
First Characterisation of the Phoma Species Complex on Maize Leaves in Central Europe


Abstract
:1. Introduction
2. Results
2.1. Collecting Fungal Isolates from Symptomatic Leaves
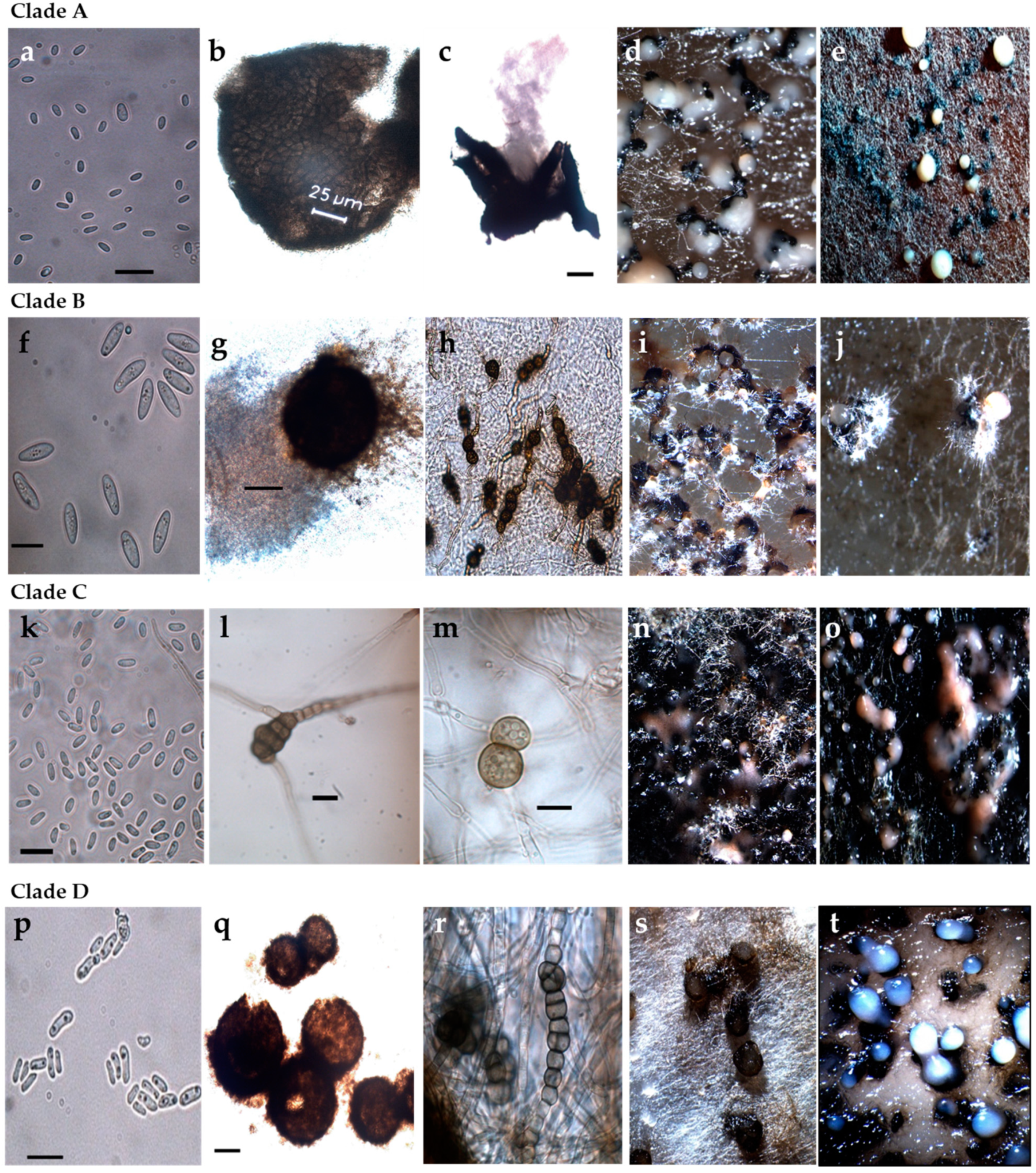
2.2. Morphology and Cultural Characteristics Related to Phylogenetic Clades
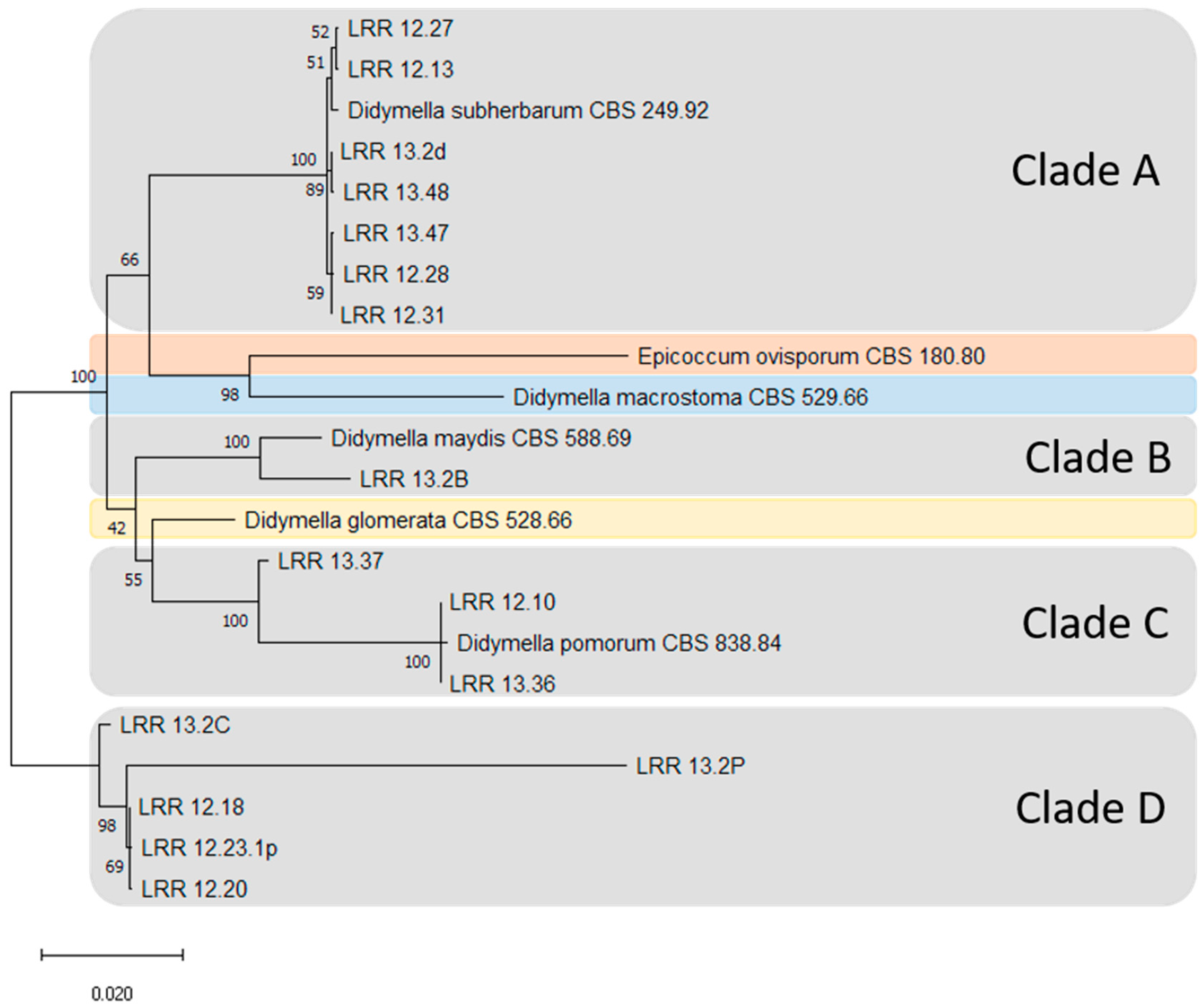
2.3. Phylogenetic Analysis
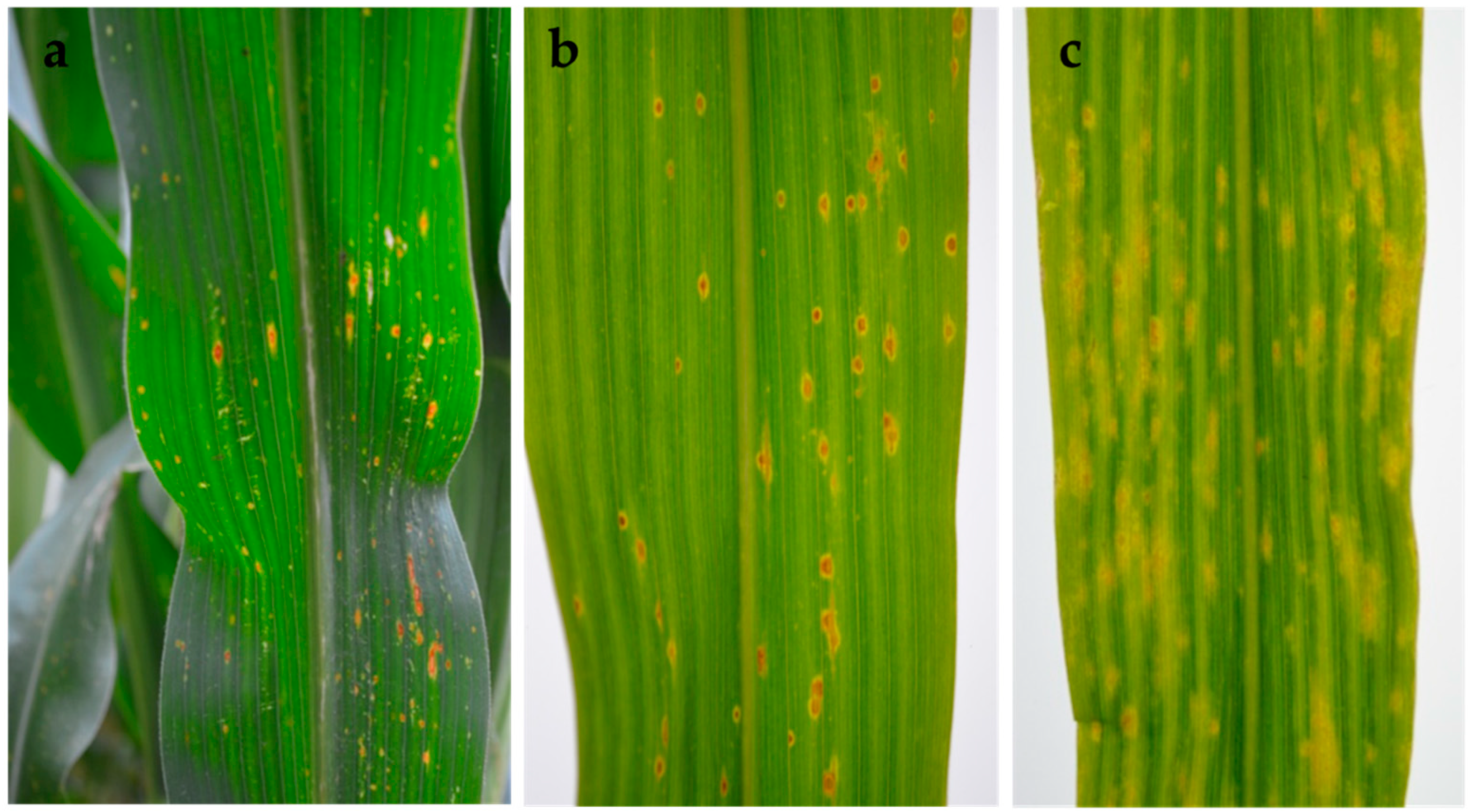
2.4. Pathogenicity Tests
3. Discussion
4. Materials and Methods
4.1. Isolation from Leaves and Culturing of Phoma-like Species
4.2. Morphological and Cultural Characterisation of Selected Phoma-Like Strains
4.3. Growth of Fungal Isolates for DNA Extraction
4.4. DNA Extraction
4.5. PCR Amplification of Multiple Loci
4.6. Cloning and Sequencing of PCR Amplicons
4.7. Sequence Analysis
4.8. Pathogenicity Tests
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abbate, L.; Mercati, F.; Del Bosco, S.F. An overview on citrus mal secco disease: Approaches and strategies to select tolerant genotypes in C. limon. Crop. Breed. Genet. Genom. 2019, 1, e190018. [Google Scholar]
- Harveson, R.; Mathew, F.; Gulya, T.; Markell, S.; Block, C.; Thompson, S. Sunflower stalk diseases initiated through leaf infections. Plant Health Prog. 2018, 19, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Mirleau-Thebaud, V.; Scheiner, J.; Daydé, J. Influence of soil tillage and Phoma macdonaldii on sunflower (Helianthus annuus L.) yield and oil quality. Phyton Int. J. Exp. Bot. 2011, 80, 203–210. [Google Scholar]
