
Role of Helicobacter pylori and Other Environmental Factors in the Development of Gastric Dysbiosis

 ,
,  , ,
, ,
Abstract
:1. Introduction
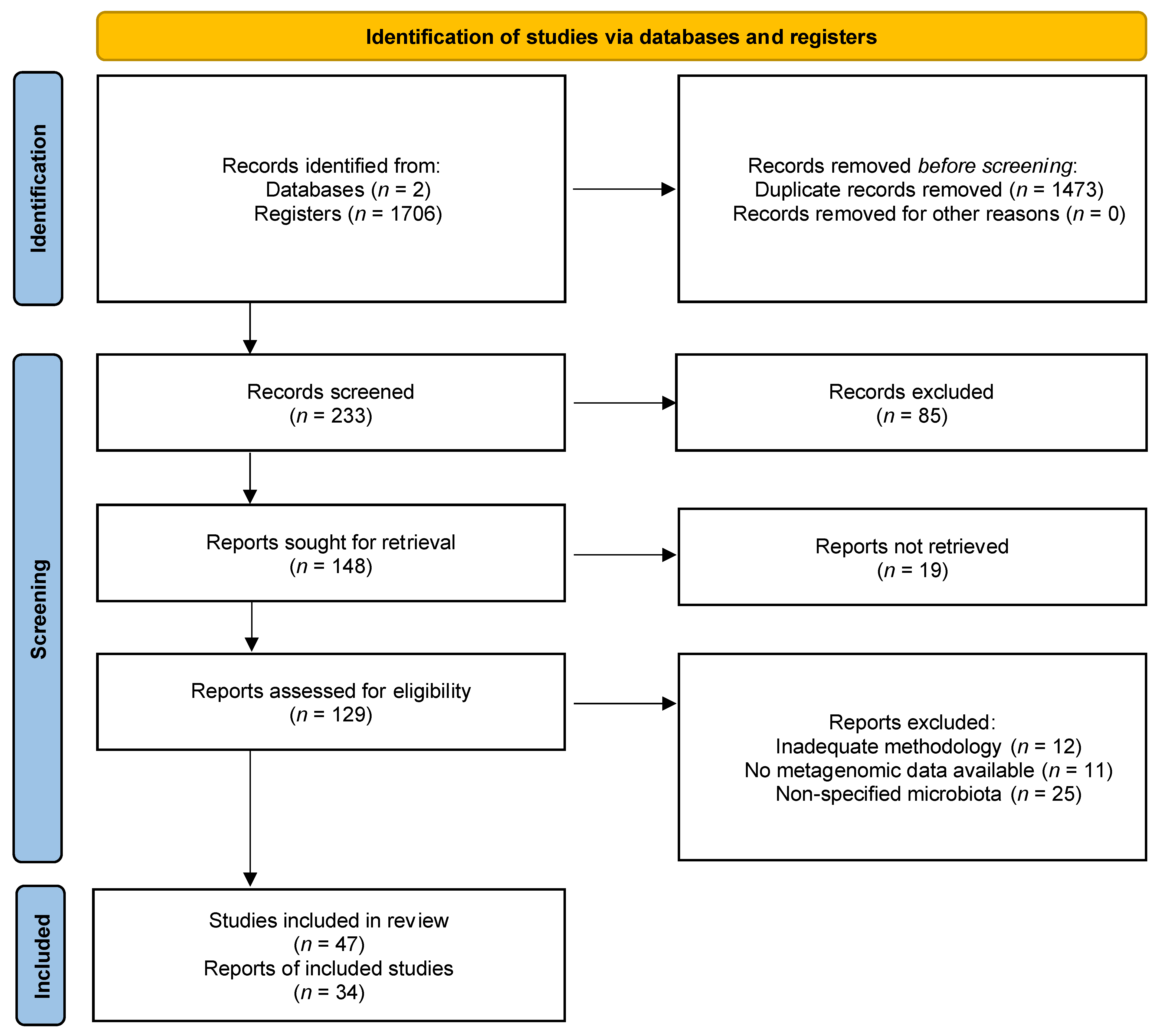
2. Methodology
2.1. Study Design
2.2. Article Research
2.3. Selection of Studies
2.4. Data Extraction
3. Main Findings and Discussion
3.1. Gastrointestinal Microbiota
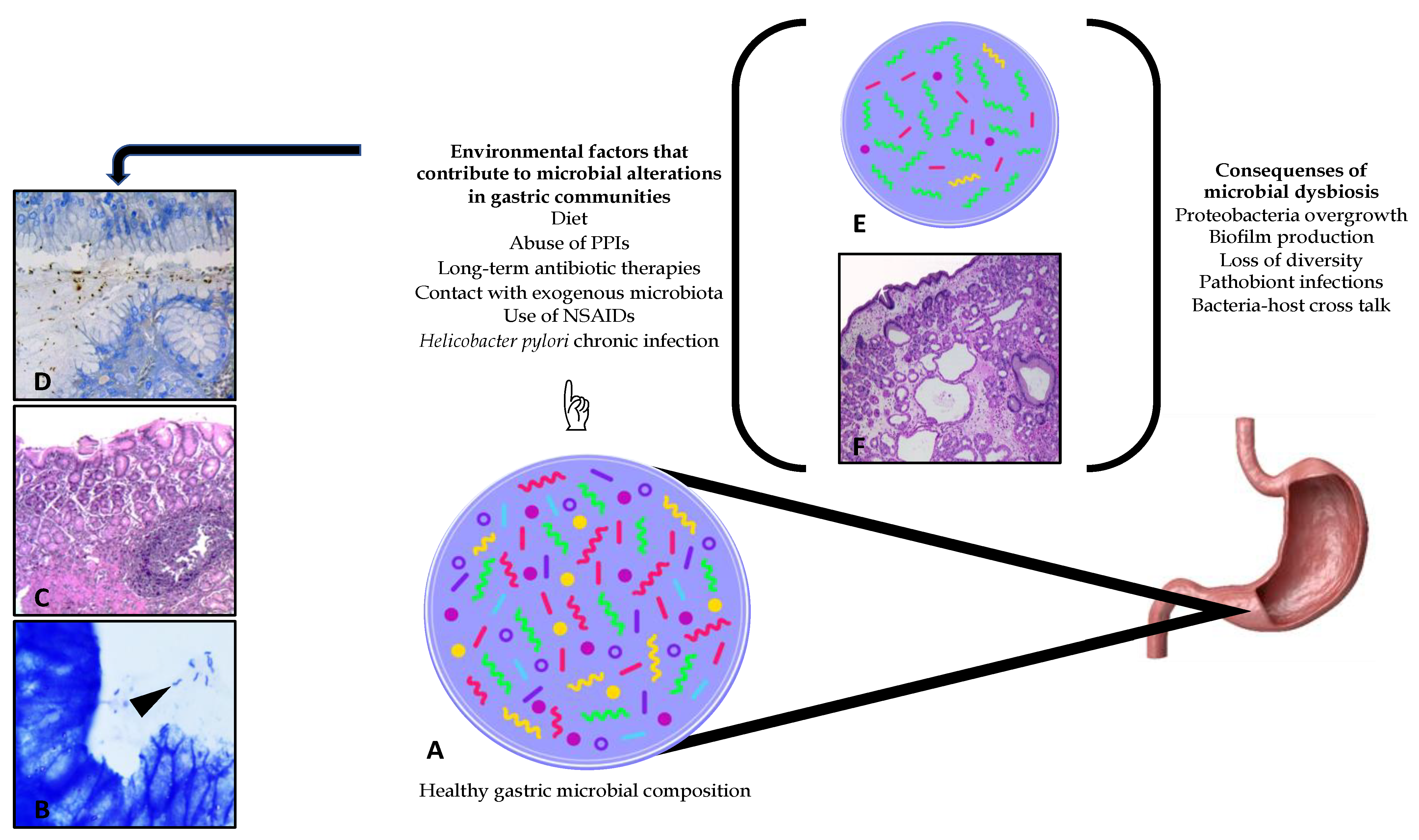
3.2. Gastric Microbiota
3.3. Dysbiosis Generated in the Gastric Microenvironment by Helicobacter pylori Status Diseases
3.3.1. Dysbiosis
3.3.2. Helicobacter pylori
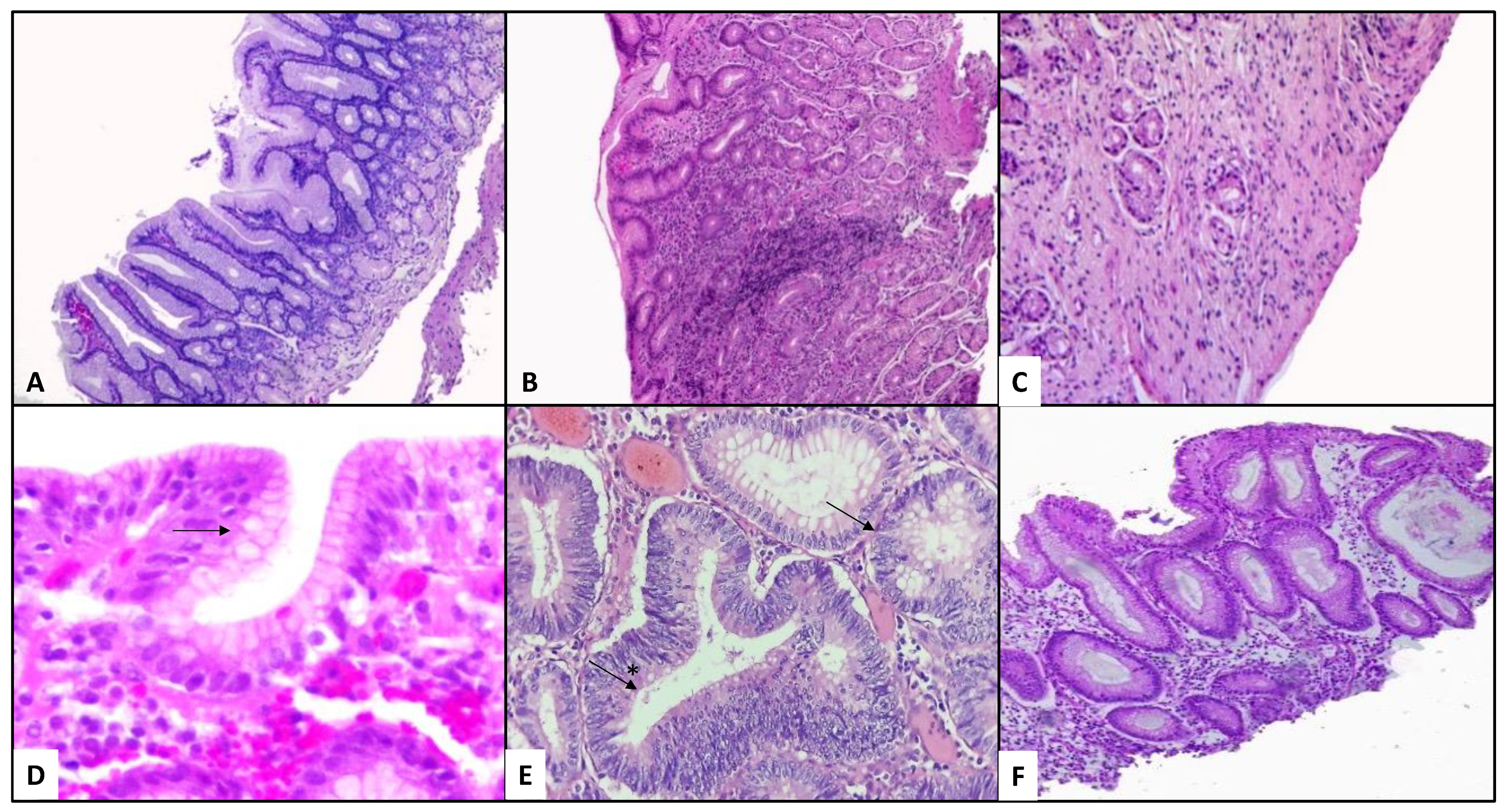
3.3.3. Gastric Dysbiosis in Patients with Gastritis and H. pylori Variable Status
3.3.4. Dysbiosis in Patients with Gastric Cancer and H. pylori Variable Status
3.3.5. Alterations in the Gastric Microbiota after Multiple Therapeutic Interventions
3.3.6. Abuse of Proton Pump Inhibitors
3.3.7. Use of Antibiotics
3.3.8. H. pylori Eradication Treatments
3.3.9. Microbiota Recovery after Antibiotic Eradication Treatment
3.3.10. Probiotics
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lederberg, J.; McCray, A.T. ‘Ome Sweet’ Omics—A Genealogical Treasury of Words. Sci. 2001, 15, 8. [Google Scholar]
- Beirão, E.M.; Padovan, A.C.B.; Furtado, J.J.D.; Colombo, A.L.; Medeiros, E.A.S. Does the Change on Gastrointestinal Tract Microbiome Affects Host? Braz. J. Infect. Dis. 2014, 18, 660–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buelow, E.; Bello González, T.D.J.; Fuentes, S.; de Steenhuijsen Piters, W.A.A.; Lahti, L.; Bayjanov, J.R.; Majoor, E.A.M.; Braat, J.C.; van Mourik, M.S.M.; Oostdijk, E.A.N.; et al. Comparative Gut Microbiota and Resistome Profiling of Intensive Care Patients Receiving Selective Digestive Tract Decontamination and Healthy Subjects. Microbiome 2017, 5, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, V.B.; Schmidt, T.M. Overview of the Gastrointestinal Microbiota. Adv. Exp. Med. Biol. 2008, 635, 29–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klymiuk, I.; Bilgilier, C.; Stadlmann, A.; Thannesberger, J.; Kastner, M.-T.; Högenauer, C.; Püspök, A.; Biowski-Frotz, S.; Schrutka-Kölbl, C.; Thallinger, G.G.; et al. The Human Gastric Microbiome Is Predicated upon Infection with Helicobacter pylori. Front. Microbiol. 2017, 8, 2508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillman, E.T.; Lu, H.; Yao, T.; Nakatsu, C.H. Microbial Ecology along the Gastrointestinal Tract. Microbes Environ. 2017, 32, 300–313. [Google Scholar] [CrossRef] [Green Version]
- Hung, I.F.N.; Wong, B.C.Y. Assessing the Risks and Benefits of Treating Helicobacter pylori Infection. Ther. Adv. Gastroenterol. 2009, 2, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Reshetnyak, V.I.; Burmistrov, A.I.; Maev, I.V. Helicobacter pylori: Commensal, Symbiont or Pathogen? World J. Gastroenterol. 2021, 27, 545–560. [Google Scholar] [CrossRef] [PubMed]
- Haddaway, N.R.; McGuinness, L.A.; Pritchard, C.C. PRISMA2020: R Package and ShinyApp for Producing PRISMA 2020 Compliant Flow Diagrams (Version 0.0.2); Zenodo: Geneve, Switzerland, 2021. [Google Scholar] [CrossRef]
- Belizário, J.E.; Napolitano, M. Human Microbiomes and Their Roles in Dysbiosis, Common Diseases, and Novel Therapeutic Approaches. Front. Microbiol. 2015, 6, 1050. [Google Scholar] [CrossRef] [Green Version]
