

The Enterotoxin Production and Antimicrobial Resistance of Campylobacter Strains Originating from Slaughter Animals
 , , , and
, , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Bacterial Strains
2.2. Enterotoxin Production Assay
2.2.1. Preparation of Culture Filtrates
2.2.2. The Influence of Enterotoxin on CHO Cells
2.2.3. Suckling Mouse Assay
2.3. Antimicrobial Resistance
2.4. Statistical Analysis
3. Results
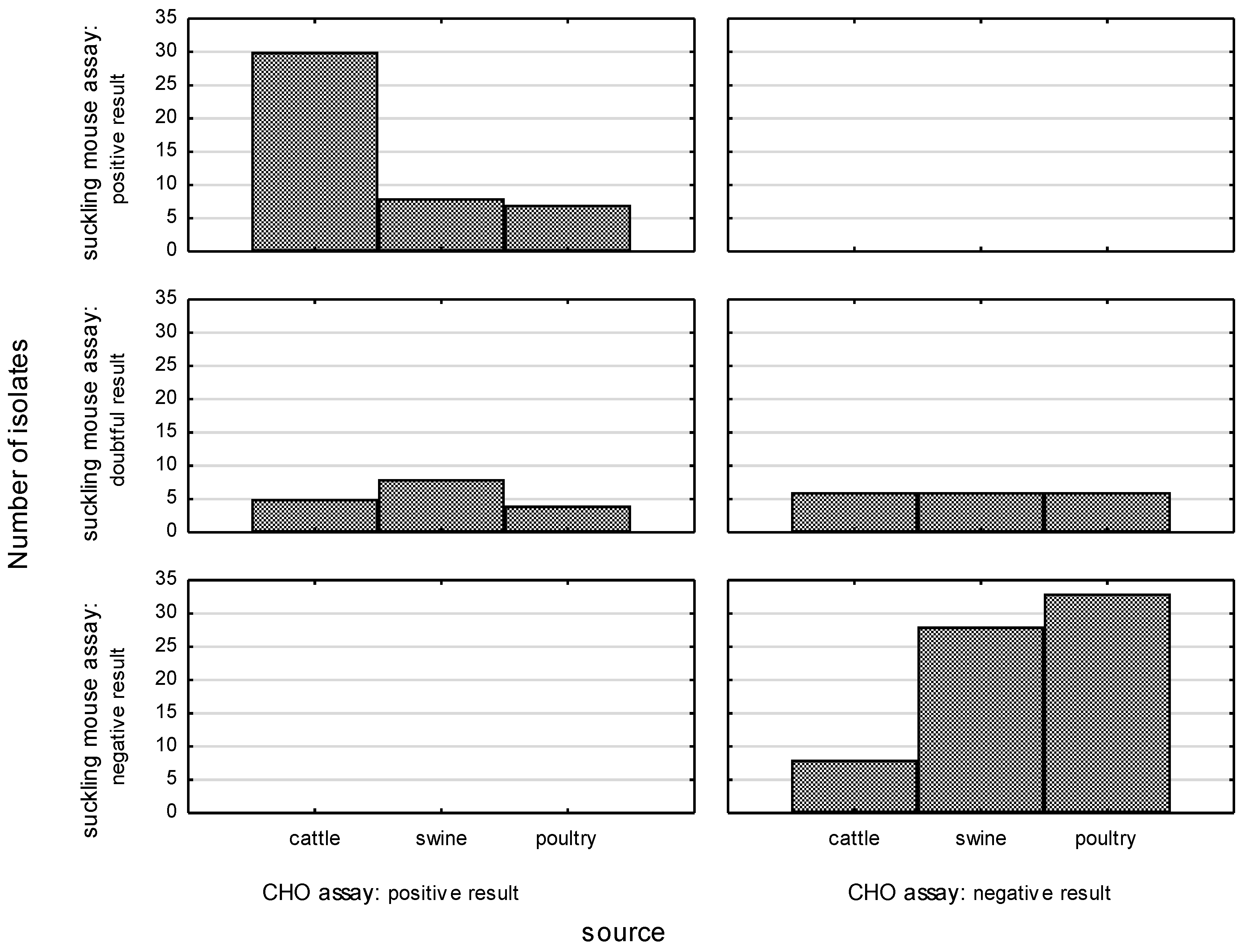
3.1. Enterotoxin Production Assay
3.1.1. The Influence of Enterotoxin on CHO Cells
3.1.2. Suckling Mouse Assay
3.2. Antimicrobial Resistance
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Igwaran, A.; Okoh, I. Human campylobacteriosis: A public health concern of global importance. Heliyon 2019, 5, e02814. [Google Scholar] [CrossRef] [PubMed]
- Kaakoush, N.O.; Castano-Rodriguez, N.; Mitchell, H.M.; Man, S.M. Global epidemiology of Campylobacter infection. Clin. Microbiol. Rev. 2015, 38, 687–720. [Google Scholar] [CrossRef] [PubMed]
- The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, 697. [CrossRef]
- Janssen, R.; Krogfelt, K.A.; Cawthraw, S.A.; van Pelt, W.; Wagenaar, J.A.; Owen, R.J. Host-pathogen interactions in Campylobacter infections: The host perspective. Clin. Microbiol. Rev. 2008, 21, 505–518. [Google Scholar] [CrossRef]
- Heredia, N.; García, S. Animals as sources of food-borne pathogens: A review. Anim. Nutr. 2018, 4, 250–255. [Google Scholar] [CrossRef]
- Toledo, Z.; Simaluiza, R.J.; Fernández, H. Occurrence and antimicrobial resistance of Campylobacter jejuni and Campylobacter coli isolated from domestic animals from Southern Ecuador. Cienc. Rural 2018, 48, e20180003. [Google Scholar] [CrossRef]
- Fernández, H. Campylobacter y campylobacteriosis: Una mirada desde América del Sur. Campylobacter and campylobacteriosis: A view from South America. Rev. Peru. Med. Exp. Salud. Publica 2011, 28, 121–127. [Google Scholar]
- Epps, S.V.; Harvey, R.B.; Hume, M.E.; Phillips, T.D.; Anderson, R.C.; Nisbet, D.J. Foodborne Campylobacter: Infections, metabolism, pathogenesis and reservoirs. Int. J. Environ. Res. Public Health. 2013, 10, 6292–6304. [Google Scholar] [CrossRef]
- Narvaez-Bravo, C.; Taboada, E.N.; Mutschall, S.K.; Aslam, M. Epidemiology of antimicrobial resistant Campylobacter spp. isolated from retail meats in Canada. Int. J. Food Microbiol. 2017, 253, 43–47. [Google Scholar] [CrossRef]
