
The Prevalence of Pathogens among Ticks Collected from Livestock in Kazakhstan
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
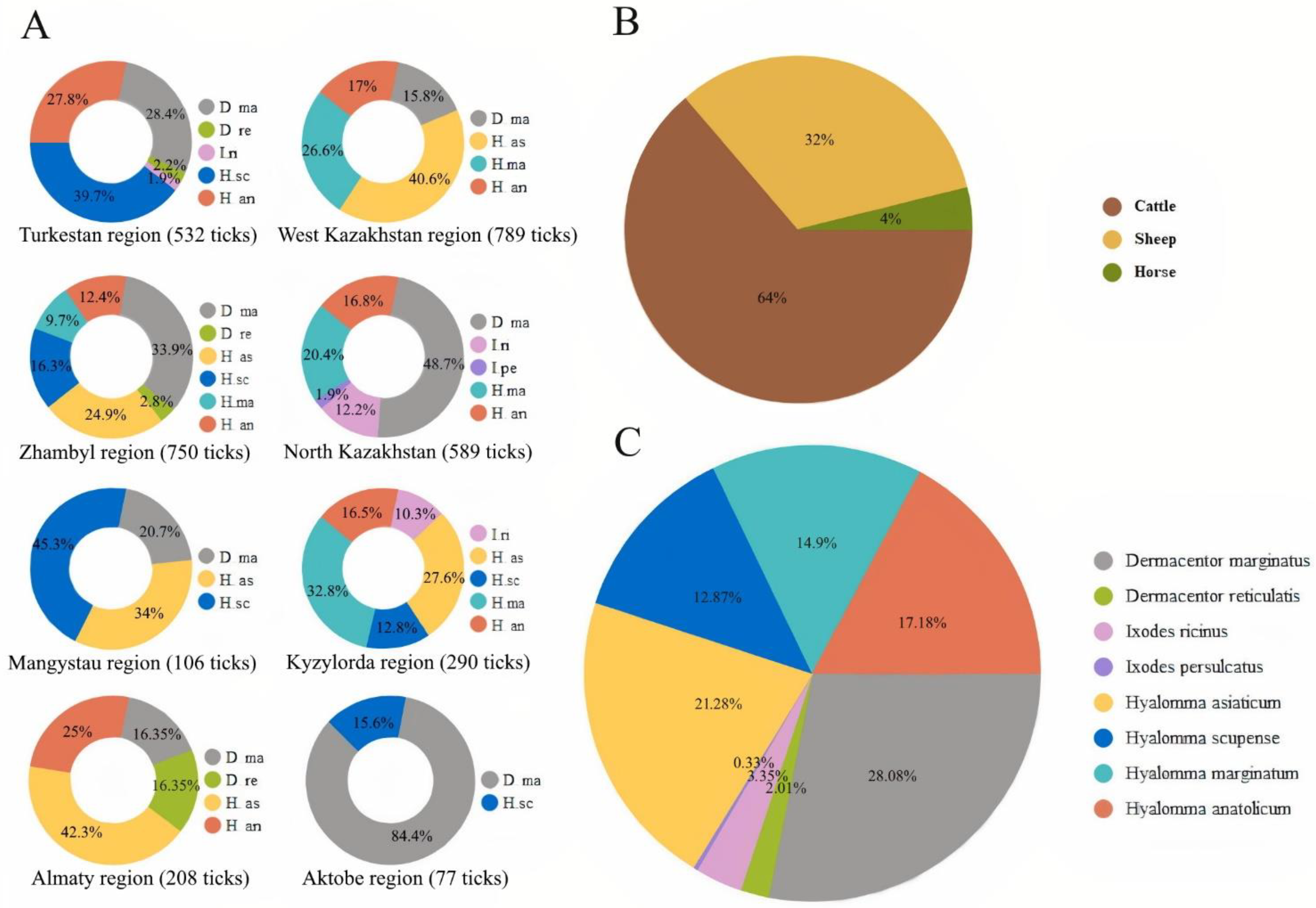
2.1. Species Composition of Ixodid Ticks in Livestock
2.2. PCR Detection of Tick-Borne Pathogens
2.3. Sequence Analysis of Tick-Borne Pathogens
3. Discussion
4. Materials and Methods
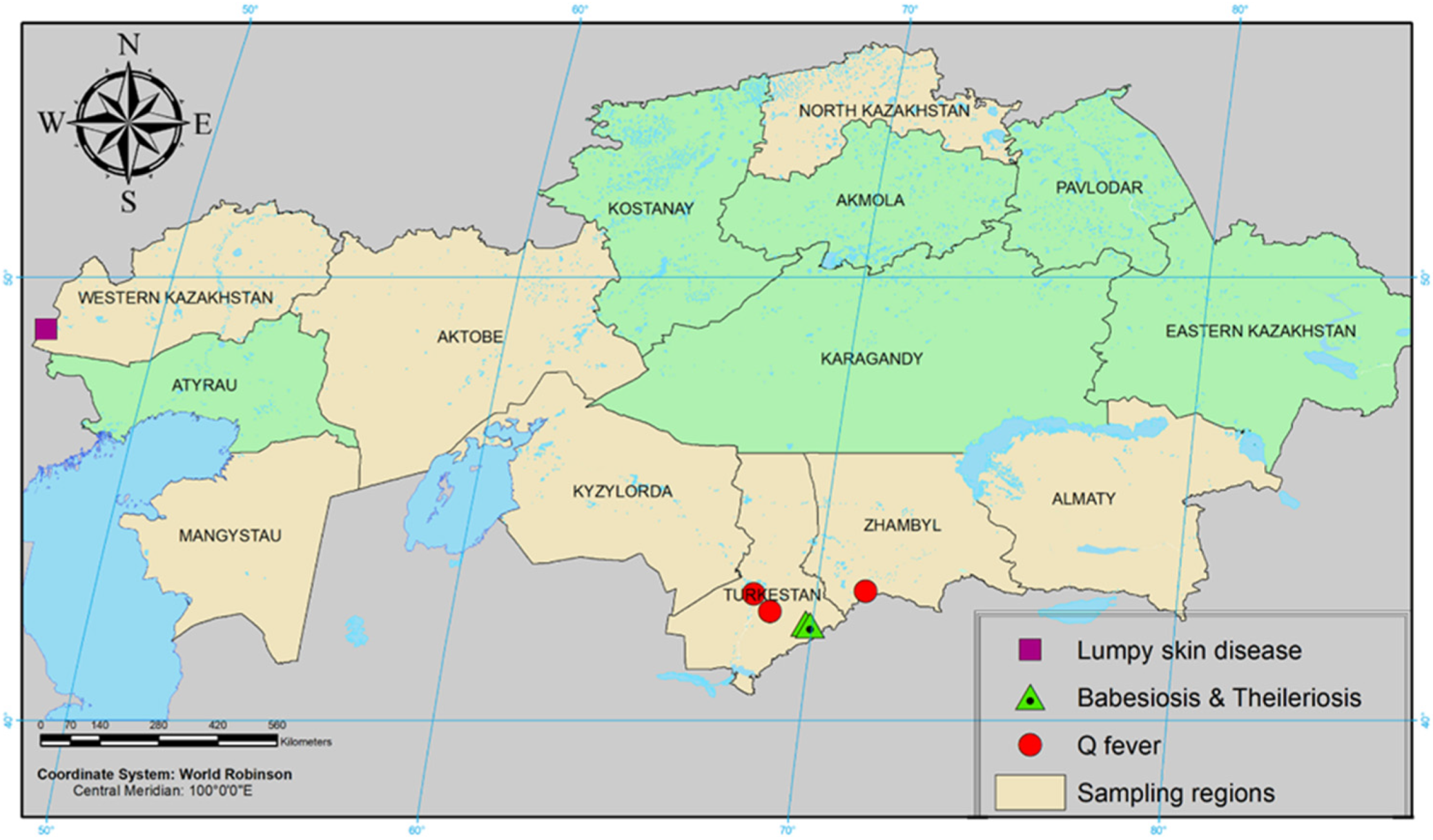
4.1. Study Area and Collection of Samples
4.2. Sample Preparation and DNA/RNA Purification
4.3. Confirmation of the Presence of Tick-Borne Pathogens by PCR Method
4.4. Sequencing Assays, BLASTn Analysis and Phylogenetic Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rajput, Z.I.; Hu, S.-H.; Chen, W.-J.; Arijo, A.G.; Xiao, C.-W. Importance of ticks and their chemical and immunological control in livestock. J. Zhejiang Univ. Sci. B 2006, 7, 912–921. [Google Scholar] [CrossRef] [PubMed]
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, S3–S14. [Google Scholar] [CrossRef] [PubMed]
- Harrus, S.; Baneth, G. Drivers for the emergence and re-emergence of vector-borne protozoal and bacterial diseases. Int. J. Parasitol. 2005, 35, 1309–1318. [Google Scholar] [CrossRef]
- Perfilyeva, Y.V.; Shapiyeva, Z.Z.; Ostapchuk, Y.O.; Berdygulova, Z.A.; Bissenbay, A.O.; Kulemin, M.V.; Ismagulova, G.A.; Skiba, Y.A.; Sayakova, Z.Z.; Mamadaliyev, S.M.; et al. Tick-borne pathogens and their vectors in Kazakhstan—A review. Ticks Tick-Borne Dis. 2020, 11, 101498. [Google Scholar] [CrossRef] [PubMed]
- Sultankulova, K.T.; Shynybekova, G.O.; Kozhabergenov, N.S.; Mukhami, N.N.; Chervyakova, O.V.; Burashev, Y.D.; Zakarya, K.D.; Nakhanov, A.K.; Barakbayev, K.B.; Orynbayev, M.B. The Prevalence and Genetic Variants of the CCHF Virus Circulating among Ticks in the Southern Regions of Kazakhstan. Pathogens 2022, 11, 841. [Google Scholar] [CrossRef] [PubMed]
- Dobler, G.; Erber, W. The TBE Book, 2nd ed.; Global Health Press: Singapore, 2019; pp. 189–195. [Google Scholar] [CrossRef]
- Sang, C.; Yang, M.; Xu, B.; Liu, G.; Yang, Y.; Kairullayev, K.; Bauyrzhan, O.; Hazihan, W.; Hornok, S.; Wang, Y. Tick distribution and detection of Babesia and Theileria species in Eastern and Southern Kazakhstan. Ticks Tick-Borne Dis. 2021, 12, 101817. [Google Scholar] [CrossRef]