- Bik, E.M.; Eckburg, P.B.; Gill, S.R.; Nelson, K.E.; Purdom, E.A.; Francois, F.; Perez-Perez, G.; Blaser, M.J.; Relman, D.A. Molecular Analysis of the Bacterial Microbiota in the Human Stomach. Proc. Natl. Acad. Sci. USA 2006, 103, 732–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.-X.; Wong, G.L.-H.; To, K.-F.; Wong, V.W.-S.; Lai, L.H.; Chow, D.K.-L.; Lau, J.Y.-W.; Sung, J.J.-Y.; Ding, C. Bacterial Microbiota Profiling in Gastritis without Helicobacter pylori Infection or Non-Steroidal Anti-Inflammatory Drug Use. PLoS ONE 2009, 4, e7985. [Google Scholar] [CrossRef]
- Rajilić-Stojanović, M.; de Vos, W.M. The First 1000 Cultured Species of the Human Gastrointestinal Microbiota. FEMS Microbiol. Rev. 2014, 38, 996–1047. [Google Scholar] [CrossRef]
- Mailhe, M.; Ricaboni, D.; Vitton, V.; Gonzalez, J.-M.; Bachar, D.; Dubourg, G.; Cadoret, F.; Robert, C.; Delerce, J.; Levasseur, A.; et al. Repertoire of the Gut Microbiota from Stomach to Colon Using Culturomics and Next-Generation Sequencing. BMC Microbiol. 2018, 18, 157. [Google Scholar] [CrossRef]
- Petra, C.V.; Rus, A.; Dumitraşcu, D.L. Gastric Microbiota: Tracing the Culprit. Clujul Med. 2017, 90, 369–376. [Google Scholar] [CrossRef] [Green Version]
- Stearns, J.C.; Lynch, M.D.J.; Senadheera, D.B.; Tenenbaum, H.C.; Goldberg, M.B.; Cvitkovitch, D.G.; Croitoru, K.; Moreno-Hagelsieb, G.; Neufeld, J.D. Bacterial Biogeography of the Human Digestive Tract. Sci. Rep. 2011, 1, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekaboruah, E.; Suryavanshi, M.V.; Chettri, D.; Verma, A.K. Human Microbiome: An Academic Update on Human Body Site Specific Surveillance and Its Possible Role. Arch. Microbiol. 2020, 202, 2147–2167. [Google Scholar] [CrossRef] [PubMed]
- Rajilic-Stojanovic, M.; Figueiredo, C.; Smet, A.; Hansen, R.; Kupcinskas, J.; Rokkas, T.; Andersen, L.; Machado, J.C.; Ianiro, G.; Gasbarrini, A.; et al. Systematic Review: Gastric Microbiota in Health and Disease. Aliment. Pharmacol. Ther. 2020, 51, 582–602. [Google Scholar] [CrossRef]
- Noto, J.M.; Peek, R.M.J. The Gastric Microbiome, Its Interaction with Helicobacter pylori, and Its Potential Role in the Progression to Stomach Cancer. PLoS Pathog. 2017, 13, e1006573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.-C.; Lu, C.-W.; Lin, C.-J. Treatment of Helicobacter pylori Infection: Current Status and Future Concepts. World J. Gastroenterol. 2014, 20, 5283–5293. [Google Scholar] [CrossRef]
- Linz, B.; Balloux, F.; Moodley, Y.; Manica, A.; Liu, H.; Roumagnac, P.; Falush, D.; Stamer, C.; Prugnolle, F.; van der Merwe, S.W.; et al. An African Origin for the Intimate Association between Humans and Helicobacter pylori. Nature 2007, 445, 915–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moodley, Y.; Linz, B.; Bond, R.P.; Nieuwoudt, M.; Soodyall, H.; Schlebusch, C.M.; Bernhöft, S.; Hale, J.; Suerbaum, S.; Mugisha, L.; et al. Age of the Association between Helicobacter pylori and Man. PLoS Pathog. 2012, 8, e1002693. [Google Scholar] [CrossRef] [Green Version]
- Chey, W.D.; Leontiadis, G.I.; Howden, C.W.; Moss, S.F. ACG Clinical Guideline: Treatment of Helicobacter pylori Infection. Am. J. Gastroenterol. 2017, 112, 212–239. [Google Scholar] [CrossRef] [PubMed]
- Wroblewski, L.E.; Peek, R.M., Jr.; Wilson, K.T. Helicobacter pylori and Gastric Cancer: Factors That Modulate Disease Risk. Clin. Microbiol. Rev. 2010, 23, 713–739. [Google Scholar] [CrossRef] [Green Version]