- Newell, D.G.; Mughini-Gras, L.; Kalupahana, R.S.; Wagenaar, J.A. Campylobacter epidemiology—Sources and routes of transmission for human infection. In Campylobacter, 1st ed.; Klein, G., Ed.; Elsevier Inc. Academic Press: Cambridge, MA, USA, 2017; pp. 85–110. [Google Scholar]
- Cody, A.J.; Maiden, M.C.J.; Strachan, N.J.C.; McCarthy, N.D. A systematic review of source attribution of human campylobacteriosis using multilocus sequence typing. Euro Surveill. 2019, 24, 1800696. [Google Scholar] [CrossRef]
- Wilson, D.J.; Gabriel, E.; Leatherbarrow, A.J.; Cheesbrough, J.; Gee, S.; Bolton, E.; Fox, A.; Fearnhead, P.; Hart, C.A.; Diggle, P.J. Tracing the source of campylobacteriosis. PLoS Genet. 2008, 4, e1000203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wysok, B.; Uradziński, J.; Wojtacka, J. Determination of the cytotoxic activity of Campylobacter strains isolated from bovine and swine carcasses in north-eastern Poland. Pol. J. Veter. Sci. 2015, 18, 579–586. [Google Scholar] [CrossRef] [PubMed]
- An, J.U.; Ho, H.; Kim, J.; Kim, W.H.; Kim, J.; Lee, S.; Mun, S.H.; Guk, J.H.; Hong, S.; Cho, S. Dairy cattle, a potential reservoir of human campylobacteriosis: Epidemiological and molecular characterization of Campylobacter jejuni from cattle farms. Front. Microbiol. 2018, 9, 3136. [Google Scholar] [CrossRef] [PubMed]
- Man, S.M. The clinical importance of emerging Campylobacter species. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 669–685. [Google Scholar] [CrossRef]
- Allos, B.M.; Blaser, M.J. Campylobacter jejuni and related species. In Principles and Practice of Infectious Diseases, 7th ed.; Mandell, G.L., Bennett, J.E., Dolin, R., Eds.; Elsevier: Philadelphia, PA, USA, 2009; pp. 2793–2802. [Google Scholar]
- Wassenaar, T.M. Toxin production by Campylobacter spp. Clin. Microbiol. Rev. 1997, 10, 466–476. [Google Scholar] [CrossRef]
- Coote, J.G.; Arain, T. A rapid, colourimetric assay for cytotoxin activity in Campylobacter jejuni. FEMS Immunol. Med. Microbiol. 1996, 13, 65–70. [Google Scholar] [CrossRef]
- Guévremont, E.; Higgins, R.; Quessy, S. Characterization of Campylobacter isolates recovered from clinically healthy pigs and from sporadic cases of campylobacteriosis in humans. J. Food Prot. 2004, 67, 228–234. [Google Scholar] [CrossRef]
- Pickett, C.L.; Pesci, E.C.; Cottle, D.L.; Russell, G.; Erdem, A.N.; Zeytin, H. Prevalence of cytolethal distending toxin production in Campylobacter jejuni and relatedness of Campylobacter sp. cdtB gene. Infect. Immun. 1996, 64, 2070–2078. [Google Scholar] [CrossRef]
