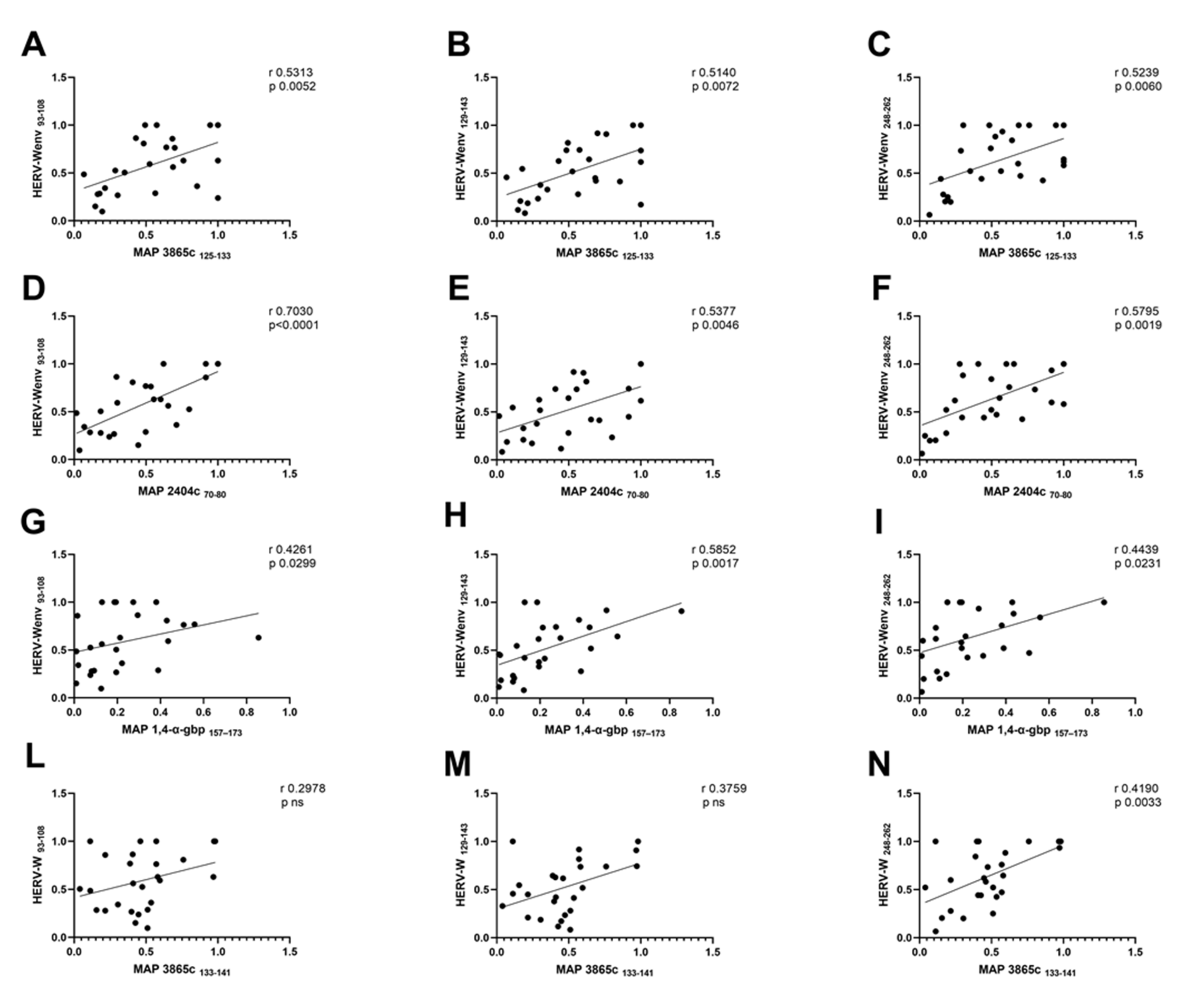
3.3. Analysis of Correlation between Anti-MAP and Anti-HERVenv Antibody Titers
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Taplin, C.; Barker, J. Autoantibodies in Type 1 Diabetes. Autoimmunity 2008, 41, 11–18. [Google Scholar] [CrossRef]
- Islam, S.T.; Srinivasan, S.; Craig, M.E. Environmental Determinants of Type 1 Diabetes: A Role for Overweight and Insulin Resistance. J. Paediatr. Child Health 2014, 50, 874–879. [Google Scholar] [CrossRef] [PubMed]
- Noble, J.A.; Valdes, A.M. Genetics of the HLA Region in the Prediction of Type 1 Diabetes. Curr. Diabetes Rep. 2011, 11, 533–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyaga, D.M.; Vickers, M.H.; Jefferies, C.; Perry, J.K.; O’Sullivan, J.M. Type 1 Diabetes Mellitus-Associated Genetic Variants Contribute to Overlapping Immune Regulatory Networks. Front. Genet. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Redondo, M.J.; Fain, P.R.; Eisenbarth, G.S. Genetics of Type 1A Diabetes. Recent Prog. Horm. Res. 2001, 56, 69–89. [Google Scholar] [CrossRef]
- Craig, M.E.; Hattersley, A.; Donaghue, K.C. Definition, Epidemiology and Classification of Diabetes in Children and Adolescents. Pediatric Diabetes 2009, 10, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Esposito, S.; Toni, G.; Tascini, G.; Santi, E.; Berioli, M.G.; Principi, N. Environmental Factors Associated with Type 1 Diabetes. Front. Endocrinol. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Knip, M.; Veijola, R.; Virtanen, S.M.; Hyöty, H.; Vaarala, O.; Åkerblom, H.K. Environmental Triggers and Determinants of Type 1 Diabetes. Diabetes 2005, 54, S125–S136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cusick, M.F.; Libbey, J.E.; Fujinami, R.S. Molecular Mimicry as a Mechanism of Autoimmune Disease. Clin. Rev. Allergy Immunol. 2012, 42, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Sechi, L.A.; Paccagnini, D.; Salza, S.; Pacifico, A.; Ahmed, N.; Zanetti, S. Mycobacterium Avium Subspecies Paratuberculosis Bacteremia in Type 1 Diabetes Mellitus: An Infectious Trigger? Clin. Infect. Dis. 2008, 46, 148–149. [Google Scholar] [CrossRef] [Green Version]