- Atherton, J.C.; Blaser, M.J. Coadaptation of Helicobacter pylori and Humans: Ancient History, Modern Implications. J. Clin. Investig. 2009, 119, 2475–2487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maldonado-Contreras, A.; Goldfarb, K.C.; Godoy-Vitorino, F.; Karaoz, U.; Contreras, M.; Blaser, M.J.; Brodie, E.L.; Dominguez-Bello, M.G. Structure of the Human Gastric Bacterial Community in Relation to Helicobacter pylori Status. ISME J. 2011, 5, 574–579. [Google Scholar] [CrossRef] [PubMed]
- Eun, C.S.; Kim, B.K.; Han, D.S.; Kim, S.Y.; Kim, K.M.; Choi, B.Y.; Song, K.S.; Kim, Y.S.; Kim, J.F. Differences in Gastric Mucosal Microbiota Profiling in Patients with Chronic Gastritis, Intestinal Metaplasia, and Gastric Cancer Using Pyrosequencing Methods. Helicobacter 2014, 19, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Sipponen, P.; Maaroos, H.-I. Chronic Gastritis. Scand. J. Gastroenterol. 2015, 50, 657–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strickland, R.G. Gastritis. Front. Gastrointest. Res. 1975, 1, 12–48. [Google Scholar] [CrossRef]
- Dixon, M.F. Pathology of Gastritis and Peptic Ulceration. In Helicobacter Pylori: Physiology and Genetics; Mobley, H.L., Mendz, G.L., Hazell, S.L., Eds.; ASM Press: Washington, DC, USA, 2001. [Google Scholar]
- Watari, J.; Chen, N.; Amenta, P.S.; Fukui, H.; Oshima, T.; Tomita, T.; Miwa, H.; Lim, K.-J.; Das, K.M. Helicobacter pylori Associated Chronic Gastritis, Clinical Syndromes, Precancerous Lesions, and Pathogenesis of Gastric Cancer Development. World J. Gastroenterol. 2014, 20, 5461–5473. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, J.; Xu, J.; Wei, X.; Yang, J.; Liu, Y.; Li, H.; Zhao, C.; Wang, Y.; Zhang, L.; et al. Helicobacter pylori Infection Aggravates Dysbiosis of Gut Microbiome in Children with Gastritis. Front. Cell Infect. Microbiol. 2019, 9, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aviles-Jimenez, F.; Vazquez-Jimenez, F.; Medrano-Guzman, R.; Mantilla, A.; Torres, J. Stomach Microbiota Composition Varies between Patients with Non-Atrophic Gastritis and Patients with Intestinal Type of Gastric Cancer. Sci. Rep. 2014, 4, 4202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, L.; Yu, J. Effect of Helicobacter pylori Infection on the Composition of Gastric Microbiota in the Development of Gastric Cancer. Gastrointest. Tumors 2015, 2, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Sugano, K.; Tack, J.; Kuipers, E.J.; Graham, D.Y.; El-Omar, E.M.; Miura, S.; Haruma, K.; Asaka, M.; Uemura, N.; Malfertheiner, P. Kyoto Global Consensus Report on Helicobacter pylori Gastritis. Gut 2015, 64, 1353–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meining, A.; Stolte, M.; Hatz, R.; Lehn, N.; Miehlke, S.; Morgner, A.; Bayerdörffer, E. Differing Degree and Distribution of Gastritis in Helicobacter pylori-Associated Diseases. Virchows Arch. 1997, 431, 11–15. [Google Scholar] [CrossRef]
- Watabe, H.; Mitsushima, T.; Yamaji, Y.; Okamoto, M.; Wada, R.; Kokubo, T.; Doi, H.; Yoshida, H.; Kawabe, T.; Omata, M. Predicting the Development of Gastric Cancer from Combining Helicobacter pylori Antibodies and Serum Pepsinogen Status: A Prospective Endoscopic Cohort Study. Gut 2005, 54, 764–768. [Google Scholar] [CrossRef] [Green Version]
- Naylor, G.M.; Gotoda, T.; Dixon, M.; Shimoda, T.; Gatta, L.; Owen, R.; Tompkins, D.; Axon, A. Why Does Japan Have a High Incidence of Gastric Cancer? Comparison of Gastritis between UK and Japanese Patients. Gut 2006, 55, 1545–1552. [Google Scholar] [CrossRef] [Green Version]