- Findik, A.; Ica, T.; Onuk, E.E.; Percin, D.; Kevenk, T.O.; Ciftci, A. Molecular typing and cdt genes prevalence of Campylobacter jejuni isolates from various sources. Trop. Anim. Health Prod. 2011, 43, 711–719. [Google Scholar] [CrossRef]
- Pore, D.; Hoque, K.M.; Chakrabarti, M.K. Animal models in advancement of research in enteric diseases. In Animal Biotechnology, 2nd ed.; Ashish, S.V., Singh, A., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 199–219. [Google Scholar]
- Lipps, B.V. In vitro assay of biological and chemical toxins using antibodies against lethal toxin neutralizing factor. J. Venom. Anim. Toxins 2002, 8, 214–255. [Google Scholar] [CrossRef]
- Fernández, H.; Lobos, M.; Concha, M. Inducing Enterotoxigenic Properties in Campylobacter jejuni and Campylobacter coli by Serial Intraperitoneal Passage in Mice. Mem. Inst. Oswaldo Cruz 1999, 94, 101–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Palacios, G.M.; Torres, J.; Torres, N.I.; Escamilla, E.; Ruiz-Palacios, B.R.; Tamayo, J. Cholera-like enterotoxin produced by Campylobacter jejuni. Lancetii 1983, 30, 250–253. [Google Scholar] [CrossRef]
- Fernández, H.; Toro, Z. The relationship of Campylobacter jejuni subsp. jejuni enterotoxigenicity and the increase of cAMP and electrolyte changes in the rat intestine. Rev. Med. Chil. 1998, 126, 919–923. [Google Scholar] [PubMed]
- Klipstein, F.A.; Engert, R.F. Properties of crude Campylobacter jejuni heat-labile enterotoxin. Infect Immun. 1984, 45, 314–319. [Google Scholar] [CrossRef]
- Goossens, H.; Butzler, J.P.; Takeda, Y. Demonstration of cholera-like enterotoxin production by Campylobacter jejuni. FEMS Microbiol. Lett. 1985, 29, 73–76. [Google Scholar] [CrossRef]
- Luangtongkum, T.; Jeon, B.; Han, J.; Plummer, P.; Logue, C.M.; Zhang, Q. Antibiotic resistance in Campylobacter: Emergence, transmission and persistence. Future Microbiol. 2009, 4, 189–200. [Google Scholar] [CrossRef]
- Wieczorek, K.; Osek, J. Antimicrobial resistance mechanisms among Campylobacter. Biomed. Res. Int. 2013, 2013, 340605. [Google Scholar] [CrossRef]
- Reddy, S.; Zishiri, O.T. Detection and prevalence of antimicrobial resistance genes in Campylobacter spp. isolated from chickens and humans. J. Vet. Res. 2017, 84, e1–e6. [Google Scholar] [CrossRef]