- Zhang, X.; Fernando, W.G.D. Insights into fighting against blackleg disease of Brassica napus in Canada. Crop. Pasture Sci. 2018, 69, 40–47. [Google Scholar] [CrossRef]
- Fitt, B.D.; Brun, H.; Barbetti, M.J.; Rimmer, S.R. World-wide importance of phoma stem canker (Leptosphaeria maculans and L. biglobosa) on oilseed rape (Brassica napus). Eur. J. Plant Pathol. 2006, 114, 3–15. [Google Scholar] [CrossRef]
- Van de Wouw, A.P.; Howlett, B.J. Advances in understanding the Leptosphaeria maculans—Brassica pathosystem and their impact on disease management. Can. J. Plant Pathol. 2020, 42, 149–163. [Google Scholar] [CrossRef]
- Yuan, G.Q.; Liao, T.; Tan, H.W.; Li, Q.Q.; Lin, W. First report of leaf spot caused by Phoma sorghina on tobacco in China. Plant Dis. 2016, 100, 1790. [Google Scholar] [CrossRef]
- Navi, S.S.; Bandyopadhyay, R.; Reddy, R.K.; Thakur, R.P.; Yang, X.B. Effects of wetness duration and grain development stages on sorghum grain mold infection. Plant Dis. 2005, 89, 872–878. [Google Scholar] [CrossRef]
- Oliveira, R.C.; Goncalves, S.S.; Oliveira, M.S.; Dilkin, P.; Mallmann, C.A.; Freitas, R.S. Natural occurrence of tenuazonic acid and Phoma sorghina in Brazilian sorghum grains at different maturity stages. Food Chem. 2017, 230, 491–496. [Google Scholar] [CrossRef]