- Manca Bitti, M.L.; Masala, S.; Capasso, F.; Rapini, N.; Piccinini, S.; Angelini, F.; Pierantozzi, A.; Lidano, R.; Pietrosanti, S.; Paccagnini, D.; et al. Mycobacterium Avium Subsp. Paratuberculosis in an Italian Cohort of Type 1 Diabetes Pediatric Patients. Clin. Dev. Immunol. 2012, 2012, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Harris, N.B.; Barletta, R.G. Mycobacterium Avium Subsp. Paratuberculosis in Veterinary Medicine. Clin. Microbiol. Rev. 2001, 14, 489–512. [Google Scholar] [CrossRef] [Green Version]
- Ellingson, J.L.E.; Anderson, J.L.; Koziczkowski, J.J.; Radcliff, R.P.; Sloan, S.J.; Allen, S.E.; Sullivan, N.M. Detection of Viable Mycobacterium Avium Subsp. Paratuberculosis in Retail Pasteurized Whole Milk by Two Culture Methods and PCR. J. Food Prot. 2005, 68, 966–972. [Google Scholar] [CrossRef] [PubMed]
- Waddell, L.; Rajić, A.; Stärk, K.; McEwen, S.A. Mycobacterium Avium Ssp. Paratuberculosis Detection in Animals, Food, Water and Other Sources or Vehicles of Human Exposure: A Scoping Review of the Existing Evidence. Prev. Vet. Med. 2016, 132, 32–48. [Google Scholar] [CrossRef] [Green Version]
- Pierce, E.S. Possible Transmission of Mycobacterium Avium Subspecies Paratuberculosis through Potable Water: Lessons from an Urban Cluster of Crohn’s Disease. Gut Pathog. 2009, 1, 17. [Google Scholar] [CrossRef] [Green Version]
- Bannantine, J.P.; Bermudez, L.E. No Holes Barred: Invasion of the Intestinal Mucosa by Mycobacterium Avium Subsp. Paratuberculosis. Infect. Immun. 2013, 81, 3960–3965. [Google Scholar] [CrossRef] [Green Version]
- Rani, P.; Sechi, L.; Ahmed, N. Mycobacterium Avium Subsp. Paratuberculosis as a Trigger of Type-1 Diabetes: Destination Sardinia, or Beyond? Gut Pathog. 2010, 2, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gröger, V.; Cynis, H. Human Endogenous Retroviruses and Their Putative Role in the Development of Autoimmune Disorders Such as Multiple Sclerosis. Front. Microbiol. 2018, 9, 265. [Google Scholar] [CrossRef]
- Hansen, D.T.; Petersen, T.; Christensen, T. Retroviral Envelope Proteins: Involvement in Neuropathogenesis. J. Neurol. Sci. 2017, 380, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Lavillette, D.; Marin, M.; Ruggieri, A.; Mallet, F.; Cosset, F.-L.; Kabat, D. The Envelope Glycoprotein of Human Endogenous Retrovirus Type W Uses a Divergent Family of Amino Acid Transporters/Cell Surface Receptors. J. Virol. 2002, 76, 6442–6452. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.F.; Coffin, J.M. A Novel Endogenous Retrovirus-Related Element in the Human Genome Resembles a DNA Trans-poson: Evidence for an Evolutionary Link? Genomics 2002, 80, 453–455. [Google Scholar] [CrossRef]
