
Amphotericin B Polymer Nanoparticles Show Efficacy against Candida Species Biofilms

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. AmB Formulation for Biofilm Studies
2.3. AmB Eye Drops
2.4. AmB Activity in Biofilms
2.5. AmB Activity in Planktonic C. albicans Cell Suspensions
2.6. Texas Red (TR) Labelling of MET Polymer
2.7. Confocal Laser Scanning Microscopy (CLSM)
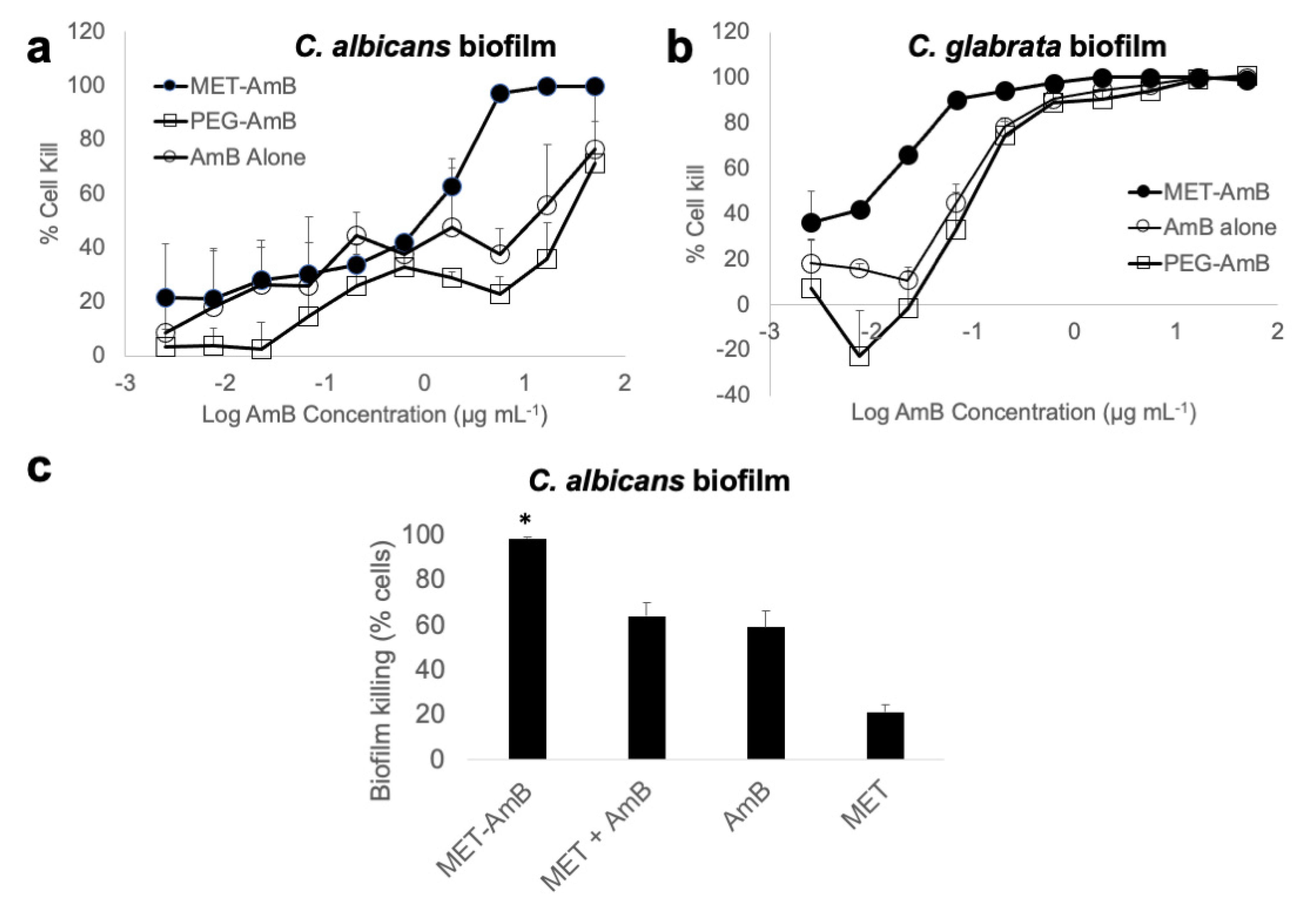
3. Results
3.1. Formulations
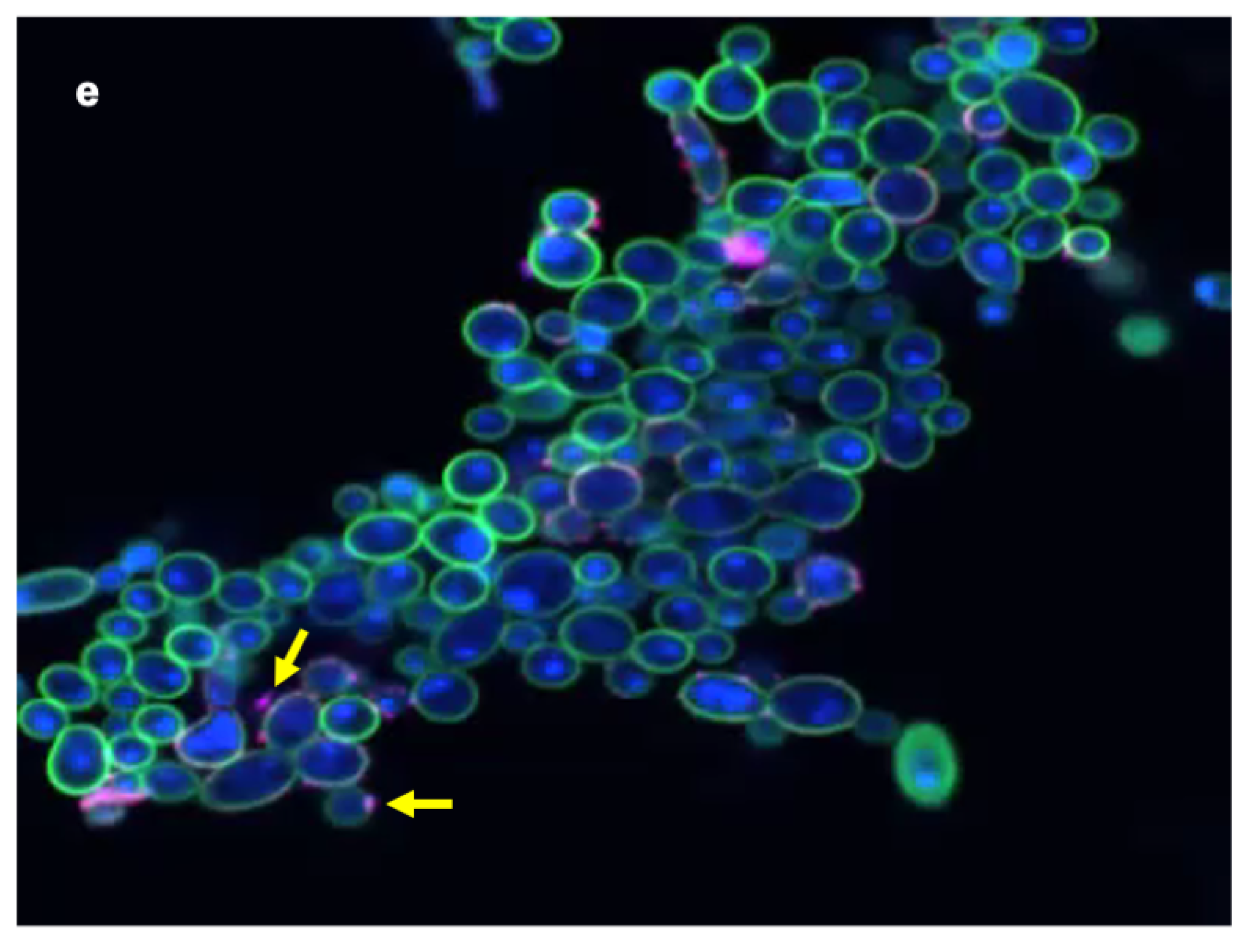
3.2. Confocal Laser Scanning Microscopy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bell, M. Biofilms: A Clinical Perspective. Curr. Infect. Dis. Rep. 2001, 3, 483–486. [Google Scholar] [CrossRef]
- Fanning, S.; Mitchell, A.P. Fungal biofilms. PLoS Pathog. 2012, 8, e1002585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalheiro, M.; Teixeira, M.C. Candida Biofilms: Threats, Challenges, and Promising Strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Nobile, C.J.; Johnson, A.D. Candida albicans Biofilms and Human Disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Xu, Y.C.; Hsueh, P.R. Epidemiology of candidemia and antifungal susceptibility in invasive Candida species in the Asia-Pacific region. Future Microbiol. 2016, 11, 1461–1477. [Google Scholar] [CrossRef] [PubMed]
- Pierce, C.G.; Vila, T.; Romo, J.A.; Montelongo-Jauregui, D.; Wall, G.; Ramasubramanian, A.; Lopez-Ribot, J.L. The Candida albicans Biofilm Matrix: Composition, Structure and Function. J. Fungi 2017, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Thein, Z.M.; Samaranayake, Y.H.; Samaranayake, L.P. In vitro biofilm formation of Candida albicans and non-albicans Candida species under dynamic and anaerobic conditions. Arch. Oral Biol. 2007, 52, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Lagree, K.; Desai, J.V.; Finkel, J.S.; Lanni, F. Microscopy of fungal biofilms. Curr. Opin. Microbiol. 2018, 43, 100–107. [Google Scholar] [CrossRef]