- Perello, A.E.; Moreno, M.V. First report of Phoma sorghina (Sacc.) Boerema Dorenbosch & van Kest on wheat leaves (Triticum aestivum L.) in Argentina. Mycopathologia 2005, 159, 75–78. [Google Scholar]
- Larsen, J.E.; Hollingsworth, C.R.; Flor, J.; Dornbusch, M.R.; Simpson, N.L.; Samac, D.A. Distribution of Phoma sclerotioides on alfalfa and winter wheat crops in the North Central United States. Plant Dis. 2007, 91, 551–558. [Google Scholar] [CrossRef] [Green Version]
- Silva-Lobo, V.L.; de Aguiar, J.T.; de Côrtes, M.V.C.B.; de Filippi, M.C.C.; Prabhu, A.S. Critérios para avaliação da resistência à mancha parda e relação entre a mancha parda na folha bandeira e a mancha de grãos em genótipos de arroz. In Boletim de Pesquisa e Desenvolvimento/Embrapa Arroz e Feijão; Empresa Brasileira de Pesquisa Agropecuária, Embrapa Arroz e Feijão: Santo Antônio de Goiás, Brazil, 2014. [Google Scholar]
- Lin, Z.Y.; Que, Y.X.; Deng, Z.H.; Xu, S.Q.; Rao, G.P.; Zhang, M.Q. First report of Phoma sp causing twisting and curling of crown leaves of sugarcane in mainland of China. Plant Dis. 2014, 98, 850. [Google Scholar] [CrossRef]
- Lorenzetti, E.R.; Pozza, E.A.; Souza, P.E.D.; Santos, L.A.; Alves, E.; Silva, A.C.D. Effect of temperature and leaf wetness on Phoma tarda and phoma leaf spot in coffee seedlings. Coffee Sci. 2015, 10, 1–9. [Google Scholar]