- Rugge, M.; de Boni, M.; Pennelli, G.; de Bona, M.; Giacomelli, L.; Fassan, M.; Basso, D.; Plebani, M.; Graham, D.Y. Gastritis OLGA-Staging and Gastric Cancer Risk: A Twelve-Year Clinico-Pathological Follow-up Study. Aliment. Pharmacol. Ther. 2010, 31, 1104–1111. [Google Scholar] [CrossRef]
- den Hoed, C.M.; Holster, I.L.; Capelle, L.G.; de Vries, A.C.; den Hartog, B.; Ter Borg, F.; Biermann, K.; Kuipers, E.J. Follow-up of Premalignant Lesions in Patients at Risk for Progression to Gastric Cancer. Endoscopy 2013, 45, 249–256. [Google Scholar] [CrossRef]
- Ahn, H.J.; Lee, D.S. Helicobacter pylori in Gastric Carcinogenesis. World J. Gastrointest. Oncol. 2015, 7, 455–465. [Google Scholar] [CrossRef]
- Ferreira, R.M.; Pereira-Marques, J.; Pinto-Ribeiro, I.; Costa, J.L.; Carneiro, F.; Machado, J.C.; Figueiredo, C. Gastric Microbial Community Profiling Reveals a Dysbiotic Cancer-Associated Microbiota. Gut 2018, 67, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Castaño-Rodríguez, N.; Goh, K.-L.; Fock, K.M.; Mitchell, H.M.; Kaakoush, N.O. Dysbiosis of the Microbiome in Gastric Carcinogenesis. Sci. Rep. 2017, 7, 15957. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Shao, L.; Liu, X.; Ji, F.; Mei, Y.; Cheng, Y.; Liu, F.; Yan, C.; Li, L.; Ling, Z. Alterations of Gastric Mucosal Microbiota across Different Stomach Microhabitats in a Cohort of 276 Patients with Gastric Cancer. EBioMedicine 2019, 40, 336–348. [Google Scholar] [CrossRef] [Green Version]
- Gunathilake, M.N.; Lee, J.; Choi, I.J.; Kim, Y.-I.; Ahn, Y.; Park, C.; Kim, J. Association between the Relative Abundance of Gastric Microbiota and the Risk of Gastric Cancer: A Case-Control Study. Sci. Rep. 2019, 9, 13589. [Google Scholar] [CrossRef] [PubMed]
- Take, S.; Mizuno, M.; Ishiki, K.; Nagahara, Y.; Yoshida, T.; Yokota, K.; Oguma, K. Baseline Gastric Mucosal Atrophy Is a Risk Factor Associated with the Development of Gastric Cancer after Helicobacter pylori Eradication Therapy in Patients with Peptic Ulcer Diseases. J. Gastroenterol. 2007, 42 (Suppl. S17), 21–27. [Google Scholar] [CrossRef] [PubMed]
- Ford, A.C.; Forman, D.; Hunt, R.H.; Yuan, Y.; Moayyedi, P. Helicobacter pylori Eradication Therapy to Prevent Gastric Cancer in Healthy Asymptomatic Infected Individuals: Systematic Review and Meta-Analysis of Randomised Controlled Trials. BMJ 2014, 348, g3174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Xu, W.; Lee, A.; He, J.; Huang, B.; Zheng, W.; Su, T.; Lai, S.; Long, Y.; Chu, H.; et al. The Impact of Helicobacter pylori Infection, Eradication Therapy and Probiotic Supplementation on Gut Microenvironment Homeostasis: An Open-Label, Randomized Clinical Trial. EBioMedicine 2018, 35, 87–96. [Google Scholar] [CrossRef] [Green Version]
- de Vries, A.C.; Kuipers, E.J.; Rauws, E.A.J. Helicobacter pylori Eradication and Gastric Cancer: When Is the Horse out of the Barn? Am. J. Gastroenterol. 2009, 104, 1342–1345. [Google Scholar] [CrossRef]
- Shin, J.M.; Kim, N. Pharmacokinetics and Pharmacodynamics of the Proton Pump Inhibitors. J. Neurogastroenterol. Motil. 2013, 19, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Parsons, B.N.; Ijaz, U.Z.; D’Amore, R.; Burkitt, M.D.; Eccles, R.; Lenzi, L.; Duckworth, C.A.; Moore, A.R.; Tiszlavicz, L.; Varro, A.; et al. Comparison of the Human Gastric Microbiota in Hypochlorhydric States Arising as a Result of Helicobacter pylori-Induced Atrophic Gastritis, Autoimmune Atrophic Gastritis and Proton Pump Inhibitor Use. PLoS Pathog. 2017, 13, e1006653. [Google Scholar] [CrossRef] [Green Version]