- Silva, J.; Leite, D.; Fernandes, M.; Mena, C.; Gibbs, P.A.; Teixeira, P. Campylobacter spp. as a Foodborne Pathogen: A Review. Front. Microbiol. 2011, 2, 200. [Google Scholar] [CrossRef]
- Blaser, M.J.; Engberg, J. Clinical aspects of Campylobacter jejuni and Campylobacter coli infections. In Campylobacter; Nachamkin, I., Szymanski, C., Blaser, M.J., Eds.; ASM Press: Washington, DC, USA, 2008; pp. 99–121. [Google Scholar]
- Wieczorek, K.; Osek, J. Identification of virulence genes in Campylobacter jejuni and C. coli isolates by PCR. Bull. Vet. Inst. Pul. 2008, 52, 211–216. [Google Scholar]
- Engberg, J.; Aarestrup, F.M.; Taylor, D.E.; Gerner-Smidt, P.; Nachamkin, I. Quinolone and macrolide resistance in Campylobacter jejuni and C. coli: Resistance mechanisms and trends in human isolates. Emerg. Infect. Dis. 2001, 7, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Alfredson, D.A.; Korolik, V. Antibiotic resistance and resistance mechanisms in Campylobacter jejuni and Campylobacter coli. FEMS Microbiol. Lett. 2007, 277, 123–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, L.; Lee, Y. Antimicrobial resistance of 114 porcine isolates of Campylobacter coli. Int. J. Food Microbiol. 2007, 118, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Wysok, B.; Wojtacka, J.; Kivistö, R. Pathogenicity of Campylobacter strains of poultry and human origin from Poland. Int. J. Food Microbiol. 2020, 334, 108830. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, K.; Osek, J. Multiplex PCR assays for simultaneous identification of Campylobacter jejuni and Campylobacter coli. Med. Weter 2005, 61, 797–799. [Google Scholar]
- Belbouri, A.; Mégraud, F. Enterotoxin-like activity produced by Campylobacter jejuni and Campylobacter coli isolated from patients and healthy controls in Algeria. FEMS Microbiol. Lett. 1988, 51, 25–28. [Google Scholar] [CrossRef]
- Giannella, R.A. Suckling mouse model for detection of heat-stable Escherichia coli enterotoxin: Characteristics of the model. Infect. Immun. 1976, 14, 95–99. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. EU Protocol for Harmonised Monitoring of Antimicrobial Resistance in Human Salmonella and Campylobacter Isolates; ECDC: Stockholm, Sweden, 2016; Available online: https://www.ecdc.europa.eu/en/publications-data/eu-protocol-harmonised-monitoring-antimicrobial-resistance-human-salmonella-and-0 (accessed on 10 September 2022).