- Head, J.R.; Bumburidi, Y.; Mirzabekova, G.; Rakhimov, K.; Dzhumankulov, M.; Salyer, S.J.; Knust, B.; Berezovskiy, D.; Kulatayeva, M.; Zhetibaev, S.; et al. Risk Factors for and Seroprevalence of Tickborne Zoonotic Diseases among Livestock Owners, Kazakhstan. Emerg. Infect. Dis. 2020, 26, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Orynbayev, M.B.; Rystaeva, R.A. Isolation of Coxiella burnetii from ticks in the Republic of Kazakhstan. Biosecur. Biotechnol. 2020, 1, 62–67. (In Russian) [Google Scholar]
- Kurmangaliyeva, K.; Atygaeva, S. Case of Lyme disease in a non-endemic-region. J. Infectol. 2014, 6, 93–94. (In Russian) [Google Scholar]
- Temirbekov, Z.; Kiryushchenko, T. A new focus of Crimean hemorrhagic fever in the Moiynkum district of the Dzhambul region. In Regional Features of the Epidemiology of Infectious Diseases in Kazakhstan; Research Institute of Epidemiology, Microbiology and Infectious Diseases: Almaty, Kazakhstan, 1984; pp. 122–125. (In Russian) [Google Scholar]
- Dmitrovskiy, A. TBE in Kazakhstan. Tick-borne encephalitis. In The TBE Book, 2nd ed.; Dobler, G., Erber, W., Eds.; Global Health Press: Singapore, 2019; pp. 189–195. [Google Scholar]
- Tutushin, M.I. Distribution and Seasonal and Age Variation of Bovine Theileriasis in the South of Kazakhstan; Vostochnoe Otdelenie VASKhNIL: Alma-Ata, USSR, 1981; pp. 130–134. (In Russian) [Google Scholar]
- Dennis, D.T.; Piesman, J.F. Overview of tick-borne infections of humans. In Tick-Borne Diseases of Humans; Goodman, J.L., Den nis, D.T., Sonenshine, D.E., Eds.; ASM Press: Washington, DC, USA, 2005; pp. 3–11. [Google Scholar]
- Estrada-Peña, A.; Salman, M. Current limitations in the control and spread of ticks that affect livestock: A review. Agriculture 2013, 3, 221–235. [Google Scholar] [CrossRef] [Green Version]