- Kucharikova, S.; Tournu, H.; Lagrou, K.; Van Dijck, P.; Bujdakova, H. Detailed comparison of Candida albicans and Candida glabrata biofilms under different conditions and their susceptibility to caspofungin and anidulafungin. J. Med. Microbiol. 2011, 60, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Rodrigues, C.F.; Araújo, D.; Rodrigues, M.E.; Henriques, M. Candida Species Biofilms’ Antifungal Resistance. J. Fungi 2017, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Elder, M.J.; Matheson, M.; Stapleton, F.; Dart, J.K.G. Biofilm Formation in Infectious Crystalline Keratopathy Due to Candida albicans. Cornea 1996, 15, 301–304. [Google Scholar] [CrossRef]
- Ranjith, K.; Kalyana Chakravarthy, S.; Adicherla, H.; Sharma, S.; Shivaji, S. Temporal Expression of Genes in Biofilm-Forming Ocular Candida albicans Isolated From Patients With Keratitis and Orbital Cellulitis. Investig. Ophthalmol. Vis. Sci. 2018, 59, 528–538. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Sun, X.; Wang, Z.; Zhang, Y.; Hou, W. Keratitis-Associated Fungi Form Biofilms with Reduced Antifungal Drug Susceptibility. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7774–7778. [Google Scholar] [CrossRef] [Green Version]
- Gallis, H.A.; Drew, R.H.; Pickard, W.W. Amphotericin B: 30 years of clinical experience. Rev. Infect. Dis. 1990, 12, 308–329. [Google Scholar] [CrossRef] [PubMed]
- Odds, F.C.; Brown, A.J.; Gow, N.A. Antifungal agents: Mechanisms of action. Trends Microbiol. 2003, 11, 272–279. [Google Scholar] [CrossRef]
- Rodrigues, C.; Henriques, M. Liposomal and Deoxycholate Amphotericin B Formulations: Effectiveness against Biofilm Infections of Candida spp. Pathogens 2017, 6, 62. [Google Scholar] [CrossRef] [Green Version]
- Hamill, R.J. Amphotericin B formulations: A comparative review of efficacy and toxicity. Drugs 2013, 73, 919–934. [Google Scholar] [CrossRef] [PubMed]