- Arny, D.C.; Smalley, E.B.; Ullstrup, A.J.; Worf, G.L.; Ahrens, R.W. Eyespot of maize, a disease new to North-America. Phytopathology 1971, 61, 54–57. [Google Scholar] [CrossRef]
- Do Amaral, A.L.; De Carli, M.L.; Barbosa Neto, J.F.; Dal Soglio, F.K. Phoma sorghina, a new pathogen associated with phaeosphaeria leaf spot on maize in Brazil. Plant Pathol. 2004, 53, 259. [Google Scholar] [CrossRef]
- Koenning, S.R.; Frye, J.W.; Pataky, J.K.; Gibbs, M.; Cotton, D. First report of Phoma terrestris causing red root rot on sweet corn (Zea mays) in North Carolina. Plant Dis. 2007, 91, 1054. [Google Scholar] [CrossRef] [PubMed]
- Carli, M. Etiological and Epidemiological Aspects of the Maize White Leaf Spot Complex; Universidade Federal do Rio Grande do Sul: Porto Alegre, Brazil, 2008. [Google Scholar]
- Su, G.; Suh, S.O.; Schneider, R.W.; Russin, J.S. Host specialization in the charcoal rot fungus, Macrophomina phaseolina. Phytopathology 2001, 91, 120–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawlal, P.; Barros, E.; Marais, G.J. Evaluation of maize cultivars for their susceptibility towards mycotoxigenic fungi under storage conditions. J. Stored Prod. Res. 2012, 48, 114–119. [Google Scholar] [CrossRef]
- Gonçalves, R.M.; Figueiredo, J.E.F.; Pedro, E.S.; Meirelles, W.F.; Leite, R.P.; Sauer, A.V.; Paccola-Meirelles, L.D. Etiology of Phaeosphaeria leaf spot disease of maize. J. Plant Pathol. 2013, 95, 559–569. [Google Scholar]