- Rochlin, I.; Toledo, A. Emerging tick-borne pathogens of public health importance: A mini-review. J. Med. Microbiol. 2020, 69, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Jiang, M.; Yang, M.; Bo, X.; Zhao, S.; Zhang, Y.; Wureli, H.; Wang, B.; Tu, C.; Wang, Y. Tick distribution in border regions of Northwestern China. Ticks Tick-Borne Dis. 2019, 10, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Tanitovskiy, V.; Maikanov, N. Fauna of ixodid ticks of western Kazakhstan and their distribution across the territory. Vestn. WKSU 2018, 2, 2–12. (In Russian) [Google Scholar]
- Maikanov, N.; Ospanov, B. The ticks’ fauna of the natural foci territory of Atyrau and West-Kazakhstan regions. Sc. J. Nat. Cent. Inf. Dis. Nat. Foci 2008, 3, 187–190. (In Russian) [Google Scholar]
- Nurmakhanov, T.; Sansyzbayev, Y.; Atshabar, B.; Deryabin, P.; Kazakov, S.; Zholshorinov, A.; Matzhanova, A.; Sadvakassova, A.; Saylaubekuly, R.; Kyraubaev, K.; et al. Crimean-Congo hemorrhagic fever virus in Kazakhstan (1948–2013). Int. J. Infect. Dis. 2015, 38, 19–23. [Google Scholar] [CrossRef] [Green Version]
- Chihota, C.M.; Rennie, L.F.; Kitching, R.P.; Mellor, P.S. Attempted mechanical transmission of lumpy skin disease virus by biting insects. Med. Vet.-Èntomol. 2003, 17, 294–300. [Google Scholar] [CrossRef]
- Wainwright, S.; El Idrissi, A.; Mattioli, R.; Tibbo, M.; Njeumi, F.; Raizman, E. Emergence of lumpy skin disease in the Eastern Mediterranean Basin countries. FAO Empres Watch. 2013, 29, 1–6. [Google Scholar]
- Zeynalova, S.; Asadov, K.; Guliyev, F.; Vatani, M.; Aliyev, V. Epizootology and molecular diagnosis of lumpy skin disease among livestock in Azerbaijan. Front. Microbiol. 2016, 7, 1022. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority (EFSA). Scientific report on lumpy skin disease: I. Data collection and analysis. EFSA J. 2017, 15, 4773. [Google Scholar]
- Azeem, S.; Sharma, B.; Shabir, S.; Akbar, H.; Venter, E. Lumpy skin disease is expanding its geographic range: A challenge for Asian livestock management and food security. Vet. J. 2022, 279, 105785. [Google Scholar] [CrossRef]
- Roche, X.; Rozstalnyy, A.; TagoPacheco, D.; Pittiglio, C.; Kamata, A.; Beltran Alcrudo, D.; Bisht, K.; Karki, S.; Kayamori, J.; Larfaoui, F.; et al. Introduction and spread of lumpy skin disease in South, East and Southeast Asia: Qualitative risk assessment and management. In FAO Animal Production and Health; Paper 183; FAO: Rome, Italy, 2020. [Google Scholar]