- Serrano, D.R.; Lalatsa, A.; Dea-Ayuela, M.A.; Bilbao-Ramos, P.E.; Garrett, N.L.; Moger, J.; Guarro, J.; Capilla, J.; Ballesteros, M.P.; Schatzlein, A.G.; et al. Oral particle uptake and organ targeting drives the activity of amphotericin B nanoparticles. Mol. Pharm. 2015, 12, 420–431. [Google Scholar] [CrossRef]
- Siew, A.; Le, H.; Thiovolet, M.; Gellert, P.; Schatzlein, A.; Uchegbu, I. Enhanced oral absorption of hydrophobic and hydrophilic drugs using quaternary ammonium palmitoyl glycol chitosan nanoparticles. Mol. Pharm. 2012, 9, 14–28. [Google Scholar] [CrossRef]
- Garrett, N.L.; Lalatsa, A.; Uchegbu, I.; Schatzlein, A.; Moger, J. Exploring uptake mechanisms of oral nanomedicines using multimodal nonlinear optical microscopy. J. Biophotonics 2012, 5, 458–468. [Google Scholar] [CrossRef]
- Badr, M.Y.; Abdulrahman, N.S.; Schatzlein, A.G.; Uchegbu, I.F. A polymeric aqueous tacrolimus formulation for topical ocular delivery. Int. J. Pharm. 2021, 599, 120364. [Google Scholar] [CrossRef]
- Uchegbu, I.F.; Breznikar, J.; Zaffalon, A.; Odunze, U.; Schatzlein, A.G. Polymeric Micelles for the Enhanced Deposition of Hydrophobic Drugs into Ocular Tissues, without Plasma Exposure. Pharmaceutics 2021, 13, 744. [Google Scholar] [CrossRef] [PubMed]
- Galdiero, E.; de Alteriis, E.; De Natale, A.; D’Alterio, A.; Siciliano, A.; Guida, M.; Lombardi, L.; Falanga, A.; Galdiero, S. Eradication of Candida albicans persister cell biofilm by the membranotropic peptide gH625. Sci. Rep. 2020, 10, 5780. [Google Scholar] [CrossRef] [Green Version]
- Tits, J.; Berman, J.; Cammue, B.P.A.; Thevissen, K. Combining Miconazole and Domiphen Bromide Results in Excess of Reactive Oxygen Species and Killing of Biofilm Cells. Front. Cell Dev. Biol. 2020, 8, 617214. [Google Scholar] [CrossRef] [PubMed]
- Tits, J.; Cammue, B.P.A.; Thevissen, K. Combination Therapy to Treat Fungal Biofilm-Based Infections. Int. J. Mol. Sci. 2020, 21, 8873. [Google Scholar] [CrossRef]
- Phillips, A.J. Treatment of non-albicans Candida vaginitis with amphotericin B vaginal suppositories. Am. J. Obs. Gynecol. 2005, 192, 2009–2012. [Google Scholar] [CrossRef] [PubMed]