- Paroni Sterbini, F.; Palladini, A.; Masucci, L.; Cannistraci, C.V.; Pastorino, R.; Ianiro, G.; Bugli, F.; Martini, C.; Ricciardi, W.; Gasbarrini, A.; et al. Effects of Proton Pump Inhibitors on the Gastric Mucosa-Associated Microbiota in Dyspeptic Patients. Appl. Environ. Microbiol. 2016, 82, 6633–6644. [Google Scholar] [CrossRef] [Green Version]
- Davies, J.; Davies, D. Origins and Evolution of Antibiotic Resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [Green Version]
- Bobo, D.; Lipatov, M.; Rodriguez-Flores, J.L.; Auton, A.; Henn, B.M. False Negatives Are a Significant Feature of Next Generation Sequencing Callsets. Biorxiv 2016, 066043. [Google Scholar] [CrossRef] [Green Version]
- Birg, A.; Ritz, N.; Lin, H.C. The Unknown Effect of Antibiotic-Induced Dysbiosis on the Gut Microbiota. In Microbiome and Metabolome in Diagnosis, Therapy, and Other Strategic Applications; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Rathinavelu, S.; Zavros, Y.; Merchant, J.L. Acinetobacter Lwoffii Infection and Gastritis. Microbes Infect. 2003, 5, 651–657. [Google Scholar] [CrossRef]
- Dias-Jácome, E.; Libânio, D.; Borges-Canha, M.; Galaghar, A.; Pimentel-Nunes, P. Gastric Microbiota and Carcinogenesis: The Role of Non-Helicobacter pylori Bacteria—A Systematic Review. Rev. Esp. Enferm. Dig. 2016, 108, 530–540. [Google Scholar] [CrossRef] [Green Version]
- Delgado, S.; Suárez, A.; Mayo, B. Identification, Typing and Characterisation of Propionibacterium Strains from Healthy Mucosa of the Human Stomach. Int. J. Food Microbiol. 2011, 149, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montalban-Arques, A.; Wurm, P.; Trajanoski, S.; Schauer, S.; Kienesberger, S.; Halwachs, B.; Högenauer, C.; Langner, C.; Gorkiewicz, G. Propionibacterium Acnes Overabundance and Natural Killer Group 2 Member D System Activation in Corpus-Dominant Lymphocytic Gastritis. J. Pathol. 2016, 240, 425–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dréno, B.; Pécastaings, S.; Corvec, S.; Veraldi, S.; Khammari, A.; Roques, C. Cutibacterium Acnes (Propionibacterium Acnes) and Acne Vulgaris: A Brief Look at the Latest Updates. J. Eur. Acad. Dermatol. Venereol. 2018, 32 (Suppl. S2), 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.; Ramesh, A. Bacteriocin-Producing Strains of Lactobacillus Plantarum Inhibit Adhesion of Staphylococcus Aureus to Extracellular Matrix: Quantitative Insight and Implications in Antibacterial Therapy. J. Med. Microbiol. 2015, 64, 1514–1526. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhou, J.; Xin, Y.; Geng, C.; Tian, Z.; Yu, X.; Dong, Q. Bacterial Overgrowth and Diversification of Microbiota in Gastric Cancer. Eur. J. Gastroenterol. Hepatol. 2016, 28, 261–266. [Google Scholar] [CrossRef]
- Bratcher, D.F. 131—Other Corynebacteria. In Principles and Practice of Pediatric Infectious Diseases, 5th ed.; Long, S.S., Prober, C.G., Fischer, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 778–781.e2. [Google Scholar] [CrossRef]
- Tally, F.P.; Sullivan, C.E. Metronidazole: In Vitro Activity, Pharmacology and Efficacy in Anaerobic Bacterial Infections. Pharmacotherapy 1981, 1, 28–38. [Google Scholar] [CrossRef]