- Gazimagomedov, M.; Kabardiev, S.; Bittirov, A.; Abdulmagomedov, S.; Ustarov, R.; Musaev, Z.; Bittirova, A. Specific composition of Ixodidae ticks and their role in transmission of nodular dermatitis virus among cattle in the North Caucasus. J. Theory Pract. Combat. Parasit. Dis. 2017, 8, 107–110. (In Russian) [Google Scholar]
- Orynbayev, M.B.; Nissanova, R.K.; Khairullin, B.M.; Issimov, A.; Zakarya, K.D.; Sultankulova, K.T.; Kutumbetov, L.B.; Tulendibayev, A.B.; Myrzakhmetova, B.S.; Burashev, E.D.; et al. Lumpy skin disease in Kazakhstan. Trop. Anim. Health Prod. 2021, 53, 166. [Google Scholar] [CrossRef] [PubMed]
- Tissot-Dupont, H.; Raoult, D. Q fever. Infect. Dis. Clin. N. Am. 2008, 22, 505–514. [Google Scholar] [CrossRef] [PubMed]
- van Roeden, S.E.; Wever, P.C.; Kampschreur, L.; Gruteke, P.; van der Hoek, W.; Hoepelman, A.; Bleeker-Rovers, C.; Oosterheert, J. Chronic Q fever-related complications and mortality: Data from a nationwide cohort. Clin. Microbiol. Infect. 2019, 25, 1390–1398. [Google Scholar] [CrossRef] [PubMed]
- van der Hoek, W.; Dijkstra, F.; Schimmer, B.; Schneeberger, P.M.; Vellema, P.; Wijkmans, C.; ter Schegget, R.; Hackert, V.; van Duynhoven, Y. Q fever in the Netherlands: An update on the epidemiology and control measures. Eurosurveillance 2010, 15, 19520. [Google Scholar] [CrossRef]
- Cross, A.R.; Baldwin, V.M.; Roy, S.; Essex-Lopresti, A.E.; Prior, J.L.; Harmer, N.J. Zoonoses under our noses. Microbes Infect. 2019, 21, 10–19. [Google Scholar] [CrossRef]
- El-Mahallawy, H.S.; Lu, G.; Kelly, P.; Xu, D.; Li, Y.; Fan, W.; Wang, C. Q fever in China: A systematic review, 1989–2013. Epidemiol. Infect. 2015, 143, 673–681. [Google Scholar] [CrossRef]
- Körner, S.; Makert, G.R.; Ulbert, S.; Pfeffer, M.; Mertens-Scholz, K. The Prevalence of Coxiella burnetii in Hard Ticks in Europe and Their Role in Q Fever Transmission Revisited—A Systematic Review. Front. Vet. Sci. 2021, 8, 655715. [Google Scholar] [CrossRef]