- Harriott, M.M.; Lilly, E.A.; Rodriguez, T.E.; Fidel, P.L.; Noverr, M.C. Candida albicans forms biofilms on the vaginal mucosa. Microbiology (Reading) 2010, 156, 3635–3644. [Google Scholar] [CrossRef] [Green Version]
- Sherry, L.; Kean, R.; McKloud, E.; O’Donnell, L.E.; Metcalfe, R.; Jones, B.L.; Ramage, G. Biofilms Formed by Isolates from Recurrent Vulvovaginal Candidiasis Patients Are Heterogeneous and Insensitive to Fluconazole. Antimicrob. Agents Chemother. 2017, 61, e01065-17. [Google Scholar] [CrossRef] [Green Version]
- Pierce, C.G.; Uppuluri, P.; Tristan, A.R.; Wormley, F.L., Jr.; Mowat, E.; Ramage, G.; Lopez-Ribot, J.L. A simple and reproducible 96-well plate-based method for the formation of fungal biofilms and its application to antifungal susceptibility testing. Nat. Protoc. 2008, 3, 1494–1500. [Google Scholar] [CrossRef]
- Rodriguez-Tudela, J.L.; Donnelly, J.P.; Arendrup, M.C.; Arikan, S.; Barchiesi, F.; Bille, J.; Chryssanthou, E.; Cuenca-Estrella, M.; Dannaoui, E.; Fegele, W.; et al. Subcommittee on Antifungal Susceptibility Testing of the ESCMID European Committee for Antimicrobial Susceptibility Testing. EUCAST Technical Note on the method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for conidia-forming moulds. Clin. Microbiol. Infect. 2008, 14, 982–984. [Google Scholar] [CrossRef]
- Carney, L.G.; Hill, R.M. Human Tear pH: Diurnal Variations. Arch. Ophthalmol. 1976, 94, 821–824. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, R.A.; Teixeira, C.E.C.; Brilhante, R.S.N.; Castelo-Branco, D.S.C.M.; Paiva, M.A.N.; Giffoni Leite, J.J.; Lima, D.T.; Monteiro, A.J.; Sidrim, J.J.C.; Rocha, M.F.G. Minimum inhibitory concentrations of amphotericin B, azoles and caspofungin against Candida species are reduced by farnesol. Med. Mycol. 2013, 51, 53–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyrc, K.; Milewska, A.; Duran, E.B.; Botwina, P.; Dabrowska, A.; Jedrysik, M.; Benedyk, M.; Lopes, R.; Arenas-Pinto, A.; Badr, M.; et al. SARS-CoV-2 inhibition using a mucoadhesive, amphiphilic chitosan that may serve as an anti-viral nasal spray. Sci. Rep. 2021, 11, 20012. [Google Scholar] [CrossRef]