- Odeyemi, O.A.; Sani, N.A. Antibiotic resistance and burden of foodborne diseases in developing countries. Future Sci. OA 2016, 2, FSO139. [Google Scholar] [CrossRef]
- Wilson, J.W.; Schurr, M.J.; LeBlanc, C.L.; Ramamurthy, R.; Buchanan, K.L.; Nickerson, C.A. Mechanisms of bacterial pathogenicity. Postgrad. Med. J. 2002, 78, 216–224. [Google Scholar] [CrossRef]
- Biswas, D.; Fernando, U.; Reiman, C.; Willson, P.; Potter, A.; Allan, B. Effect of cytolethal distending toxin of Campylobacter jejuni on adhesion and internalization in cultured cells and in colonization of the chicken gut. Avian Dis. 2006, 50, 586–593. [Google Scholar] [CrossRef]
- Fernández, H.; Trabulsi, L.R. Invasive and enterotoxic properties in Campylobacter jejuni and Campylobacter coli strains isolated from humans and animals. Biol Res. 1995, 28, 205–210. [Google Scholar] [PubMed]
- Florin, I.; Antillon, F. Production of enterotoxin and cytotoxin in Campylobacter jejuni strains isolated in Costa Rica. J. Med. Microbiol. 1992, 37, 22–29. [Google Scholar] [CrossRef]
- Chattopadhyay, U.K.; Rathore, R.S.; Pal, D.; Das, M.S. Enterotoxigenicity of human and animal isolates of Campylobacter jejuni in ligated rat ileal loops. J. Diarrhoeal. Dis. Res. 1991, 9, 20–22. [Google Scholar] [PubMed]
- Perez-Perez, G.I.; Cohn, D.L.; Guerrant, R.I.; Patton, C.M.; Reller, L.B.; Blaser, M.J. Clinical and immunologic significance of cholera-like toxin and cytotoxin production by Campylobacter species in patients with acute inflammatory diarrhea in the USA. J. Infect. Dis. 1989, 160, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Konkel, M.E.; Lobet, Y.; Cieplak, W. Examination of multiple isolates of Campylobacter jejuni for evidence of cholera toxin-like activity. In Campylobacter jejuni: Current Status and Future Trends; Nachamkin, I., Blaser, M.J., Tompkins, L.S., Eds.; American Society for Microbiology: Washington, DC, USA, 1992; pp. 193–198. [Google Scholar]
- Akhtar, S.Q.; Huq, F. Effect of Campylobacter jejuni extracts and culture supernatants on cell culture. J. Trop. Med. Hyg. 1989, 92, 80–85. [Google Scholar]
- Thornley, J.P.; Jenkins, D.; Neal, K.; Wright, T.; Brough, J.; Spiller, R.C. Relationship of Campylobacter toxigenicity in vitro to the development of postinfectious irritable bowel syndrome. J. Infect. Dis. 2001, 184, 606–609. [Google Scholar] [CrossRef]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health. 2015, 109, 309–318. [Google Scholar] [CrossRef]
- Friedman, N.D.; Temkin, E.; Carmeli, Y. The negative impact of antibiotic resistance. Clin. Microbiol. Infect. 2016, 22, 416–422. [Google Scholar] [CrossRef]
- Marshall, B.M.; Levy, S.B. Food animals and antimicrobials: Impacts on human health. Clin. Microbiol. Rev. 2011, 24, 718–733. [Google Scholar] [CrossRef]
- Trieber, C.A.; Taylor, D.E. Mechanisms of antibiotic resistance in Campylobacter. In Campylobacter, 2nd ed.; Nachamkin, I., Blaser, M.J., Eds.; ASM Press: Washington DC, USA, 2000; pp. 441–454. [Google Scholar]
- Wieczorek, K.; Osek, J. Poultry flocks as a source of Campylobacter contamination of broiler carcasses. Pol. J. Veter. Sci. 2015, 18, 101–106. [Google Scholar] [CrossRef]
- Padungtod, P.; Kaneene, J.B.; Hanson, R.; Morita, Y.; Boonmar, S. Antimicrobial resistance in Campylobacter isolated from food animals and humans in northern Thailand. FEMS Immunol. Med. Microbiol. 2006, 47, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Moser, S.; Seth-Smith, H.; Egli, A.; Kittl, S.; Overesch, G. Campylobacter jejuni from canine and bovine cases of campylobacteriosis express high antimicrobial resistance rates against (Fluoro)quinolones and Tetracyclines. Pathogens 2020, 9, 691. [Google Scholar] [CrossRef] [PubMed]
- The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2017. EFSA J. 2019, 17, 5598.