- Levet, S.; Medina, J.; Joanou, J.; Demolder, A.; Queruel, N.; Réant, K.; Normand, M.; Seffals, M.; Dimier, J.; Germi, R.; et al. An Ancestral Retroviral Protein Identified as a Therapeutic Target in Type-1 Diabetes. JCI Insight 2017, 2, e94387. [Google Scholar] [CrossRef] [Green Version]
- Bashratyan, R.; Regn, D.; Rahman, M.J.; Marquardt, K.; Fink, E.; Hu, W.Y.; Elder, J.H.; Binley, J.; Sherman, L.A.; Dai, Y.D. Type 1 Diabetes Pathogenesis Is Modulated by Spontaneous Autoimmune Responses to Endogenous Retrovirus Antigens in NOD Mice. Eur. J. Immunol. 2017, 47, 575–584. [Google Scholar] [CrossRef] [Green Version]
- Curtin, F.; Bernard, C.; Levet, S.; Perron, H.; Porchet, H.; Médina, J.; Malpass, S.; Lloyd, D.; Simpson, R. A New Therapeutic Approach for Type 1 Diabetes: Rationale for GNbAC1, an Anti-HERV-W-Env Monoclonal Antibody. Diabetes Obes. Metab. 2018, 20, 2075–2084. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes Association. Diagnosis and Classification of Diabetes Mellitus. Diabetes Care 2011, 34, S62–S69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asuero, A.G.; Sayago, A.; González, A.G. The Correlation Coefficient: An Overview. Crit. Rev. Anal. Chem. 2006, 36, 41–59. [Google Scholar] [CrossRef]
- Norris, J.M.; Johnson, R.K.; Stene, L.C. Type 1 Diabetes—Early Life Origins and Changing Epidemiology. Lancet Diabetes Endocrinol. 2020, 8, 226–238. [Google Scholar] [CrossRef]
- Niegowska, M.; Wajda-Cuszlag, M.; Stępień-Ptak, G.; Trojanek, J.; Michałkiewicz, J.; Szalecki, M.; Sechi, L.A. Anti-HERV-W Env Antibodies Are Correlated with Seroreactivity against Mycobacterium Avium Subsp. Paratuberculosis in Children and Youths at T1D Risk. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Arru, G.; Caggiu, E.; Leoni, S.; Mameli, G.; Pugliatti, M.; Sechi, G.P.; Sechi, L.A. Natalizumab Modulates the Humoral Response against HERV-Wenv73-88 in a Follow-up Study of Multiple Sclerosis Patients. J. Neurol. Sci. 2015, 357, 106–108. [Google Scholar] [CrossRef] [PubMed]
- Torrado, E.; Cooper, A.M. IL-17 and Th17 Cells in Tuberculosis. Cytokine Growth Factor Rev. 2010, 21, 455–462. [Google Scholar] [CrossRef] [Green Version]
- Stephen-Victor, E.; Fickenscher, H.; Bayry, J. IL-26: An Emerging Proinflammatory Member of the IL-10 Cytokine Family with Multifaceted Actions in Antiviral, Antimicrobial, and Autoimmune Responses. PLoS Pathog. 2016, 12, 4–9. [Google Scholar] [CrossRef]