- Singh, R.; Kumari, A.; Kaur, K.; Sethi, P.; Chakrabarti, A. Relevance of antifungal penetration in biofilm-associated resistance of Candida albicans and non-albicans Candida species. J. Med. Microbiol. 2018, 67, 922–926. [Google Scholar] [CrossRef]
- Al-Fattani, M.A.; Douglas, L.J. Penetration of Candida Biofilms by Antifungal Agents. Antimicrob. Agents Chemother. 2004, 48, 3291–3297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vediyappan, G.; Rossignol, T.; d’Enfert, C. Interaction of Candida albicans biofilms with antifungals: Transcriptional response and binding of antifungals to beta-glucans. Antimicrob. Agents Chemother. 2010, 54, 2096–2111. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, C.F.; Silva, S.; Azeredo, J.; Henriques, M. Candida glabrata’s recurrent infections: Biofilm formation during Amphotericin B treatment. Lett. Appl. Microbiol. 2016, 63, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, K.F.; Zarnowski, R.; Sanchez, H.; Edward, J.A.; Reinicke, E.L.; Nett, J.E.; Mitchell, A.P.; Andes, D.R. Community participation in biofilm matrix assembly and function. Proc. Natl. Acad. Sci. USA 2015, 112, 4092–4097. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Time Point | Concentration (mg mL−1) | (%) Remaining Drug Content | z-Average (nm) | Polydispersity Index (PDI) | Zeta Potential (mV) | pH | Osmolarity (mOsm) | |
|---|---|---|---|---|---|---|---|---|
| Day 0 | 1.81 ± 0.06 | - | 59.7 ± 1.4 | 0.381 ± 0.050 | 28.0 ± 0.6 | 7.2 ± 0.1 | 355 ± 19 | |
| Week 1 | 5 °C | 1.90 ± 0.06 | 106.7 ± 4.2 | 77.9 ± 5.9 * | 0.252 ± 0.020 * | 27.4 ± 0.9 | 6.9 ± 0.0 * | 331 ± 3 |
| RT | 1.81 ± 0.07 | 101.4 ± 3.3 | 74.4 ± 5.2 * | 0.241 ± 0.019 * | 27.3 ± 0.6 | 6.8 ± 0.1 | 339 ± 7 | |