- Uemura, N.; Okamoto, S.; Yamamoto, S.; Matsumura, N.; Yamaguchi, S.; Yamakido, M.; Taniyama, K.; Sasaki, N.; Schlemper, R.J. Helicobacter pylori Infection and the Development of Gastric Cancer. N. Engl. J. Med. 2001, 345, 784–789. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.J.Y.; Coker, O.O.; Chu, E.; Szeto, C.H.; Luk, S.T.Y.; Lau, H.C.H.; Yu, J. Gastric Microbes Associated with Gastric Inflammation, Atrophy and Intestinal Metaplasia 1 Year after Helicobacter pylori Eradication. Gut 2020, 69, 1572–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamsson, I.; Nord, C.E.; Lundquist, P.; Sjöstedt, S.; Edlund, C. Comparative Effects of Omeprazole, Amoxycillin plus Metronidazole versus Omeprazole, Clarithromycin plus Metronidazole on the Oral, Gastric and Intestinal Microflora in Helicobacter pylori-Infected Patients. J. Antimicrob. Chemother. 1999, 44, 629–640. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhou, X.; Xiao, S.; Ye, F.; Zhang, G. The Effect of Helicobacter pylori Eradication on the Gastrointestinal Microbiota in Patients with Duodenal Ulcer. J. Gastrointestin. Liver Dis. 2016, 25, 139–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.H.; Qin, Y.; Sham, P.C.; Lau, K.S.; Chu, K.-M.; Leung, W.K. Alterations in Gastric Microbiota After H. Pylori Eradication and in Different Histological Stages of Gastric Carcinogenesis. Sci. Rep. 2017, 7, 44935. [Google Scholar] [CrossRef] [PubMed]
- Olekhnovich, E.; Manolov, A.; Samoilov, A.; Prianichnikov, N.; Malakhova, M.; Tyakht, A.; Pavlenko, A.; Babenko, V.; Larin, A.; Kovarsky, B.; et al. Shifts in the Human Gut Microbiota Structure Caused by Quadruple Helicobacter pylori Eradication Therapy. Front. Microbiol. 2019, 10, 1902. [Google Scholar] [CrossRef] [Green Version]
- Shaw, L.P.; Bassam, H.; Barnes, C.P.; Walker, A.S.; Klein, N.; Balloux, F. Modelling Microbiome Recovery after Antibiotics Using a Stability Landscape Framework. ISME J. 2019, 13, 1845–1856. [Google Scholar] [CrossRef] [Green Version]
- Silva, D.; Sardi, J.; Pitangui, N.; MagreRoque, S.; Silva, A.; Rosalen, P. Probiotics as an Alternative Antimicrobial Therapy: Current Reality and Future Directions. J. Funct. Foods 2020, 73. [Google Scholar] [CrossRef]
- Cunningham, M.; Azcarate-Peril, M.A.; Barnard, A.; Benoit, V.; Grimaldi, R.; Guyonnet, D.; Holscher, H.D.; Hunter, K.; Manurung, S.; Obis, D.; et al. Shaping the Future of Probiotics and Prebiotics. Trends Microbiol. 2021, 29, 667–685. [Google Scholar] [CrossRef]
- Ademe, M. Benefits of Fecal Microbiota Transplantation: A Comprehensive Review. J. Infect. Dev. Ctries. 2020, 14, 1074–1080. [Google Scholar] [CrossRef]
- Marrs, T.; Walter, J. Pros and Cons: Is Faecal Microbiota Transplantation a Safe and Efficient Treatment Option for Gut Dysbiosis? Allergy 2021, 76, 2312–2317. [Google Scholar] [CrossRef]
- Ji, J.; Yang, H. Using Probiotics as Supplementation for Helicobacter pylori Antibiotic Therapy. Int. J. Mol. Sci. 2020, 21, 1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Yu, Z.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Surface Components and Metabolites of Probiotics for Regulation of Intestinal Epithelial Barrier. Microb. Cell Factories 2020, 19, 23. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhao, H.; Liu, L.; Wu, X.; Gao, Q.; Zhao, Y. Effects of Lactobacillus on the Inhibition of Helicobacter pylori Growth. Biotechnol. Biotechnol. Equip. 2018, 32, 1533–1540. [Google Scholar] [CrossRef] [Green Version]