- Umemura, M.; Yahagi, A.; Hamada, S.; Begum, M.D.; Watanabe, H.; Kawakami, K.; Suda, T.; Sudo, K.; Nakae, S.; Iwakura, Y.; et al. IL-17-Mediated Regulation of Innate and Acquired Immune Response against Pulmonary Mycobacterium Bovis Bacille Calmette-Guérin Infection. J. Immunol. 2007, 178, 3786–3796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raffatellu, M.; Santos, R.L.; Verhoeven, D.E.; George, M.D.; Wilson, R.P.; Winter, S.E.; Godinez, I.; Sankaran, S.; Paixao, T.A.; Gordon, M.A.; et al. Simian Immunodeficiency Virus-Induced Mucosal Interleukin-17 Deficiency Promotes Salmonella Dissemination from the Gut. Nat. Med. 2008, 14, 421–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.E.; Park, H.T.; Jung, Y.H.; Yoo, H.S. Gene Expression Profiles of Immune-Regulatory Genes in Whole Blood of Cattle with a Subclinical Infection of Mycobacterium Avium Subsp. Paratuberculosis. PLoS ONE 2018, 13, e0196502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munn, D.H.; Sharma, M.D.; Baban, B.; Harding, H.P.; Zhang, Y.; Ron, D.; Mellor, A.L. GCN2 Kinase in T Cells Mediates Proliferative Arrest and Anergy Induction in Response to Indoleamine 2,3-Dioxygenase. Immunity 2005, 22, 633–642. [Google Scholar] [CrossRef] [Green Version]
- Nellåker, C.; Yao, Y.; Jones-Brando, L.; Mallet, F.; Yolken, R.H.; Karlsson, H. Transactivation of Elements in the Human Endogenous Retrovirus W Family by Viral Infection. Retrovirology 2006, 3, 1–11. [Google Scholar] [CrossRef]
- Morris, G.; Maes, M.; Murdjeva, M.; Puri, B.K. Do Human Endogenous Retroviruses Contribute to Multiple Sclerosis, and If So, How? Mol. Neurobiol. 2019, 56, 2590–2605. [Google Scholar] [CrossRef] [Green Version]
- Niegowska, M.; Rapini, N.; Piccinini, S.; Mameli, G.; Caggiu, E.; Bitti, M.L.M.; Sechi, L.A. Type 1 Diabetes At-Risk Children Highly Recognize Mycobacterium Avium Subspecies Paratuberculosis Epitopes Homologous to Human Znt8 and Proinsulin. Sci. Rep. 2016, 6, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Masala, S.; Cossu, D.; Piccinini, S.; Rapini, N.; Massimi, A.; Porzio, O.; Pietrosanti, S.; Lidano, R.; Bitti, M.L.M.; Sechi, L.A. Recognition of Zinc Transporter 8 and MAP3865c Homologous Epitopes by New-Onset Type 1 Diabetes Children from Continental Italy. Acta Diabetol. 2014, 51, 577–585. [Google Scholar] [CrossRef]