| 40 °C | 1.86 ± 0.04 | 104.1 ± 1.9 | 88.1 ± 2.2 * | 0.231 ± 0.018 * | 28.7 ± 1.0 | 6.8 ± 0.0 * | 330 ± 4 | |
| Week 2 | 5 °C | 1.84 ± 0.07 | 103.4 ± 3.7 | 79.6 ± 5.2 * | 0.245 ± 0.018 * | 26.0 ± 0.9 | 7.0 ± 0.0 * | 334 ± 4 |
| RT | 1.74 ± 0.03* | 97.8 ± 3.1 | 76.2 ± 4.4 * | 0.235 ± 0.012 * | 27.0 ± 0.8 | 7.1 ± 0.0 | 335 ± 5 | |
| 40 °C | 1.72 ± 0.03 | 96.5 ± 2.3 | 89.4 ± 2.8 * | 0.225 ± 0.010 * | 28.2 ± 3.4 | 7.0 ± 0.0 * | 336 ± 4 | |
| Week 4 | 5 °C | 1.91 ± 0.03 | 107.1 ± 1.9 | 85.9 ± 6.7 * | 0.234 ± 0.010 * | 28.3 ± 0.7 | 7.1 ± 0.0 | 338 ± 4 |
| RT | 1.89 ± 0.04 | 106.0 ± 1.0 | 80.2 ± 4.7 * | 0.225 ± 0.006 * | 26.7 ± 1.3 | 7.1 ± 0.0 | 334 ± 4 | |
| 40 °C | 1.81 ± 0.11 | 100.1 ± 4.2 | 89.5 ±1.1 * | 0.229 ± 0.008 * | 29.7 ± 1.0 | 6.9 ± 0.1 * | 336 ± 3 | |
| Formulation | MIC (μg mL−1) |
|---|---|
| MET-AmB | 0.125 |
| AmB | 0.25 |
| MET | >7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alakkad, A.; Stapleton, P.; Schlosser, C.; Murdan, S.; Odunze, U.; Schatzlein, A.; Uchegbu, I.F. Amphotericin B Polymer Nanoparticles Show Efficacy against Candida Species Biofilms. Pathogens 2022, 11, 73. https://doi.org/10.3390/pathogens11010073
Alakkad A, Stapleton P, Schlosser C, Murdan S, Odunze U, Schatzlein A, Uchegbu IF. Amphotericin B Polymer Nanoparticles Show Efficacy against Candida Species Biofilms. Pathogens. 2022; 11(1):73. https://doi.org/10.3390/pathogens11010073
Chicago/Turabian StyleAlakkad, Abdulghani, Paul Stapleton, Corinna Schlosser, Sudaxshina Murdan, Uchechukwu Odunze, Andreas Schatzlein, and Ijeoma F. Uchegbu. 2022. "Amphotericin B Polymer Nanoparticles Show Efficacy against Candida Species Biofilms" Pathogens 11, no. 1: 73. https://doi.org/10.3390/pathogens11010073
APA StyleAlakkad, A., Stapleton, P., Schlosser, C., Murdan, S., Odunze, U., Schatzlein, A., & Uchegbu, I. F. (2022). Amphotericin B Polymer Nanoparticles Show Efficacy against Candida Species Biofilms. Pathogens, 11(1), 73. https://doi.org/10.3390/pathogens11010073