- Lesbros-Pantoflickova, D.; Corthésy-Theulaz, I.; Blum, A.L. Helicobacter Pylori and Probiotics. J. Nutr. 2007, 137 (Suppl. 2), 812S–818S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, P.-X.; Fang, H.-Y.; Yang, H.-B.; Tien, N.-Y.; Wang, M.-C.; Wu, J.-J. Lactobacillus Pentosus Strain LPS16 Produces Lactic Acid, Inhibiting Multidrug-Resistant Helicobacter pylori. J. Microbiol. Immunol. Infect. 2016, 49, 168–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batdorj, B.; Trinetta, V.; Dalgalarrondo, M.; Prévost, H.; Dousset, X.; Ivanova, I.; Haertlé, T.; Chobert, J.-M. Isolation, Taxonomic Identification and Hydrogen Peroxide Production by Lactobacillus Delbrueckii Subsp. Lactis T31, Isolated from Mongolian Yoghurt: Inhibitory Activity on Food-Borne Pathogens. J. Appl. Microbiol. 2007, 103, 584–593. [Google Scholar] [CrossRef]
- Boyanova, L.; Gergova, G.; Markovska, R.; Yordanov, D.; Mitov, I. Bacteriocin-like Inhibitory Activities of Seven Lactobacillus Delbrueckii Subsp. Bulgaricus Strains against Antibiotic Susceptible and Resistant Helicobacter pylori Strains. Lett. Appl. Microbiol. 2017, 65, 469–474. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Study Subjects | Dominant Native Microbiota | Treatment | Microbial Alterations | Reference |
|---|---|---|---|---|
| H. pylori (HP) dyspeptic patients (Culture-dependent methodology) | ↑S. salivarius, S. mitior, S. sanguis, Micrococcus, Peptostreptococcus, Lactobacillus, Bifidobacterium, Fusobacterium, Veillonella | Omeprazole (20 mg) Amoxicillin (1 g) Metronidazole (400 mg) | ↑S. intermedius, Neisseria | [67] |
| ↓Staphylococcus, S. intermedius, Haemophilus, Neisseria, Enterobacteriaceae, Prevotella, Bacteroides | ↓S. salivarius, S. mitior, S. sanguis, Staphylococcus, Micrococcus, Peptostreptococcus, Lactobacillus, Bifidobacterium, Veillonella, Prevotella, Bacteroides, Fusobacterium | |||
| ↑S. salivarius, S. mitior, Enterobacteriaceae, Peptostreptococcus, Lactobacillus, Bifidobacterium, Veillonella, Prevotella, Bacteroides, Fusobacterium | Omeprazole (20 mg) Clarithromycin (250 mg) Metronidazole (400 mg) | ↑Staphylococcus, Micrococcus, Enterobacteriaceae, Veillonella | ||
| ↓S. intermedius, S. sanguis, Staphylococcus, Micrococcus, Haemophilus, Neisseria | ↓S. salivarus, S. mitior, Peptostreptococcus, Lactobacillus, Bifidobacterium, Prevotella, Bacteroides | |||
| HP peptic ulcer (Real-Time PCR) | ↑Prevotella, C. leptum ↓Enterobacteria | Rabeprazole (20 mg) Colloidal Bismuth pectin Clarithromycin (500 mg) Amoxicillin (1 g) | ↑Lactobacillus, C. leptum, Enterobacteria ↓C. coccoides | [68] |
| HP chronic gastritis | ↑H. pylori (83.70%), Eubacterium | Esomeprazole (20 mg) Amoxicillin (1 g) Clarithromycin (500 mg) | ↓H. pylori (6.88%) | [69] |
| HP intestinal metaplasia | ↑Proteobacteria no-HP (4.55%) | ↑Bacteroidetes, Fusobacteria, Actinobacteria | ||
| HP-negative patients | Haemophilus, Serratia, Neisseria, Stenotrophomonas | ↑Proteobacteria no-HP (51.70%) | ||
| HP gastric cancer (NGS) | ↑Proteobacteria Flavobacterium, Klebsiella, Serratia, Stenotrophomonas, Achromobacter, Pseudomonas | Flavobacterium, Neisseria, Serratia, Fusobacterium | ||