- Frezzi, M. Mancha ocular del maíz (Kabatiella zeae) y tizón amarillo de la hoja del maíz (Phyllosticta zeae) en la Argentina. Rev. Investig. Agropecu 1972, 9, 101–109. [Google Scholar]
- Jimenez Diaz, R.M.; Boothroyd, C.W. Suscept-pathogen relationship in maize affected by yellow leaf-blight. Phytopathology 1976, 66, 1169–1173. [Google Scholar] [CrossRef]
- McFeeley, J.C. Comparison of isolates causing yellow leaf-blight of corn in Ohio. Plant Dis. Report 1971, 55, 1064. [Google Scholar]
- Scheifele, G.; Nelson, R. The occurrence of Phyllosticta leaf spot of corn in Pennsylvania. Plant Dis. Report 1969, 53, 186–189. [Google Scholar]
- Gates, L.F.; Mortimore, C.G. Three diseases of corn (Zea mays), new to Ontario: Crazytop, a Phyllosticta leaf spot and eyespot. Can. Plant Dis. Surv. 1969, 49, 128–131. [Google Scholar]
- Carson, M.L. Yield loss potential of phaeosphaeria leaf spot of maize caused by Phaeosphaeria maydis in the United States. Plant Dis. 2005, 89, 986–988. [Google Scholar] [CrossRef] [PubMed]
- Ferraguti, F.J.; Miguez, L.; Magnone, G.; Couretot, L.A. Aparición de Mancha Blanca (Phoma sp.) en Ensayos de Maíz Tardío de la Red EEA INTA Oliveros. 2017. Available online: http://inta.gob.ar/sites/default/files/inta-aparicion-mancha-blanca-maiz.pdf (accessed on 17 September 2020).