- Elhadidy, M.; Ali, M.M.; El-Shibiny, A.; Miller, W.G.; Elkhatib, W.F.; Botteldoorn, N.; Dierick, K. Antimicrobial resistance patterns and molecular resistance markers of Campylobacter jejuni isolates from human diarrheal cases. PLoS ONE 2020, 15, e0227833. [Google Scholar] [CrossRef]
- Wieczorek, K.; Szewczyk, R.; Osek, J. Prevalence, antimicrobial resistance, and molecular characterization of Campylobacter jejuni and C. coli isolated from retail raw meat in Poland. Veterinarni Medicina 2012, 57, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Kempf, I.; Kerouanton, A.; Bougeard, S.; Nagard, B.; Rose, V.; Mourand, G.; Osterberg, J.; Denis, M.; Bengtsson, B.O. Campylobacter coli in Organic and Conventional Pig Production in France and Sweden: Prevalence and Antimicrobial Resistance. Front. Microbiol. 2017, 8, 955. [Google Scholar] [CrossRef]
- Bolinger, H.; Kathariou, S. The current state of macrolide resistance in Campylobacter spp.: Trends and impacts of resistance mechanisms. Appl. Environ. Microbiol. 2017, 83, e00416–e00417. [Google Scholar] [CrossRef]
- Fernández, H.; Mansilla, M.; González, V. Antimicrobial Susceptibility of Campylobacter jejuni subsp. jejuni Assessed by E-test and Double Dilution Agar Method in Southern Chile. Mem. Inst. Oswaldo Cruz 2000, 95, 247–249. [Google Scholar] [CrossRef]
- Kashoma, I.P.; Kassem, I.I.; Kumar, A.; Kessy, B.M.; Gebreyes, W.; Kazwala, R.R.; Rajashekara, G. Antimicrobial Resistance and Genotypic Diversity of Campylobacter Isolated from Pigs, Dairy, and Beef Cattle in Tanzania. Front. Microbiol. 2015, 6, 1240. [Google Scholar] [CrossRef]
- Nguyen, T.N.; Hotzel, H.; El-Adawy, H.; Tran, H.T.; Le, M.T.; Tomaso, H.; Neubauer, H.; Hafez, H.M. Genotyping and antibiotic resistance of thermophilic Campylobacter isolated from chicken and pig meat in Vietnam. Gut Pathog. 2016, 8, 19. [Google Scholar] [CrossRef]
- Tang, Y.; Fang, L.; Xu, C.; Zhang, Q. Antibiotic resistance trends and mechanisms in the foodborne pathogen, Campylobacter. Anim. Health Res. Rev. 2017, 18, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Mukherjee, S.; Chen, Y.; Li, C.; Young, S.; Warren, M.; Abbott, J.; Friedman, S.; Kabera, C.; Karlsson, M.; et al. Novel gentamicin resistance genes in Campylobacter isolated from humans and retail meats in the USA. J. Antimicrob. Chemother. 2015, 70, 1314–1321. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, F.; Colston, J.M.; Paredes-Olortegui, M.; François, R.; Pisanic, N.; Burga, R.; Peñataro-Yori, P.; Kosek, M.N. Antibiotic Resistance of Campylobacter Species in a Pediatric Cohort Study. Antimicrob. Agents Chemother. 2019, 63, e01911–e01918. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Ge, Y.; Xu, H.; Zhang, J.; Kuang, D.; Yang, X.; Su, X.; Huang, Z.; Shi, X.; Xu, X.; et al. Molecular characterization, antimicrobial resistance and Caco-2 cell invasion potential of Campylobacter jejuni/coli from young children with diarrhea. Pediatr. Infect. Dis. J. 2016, 35, 330–334. [Google Scholar] [CrossRef] [PubMed]