| HP-positive and negative patients (NGS) | Bacteroidetes:Firmicutes (0.94:0.84) Bifidobacterium, Lactobacillus, C. butyricum, Faecalibacterium prausnitzii, Akkermansia municiphila | Pantoprazole (40 mg) Amoxicillin (1 g) Furazolidone (100 mg) Colloidal Bismuth pectin (400 mg) | ↑Proteobacteria, Cyanobacteria ↓Firmicutes, Bacteroidetes, Verrucomicrobia, Lentisphaerae ↓Ruminococcaceae, Lachonspiracea | [48] |
| HP-negative patients (NGS) | Nitrospirae | ↑Enterobacteriaceae, Leuconostococaceae ↓Rikenellaceae, Christensenellaceae, Peptococcaceae, Clostridiales Family XI, Victivallaceae | ||
| HP-positive patients with different gastric pathologies (NGS) | ↑Eubacterium, Bacteroides, Prevotella | Amoxicillin (1 g) Clarithromycin (500 mg) Bismuth Subsalicylate (240 mg) Esomeprazole/Panteprazole (20 mg) | ↑Betaproteobacteria, Gammaproteobacteria Bacteroides, E. faecium ↑Enterobacteriaceae, Siphoviriadae ↓Actinobacteria, Verrucomicrobia, Synergistia ↓B. adolescentis Bifidobacteraceae, Coriobacteriaceae, Eubacteriaceae, Lachnospiraceae, Ruminococcaceae | [70] |
| HP-positive patients associated to: | Omeprazole (20 mg) Amoxicillin (1 g) Clarithromycin (500 mg) | [66] | ||
| (1) Atrophy | ↑Moraxella, Pasteurella, Bulleidia, Agrobacterium | ↑Pseudomonadaceae, Oxalobacteraceae, Microbacteriaceae, Enterobacteriaceae, Lachnospiraceae, Vibronaceae, Halomonadaceae ↑Acinetobacter, Ralstonia, Actinobacillus, Erwinia, Granulicatella, Streptococcus, Rothia, Leptotrichia ↓Helicobacter, Sphingomonas, Roseburia, Haemophilus influenzae, Actinobacillus parahaemolyticus, Neisseria subflava | ||
| (2) Intestinal metaplasia at progression | ↑Pseudomonas, Peptostreptococcus, Parvimonas, Halomonas | ↑Peptostreptococcus | ||
| (3) Intestinal metaplasia at regression (NGS) | ↑Lachnospira, Kaistobacter, Campylobacter, Devosia, Sphingobium | ↑Peptostreptococcus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomez-Ramirez, U.; Valencia-Mayoral, P.; Mendoza-Elizalde, S.; Murillo-Eliosa, J.R.; Solórzano Santos, F.; Contreras-Rodríguez, A.; Zúñiga, G.; Aguilar-Rodea, P.; Jiménez-Rojas, V.L.; Vigueras Galindo, J.C.; et al. Role of Helicobacter pylori and Other Environmental Factors in the Development of Gastric Dysbiosis. Pathogens 2021, 10, 1203. https://doi.org/10.3390/pathogens10091203
Gomez-Ramirez U, Valencia-Mayoral P, Mendoza-Elizalde S, Murillo-Eliosa JR, Solórzano Santos F, Contreras-Rodríguez A, Zúñiga G, Aguilar-Rodea P, Jiménez-Rojas VL, Vigueras Galindo JC, et al. Role of Helicobacter pylori and Other Environmental Factors in the Development of Gastric Dysbiosis. Pathogens. 2021; 10(9):1203. https://doi.org/10.3390/pathogens10091203
Chicago/Turabian StyleGomez-Ramirez, Uriel, Pedro Valencia-Mayoral, Sandra Mendoza-Elizalde, Juan Rafael Murillo-Eliosa, Fortino Solórzano Santos, Araceli Contreras-Rodríguez, Gerardo Zúñiga, Pamela Aguilar-Rodea, Verónica Leticia Jiménez-Rojas, Juan Carlos Vigueras Galindo, and et al. 2021. "Role of Helicobacter pylori and Other Environmental Factors in the Development of Gastric Dysbiosis" Pathogens 10, no. 9: 1203. https://doi.org/10.3390/pathogens10091203
APA StyleGomez-Ramirez, U., Valencia-Mayoral, P., Mendoza-Elizalde, S., Murillo-Eliosa, J. R., Solórzano Santos, F., Contreras-Rodríguez, A., Zúñiga, G., Aguilar-Rodea, P., Jiménez-Rojas, V. L., Vigueras Galindo, J. C., Salazar-García, M., & Velázquez-Guadarrama, N. (2021). Role of Helicobacter pylori and Other Environmental Factors in the Development of Gastric Dysbiosis. Pathogens, 10(9), 1203. https://doi.org/10.3390/pathogens10091203