- Aveskamp, M.M.; De Gruyter, J.; Crous, P.W. Biology and recent developments in the systematics of Phoma, a complex genus of major quarantine significance. Fungal Divers. 2008, 31, 1–18. [Google Scholar]
- Aveskamp, M.M.; De Gruyter, J.; Woudenberg, J.H.C.; Verkley, G.J.M.; Crous, P.W. Highlights of the Didymellaceae: A polyphasic approach to characterise Phoma and related pleosporalean genera. Stud. Mycol. 2010, 65, 1–60. [Google Scholar] [CrossRef]
- Boerema, G.H.; Gruyter, J.; Noordeloos, M.E.; Hamers, M.E. Phoma Identification Manual. Differentiation of Specific and Intra-Specific Taxa in Culture; CABI Publishing: Wallingford, UK, 2004. [Google Scholar]
- Chen, Q.; Jiang, J.R.; Zhang, G.Z.; Cai, L.; Crous, P.W. Resolving the Phoma enigma. Stud. Mycol. 2015, 82, 137–217. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Hou, L.W.; Duan, W.J.; Crous, P.W.; Cai, L. Didymellaceae revisited. Stud. Mycol. 2017, 87, 105–159. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Castor, L.L.; Ayers, J.E.; Nelson, R.R. Controlled-environment studies of epidemiology of yellow leaf-blight of corn. Phytopathology 1977, 67, 85–90. [Google Scholar] [CrossRef]
- Mukunya, D.; Boothroyd, C. Mycosphaerella zeae-maydis sp. n., the Sexual Stage of Phyllosticta maydis. Phytopathology 1973, 63, 529–532. [Google Scholar] [CrossRef]
- Yun, S.H.; Yoder, O.C.; Turgeon, B.G. Structure and function of the mating-type locus in the homothallic ascomycete, Didymella zeae-maydis. J. Microbiol. 2013, 51, 814–820. [Google Scholar] [CrossRef] [PubMed]
- Stout, G. New fungi found on the Indian corn plant in Illinois. Mycologia 1930, 22, 271–287. [Google Scholar] [CrossRef]
- Bai, J. Sphaeropsidales, Phoma, Phyllosticta. In Flora Fungorum Sinicorum; Science Press: Beijing, China, 2003; Volume 15. [Google Scholar]
- Zhang, K.; Shivas, R.G.; Cai, L. Synopsis of Phyllosticta in China. Mycology 2015, 6, 50–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koons, C.; Ayers, J.; Nelson, R. Studies on yellow leaf blight of corn caused by Phyllosticta zeae. Phytopathology 1970, 60, 575–576. [Google Scholar]
- Cervelatti, E.; Paíva, E.; Meirelles, W.F.; Casela, C.; Fernandes, F.T.; Teixeira, F.F.; Paccola-Meirelles, L. Characterization of fungal isolates from pycnidia and pseudothecia from lesions of Phaeosphaeria leaf spot in maize. Rev. Bras. Milho Sorgo 2002, 1, 30–37. [Google Scholar] [CrossRef] [Green Version]
- De Gruyter, J.; Noordeloos, M.E.; Boerema, G.H. Contributions towards a monograph of Phoma (Coelomycetes). 1.2 Section Phoma-Additional taxa with very small conidia and taxa with conidia up to 7 µm long. Persoonia 1993, 15, 369–400. [Google Scholar]
- Demİrcİ, E.R.K.O.L.; Kordalİ, Ș. Fungi isolated from corn kernels in the Eastern Black Sea Region. J. Turk. Phytopathol. 2000, 29, 79–84. [Google Scholar]
- Payak, M.; Sharma, R.; Rao, S. Aflatoxin in India II. In Aflatoxin in Maize. A Proceedings of the Workshop; CIMMYT: Mexico D.F., Mexico, 1987. [Google Scholar]