- García-Fernández, A.; Dionisi, A.M.; Arena, S.; Iglesias-Torrens, Y.; Carattoli, A.; Luzzi, I. Human Campylobacteriosis in Italy: Emergence of Multi-Drug Resistance to Ciprofloxacin, Tetracycline, and Erythromycin. Front. Microbiol. 2018, 9, 1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Young, S.R.; Tong, E.; Abbott, J.W.; Womack, N.; Friedman, S.L.; McDermott, P.F. Antimicrobial resistance of Campylobacter isolates from retail meat in the United States between 2002 and 2007. Appl. Environ. Microbiol. 2010, 76, 7949–7956. [Google Scholar] [CrossRef]
- Aksomaitiene, J.; Ramonaite, S.; Tamuleviciene, E.; Novoslavskij, A.; Alter, T.; Malakauskas, M. Overlap of Antibiotic Resistant Campylobacter jejuni MLST Genotypes Isolated From Humans, Broiler Products, Dairy Cattle and Wild Birds in Lithuania. Front. Microbiol. 2019, 10, 1377. [Google Scholar] [CrossRef]
- Wieczorek, K.; Kania, I.; Osek, J. Prevalence and antimicrobial resistance of Campylobacter spp. isolated from poultry carcasses in Poland. J. Food Prot. 2013, 76, 1451–1455. [Google Scholar] [CrossRef]
- Gibreel, A.; Taylor, D.E. Macrolide resistance in Campylobacter jejuni and Campylobacter coli. J. Antimicrob. Chemother. 2006, 58, 243–255. [Google Scholar] [CrossRef]


{kind=link}
| Target Gene | Sequences (5′–3′) | PCR Product Size (bp) | References |
|---|---|---|---|
| 16S rRNA for Campylobacter spp. | F—ATCTAATGGCTTAACCATTAAAC R—GGACGGTAACTAGTTTAGTATT | 857 | [39] |
| mapA for C. jejuni | F—CTATTTTATTTTTGAGTGCTTGTG R—GCTTTATTTGCCATTTGTTTTATTA | 589 | [39] |
| ceuE for C. coli | F—AATTGAAAATTGCTCCAACTATG R—TGATTTTATTATTTGTAGCAGCG | 462 | [39] |
| Resistance Pattern | Swine Isolates | Poultry Isolates | Cattle Isolates | ||
|---|---|---|---|---|---|
| C. coli (n = 46) | C. jejuni (n = 4) | C. coli (n = 3) | C. jejuni (n = 47) | C. jejuni (n = 49) | |
| CIP_E_TET | 6 | 1 | − | 2 | 2 |
| CIP_TET_G | − | 1 | − | − | − |
| CIP_TET | 26 | 1 | 1 | 24 | 16 |
| CIP_E | − | − | − | − | 1 |
| TET | 6 | − | 1 | 10 | 10 |
| CIP | 7 | − | 1 | 5 | 6 |
| E | − | − | − | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wysok, B.; Wojtacka, J.; Wiszniewska-Łaszczych, A.; Sołtysiuk, M.; Kobuszewska, A. The Enterotoxin Production and Antimicrobial Resistance of Campylobacter Strains Originating from Slaughter Animals. Pathogens 2022, 11, 1131. https://doi.org/10.3390/pathogens11101131
Wysok B, Wojtacka J, Wiszniewska-Łaszczych A, Sołtysiuk M, Kobuszewska A. The Enterotoxin Production and Antimicrobial Resistance of Campylobacter Strains Originating from Slaughter Animals. Pathogens. 2022; 11(10):1131. https://doi.org/10.3390/pathogens11101131
Chicago/Turabian StyleWysok, Beata, Joanna Wojtacka, Agnieszka Wiszniewska-Łaszczych, Marta Sołtysiuk, and Aleksandra Kobuszewska. 2022. "The Enterotoxin Production and Antimicrobial Resistance of Campylobacter Strains Originating from Slaughter Animals" Pathogens 11, no. 10: 1131. https://doi.org/10.3390/pathogens11101131
APA StyleWysok, B., Wojtacka, J., Wiszniewska-Łaszczych, A., Sołtysiuk, M., & Kobuszewska, A. (2022). The Enterotoxin Production and Antimicrobial Resistance of Campylobacter Strains Originating from Slaughter Animals. Pathogens, 11(10), 1131. https://doi.org/10.3390/pathogens11101131