- Qasem, A.; Abdel-Aty, A.; Abu-Suwa, H.; Naser, S.A. Oxidative Stress Due to Mycobacterium Avium Subspecies Paratubercu losis (MAP) Infection Upregulates Selenium-Dependent GPx Activity. Gut Pathog. 2016, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Valheim, M.; Siguroardóttir, Ó.G.; Storset, A.K.; Aune, L.G.; Press, C.M.L. Characterization of Macrophages and Occurrence of Cells in Intestinal Lesions of Subclinical Paratuberculosis in Goats. J. Comp. Pathol. 2004, 131, 221–232. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Peptides | Position | Sequence |
|---|---|---|
| MAP 3865c | aa 125–133 | MIAVALAGL |
| MAP 3865c | aa 133–141 | LAANFVVAL |
| MAP 2404c | aa 70–80 | RGFVVLPVTRRDVTDV |
| MAP 1,4-α-gbp | aa 157–173 | GTVELLGGPLAHPFQPL |
| HERV-Wenv | aa 93–108 | NPSCPGGLGVTVCWTY |
| HERV-Wenv | aa 129–143 | VKEVISQLTRVRHGT |
| HERV-Wenv | aa 248–262 | NSQCIRWVTPPTQIV |
| MAP Antigen | HERV-Wenv 93–108 | HERV-Wenv 129–143 | HERV-Wenv 248–262 |
|---|---|---|---|
| MAP 3865c 125–133 | r = 0.5313 p = 0.0052 | r = 0.5140 p = 0.0072 | r = 0.5239 p = 0.0060 |
| MAP 2404c 70–80 | r = 0.7030 p < 0.0001 | r = 0.5377 p = 0.0046 | r = 0.5795 p = 0.0019 |
| MAP 1,4-α-gbp 157–173 | r = 0.4261 p = 0.0299 | r = 0.5852 p = 0.0017 | r = 0.4439 p = 0.0213 |
| MAP 3865c 133–141 | r = 0.2978 p ns | r = 0.3759 p ns | r = 0.4190 p = 0.0311 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noli, M.; Meloni, G.; Manca, P.; Cossu, D.; Palermo, M.; Sechi, L.A. HERV-W and Mycobacterium avium subspecies paratuberculosis Are at Play in Pediatric Patients at Onset of Type 1 Diabetes. Pathogens 2021, 10, 1135. https://doi.org/10.3390/pathogens10091135
Noli M, Meloni G, Manca P, Cossu D, Palermo M, Sechi LA. HERV-W and Mycobacterium avium subspecies paratuberculosis Are at Play in Pediatric Patients at Onset of Type 1 Diabetes. Pathogens. 2021; 10(9):1135. https://doi.org/10.3390/pathogens10091135
Chicago/Turabian StyleNoli, Marta, Gianfranco Meloni, Pietro Manca, Davide Cossu, Mario Palermo, and Leonardo A. Sechi. 2021. "HERV-W and Mycobacterium avium subspecies paratuberculosis Are at Play in Pediatric Patients at Onset of Type 1 Diabetes" Pathogens 10, no. 9: 1135. https://doi.org/10.3390/pathogens10091135
APA StyleNoli, M., Meloni, G., Manca, P., Cossu, D., Palermo, M., & Sechi, L. A. (2021). HERV-W and Mycobacterium avium subspecies paratuberculosis Are at Play in Pediatric Patients at Onset of Type 1 Diabetes. Pathogens, 10(9), 1135. https://doi.org/10.3390/pathogens10091135