- Sørensen, J.L.; Aveskamp, M.M.; Thrane, U.; Andersen, B. Chemical characterization of Phoma pomorum isolated from Danish maize. Int. J. Food Microbiol. 2010, 136, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Stover, R.; Waite, B. An improved method of isolating Fusarium spp. from plant tissues. Phytopathology 1953, 43, 700–701. [Google Scholar]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Xin, Z.; Chen, J. A high throughput DNA extraction method with high yield and quality. Plant Methods 2012, 8, 26. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Rehner, S.; Samuels, G. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, I.; Kohn, L. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Aveskamp, M.M.; De Gruyter, J.; Spiers, A.G.; Crous, P.W. Multiple Didymella teleomorphs are linked to the Phoma clematidina morphotype. Persoonia 2009, 22, 56–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, M.R.; Sambrook, J. Molecular Cloning: Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2012. [Google Scholar]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IPP# 1 | Description | Strain Designation 2 | Location | Region | Isolated From 3 |
|---|---|---|---|---|---|
| 1562 | Phoma sp. | LRR 12.18 | Schönering | Northern Austria | 3 |
| 1563 | Phoma sp. | LRR 13.2B | Ostenfeld | Northern Germany | 2 |
| 1564 | Phoma sp. | LRR 13.2C | Ostenfeld | Northern Germany | 2 |
| 1566 | Phoma sp. | LRR 12.27 | Bad Oldesloe | Northern Germany | 3 |
| 1567 | Phoma sp. | LRR 12.13 | Stapel | Northern Germany | 3 |
| 1568 | Phoma sp. | LRR 12.10 | Braunau | Northern Austria | 4 |
| 1569 | Phoma sp. | LRR 13.2d | Ostenfeld | Northern Germany | 1 |
| 1570 | Phoma sp. | LRR 13.48 | Kassel | Central Germany | 2 |
| 1571 | Phoma sp. | LRR 13.47 | Kassel | Central Germany | 2 |
| 1572 | Phoma sp. | LRR 12.28 | Groningen | The Netherlands | 3 |
| 1573 | Phoma sp. | LRR 13.36 | Mittich | Southern Germany | 5 |
| 1574 | Phoma sp. | LRR 13.37 | Hartkirchen | Southern Germany | 5 |
| 1575 | Phoma sp. | LRR 12.23.1p | Rade | Northern Germany | 2 |
| 1576 | Phoma sp. | LRR 13.2P | Ostenfeld | Northern Germany | 6 |
| 1577 | Phoma sp. | LRR 12.20 | Nordholz | Northern Germany | 2 |
| 1585 | Phoma sp. | LRR 12.31 | Giekau | Northern Germany | 3 |
| Clade | A | B | C | D * |
|---|---|---|---|---|
| Pathogenic on maize | no | no | no | yes |
| Colony type | ||||
| White-grey-green | + | ± | ||
| Red/vinaceous/brown | ++ | |||
| Brown | + | + | ± | |
| Growth | ||||
| Fast (45–75 mm) | ++ | + | ||
| Moderate-slow (24–43 mm) | + | + | ++ | |
| Pycnidia | ||||
| Glabrous | + | + | + | |
| Pilose | ++ | |||
| Form | Globose/subglobose/irregular/flask-shaped | Globose/subglobose/irregular | Globose/subglobose | Globose/subglobose/irregular |
| Solitary | + | + | + | + |
| Confluent | + | + | ± | |
| Conspicuous ostiole/necks | ++/+ | |||
| Size (l × w) (µm) | 70–300 × 75–250 | 200–500 × 100–400 | 50–200 × 45–200 ** | 50–350 × 50–300 |
| Matrix color | Rosy-buff/white | Hyaline-white | Hyaline/pinkish | Hyaline/white |
| Chlamydospores | ||||
| Unicellular | ± | + | ± | ± |
| Multicellular | + | ± | ± | |
| Conidia | ||||
| Common size | Small | Large | Small | Small |
| Size (l × w) | 2.5–6.3 × 0.9–2.4 | 9.9–11.9 × 3.3–4.3 | 3.8–6.5 × 1.6–2.3 | 4.1–7.9 × 1.5–3.2 |
| Size (average) | 3.7 × 1.7 | 10.9 × 3.9 | 4.9 × 2.3 | 4.7 × 2.4 |
| Aseptate | + | + | + | + |
| Extrusion in cirri | + | |||
| Gutules | 0–2 | 1–5 | 0–5 | 1–5 |
| Polar | + | + | + | + |
| Apolar | + | ± |
| IPP# | Description | Strain Code | Originator | Reference |
|---|---|---|---|---|
| 1402 | Didymella maydis | CBS 588.69 | WFBI * | [32] |
| 1403 | Epicoccum ovisporum | CBS 180.80 | WFBI | [48] |
| 1404 | D. macrostoma | CBS 529.66 | WFBI | [32] |
| 1405 | D. pomorum | CBS 838.84 | WFBI | [32] |
| 1406 | D. glomerata | CBS 528.66 | WFBI | [32] |
| 1462 | D. subherbarum | CBS 249.92 | WFBI | [32] |
| Primer Name | Primer Sequence | Reference | Amplicon Sizes (bp) | PCR Profile |
|---|---|---|---|---|
| Partial 18S ribosomal RNA gene (SSU) | 1133 | 98 °C, 10 s; 60 °C, 30 s; 72 °C, 90 s; (35×) | ||
| NS1-fw | GTAGTCATATGCTTGTCTC | [50] | ||
| NS4-rev | CTTCCGTCAATTCCTTTAAG | [50] | ||
| Partial 28S ribosomal RNA gene (LSU) | 1421–1422 | 98 °C, 10 s; 60 °C, 30 s; 72 °C, 90 s; (35×) | ||
| LR0R-fw | GTACCCGCTGAACTTAAGC | [51] | ||
| LR7-rev | TACTACCACCAAGATCT | [52] | ||
| 5.8S nrRNA gene with the two flanking internal transcribed spacers (ITS) | 558–565 | 98 °C, 10 s; 59 °C, 15 s; 72 °C, 15 s; (35×) | ||
| ITS-5-fw | GGAAGTAAAAGTCGTAACAAGG | [50] | ||
| ITS-4-rev | TCCTCCGCTTATTGATATGC | [50] | ||
| Partial actin gene (ACT) | 263–275 | 98 °C, 10 s; 66 °C, 30 s, 72 °C 15 s; (30×) | ||
| ACT-512F-fw | ATGTGCAAGGCCGGTTTCGC | [53] | ||
| ACT-783R-rev | TACGAGTCCTTCTGGCCCAT | [53] | ||
| Partial beta-tubulin gene (Tub2) | 376–382 | 98 °C, 10 s; 63 °C, 30 s; 72 °C, 15 s; (30×) | ||
| TUB2Fd-fw | GTBCACCTYCARACCGGYCARTG | [54] | ||
| TUB4Rd-rev | CCRGAYTGRCCRAARACRAAGTTGTC | [54] | ||
| Partial calmodulin gene (CAL) | 531–544 | 98 °C, 10 s; 63 °C, 30 s; 72 °C, 15 s; (35×) | ||
| CAL-228F-fw-BK | GAGTTCAAGGAGGCATTCTCTC GAGTTCAAGGAGGCCTTCTCCC | This study, modified from [53], original sequence in italics | ||
| CAL-737R rev-BK | CATCTTGCGGGCCATCATAG CATCTTTCTGGCCATCATGG | This study, modified from [53], original sequence in italics | ||
| IPP No. | Description | Strain Designation | Conidia/mL |
|---|---|---|---|
| 1562 | Phoma sp. | LRR 12.18 | 5 × 106–1 × 107 |
| 1563 | Phoma sp. | LRR 13.2B | 1.1 × 106–4.5 × 106 |
| 1564 | Phoma sp. | LRR 13.2C | 1 × 107 |
| 1577 | Phoma sp. | LRR 12.20 | 1 × 107 |
| 1575 | Phoma sp. | LRR 12.23.1p | 2 × 106–4 × 106 |
| 1576 | Phoma sp. | LRR 13.2P | 1 × 107 |
| 1566 | Phoma sp. | LRR 12.27 | 1 × 107 |
| 1567 | Phoma sp. | LRR 12.13 | 1 × 107 |
| 1568 | Phoma sp. | LRR 12.10 | 1 × 107 |
| 1569 | Phoma sp. | LRR 13.2d | 1 × 107 |
| 1570 | Phoma sp. | LRR 13.48 | 1 × 107 |
| 1571 | Phoma sp. | LRR 13.47 | 1 × 107 |
| 1572 | Phoma sp. | LRR 12.28 | 1 × 107 |
| 1573 | Phoma sp. | LRR 13.36 | 1 × 107 |
| 1574 | Phoma sp. | LRR 13.37 | 1 × 107 |
| 1585 | Phoma sp. | LRR 12.31 | 1 × 107 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos Romero, L.; Tacke, D.; Koopmann, B.; von Tiedemann, A. First Characterisation of the Phoma Species Complex on Maize Leaves in Central Europe. Pathogens 2021, 10, 1216. https://doi.org/10.3390/pathogens10091216
Ramos Romero L, Tacke D, Koopmann B, von Tiedemann A. First Characterisation of the Phoma Species Complex on Maize Leaves in Central Europe. Pathogens. 2021; 10(9):1216. https://doi.org/10.3390/pathogens10091216
Chicago/Turabian StyleRamos Romero, Lucia, Dagmar Tacke, Birger Koopmann, and Andreas von Tiedemann. 2021. "First Characterisation of the Phoma Species Complex on Maize Leaves in Central Europe" Pathogens 10, no. 9: 1216. https://doi.org/10.3390/pathogens10091216
APA StyleRamos Romero, L., Tacke, D., Koopmann, B., & von Tiedemann, A. (2021). First Characterisation of the Phoma Species Complex on Maize Leaves in Central Europe. Pathogens, 10(9), 1216. https://doi.org/10.3390/pathogens10091216