- Tsoi, D.C.; Rapoport, L.P.; Samartseva, E.T. Epidemiological studies of Q fever in the area of Dzhambul in the Kazakh SSR. J. Hyg. Epidemiol. Microbiol. Immunol. 1980, 24, 206–211. (In Russian) [Google Scholar]
- Orynbayev, M.B.; Beauvais, W.; Sansyzbay, A.R.; Rystaeva, R.A.; Sultankulova, K.T.; Kerimbaev, A.A.; Kospanova, M.N.; Kock, R.A. Seroprevalence of infectious diseases in saiga antelope (Saiga tatarica tatarica) in Kazakhstan 2012–2014. Prev. Vet. Med. 2016, 127, 100–104. [Google Scholar] [CrossRef] [Green Version]
- Pchelkina, A.A.; Zhmabva, Z.M.; Zubkova, R.I. Q Fever in Northern Kazakhstan. J. Microbiol. Epidemiol. Immunobiol. 1956, 11, 32–35. [Google Scholar]
- Bilgic, H.B.; Karagenç, T.; Shiels, B.; Tait, A.; Eren, H.; Weir, W. Evaluation of cytochrome b as a sensitive target for PCR based detection of T. annulata carrier animals. Vet. Parasitol. 2010, 174, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Purnell, R. Theileria annulata as a hazard to cattle in countries on the northern Mediterranean littoral. Vet. Sci. Commun. 1978, 2, 3–10. [Google Scholar] [CrossRef]
- Bishop, R.P.; Odongo, D.O.; Mann, J. Theileria. In Genome Mapping and Genomics in Animal-Associated Microbes; Springer: Berlin/Heidelberg, Germany, 2009; pp. 191–231. [Google Scholar]
- Gharbi, M.; Darghouth, M.A. A review of Hyalomma scupense (Acari, Ixodidae) in the Maghreb region: From biology to control. Parasite 2014, 21, 12–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaman, M.A.; Mehreen, U.; Qamar, W.; Qamar, M.F.; Kashif, M.; Shahid, Z.; Abbas, R.Z. Brief account of bovine theileriosis prevalence in some South Asian countries. Agrobiol. Rec. 2020, 2, 38–48. [Google Scholar] [CrossRef]
- Selim, A.; Weir, W.; Khater, H. Prevalence and risk factors associated with tropical theileriosis in Egyptian dairy cattle. Vet. World 2022, 15, 919–924. [Google Scholar] [CrossRef]
- Shabdarbayeva, G.S.; Abdybekova, A.M. Identification of foci of blood-parasitic diseases of ruminants in the south of Kazakhstan. Eurasian Union Sci. 2016, 33, 17–21. (In Russian) [Google Scholar]
- Onyiche, T.E.; Suganuma, K.; Igarashi, I.; Yokoyama, N.; Xuan, X.; Thekisoe, O. A Review on Equine Piroplasmosis: Epidemiology, Vector Ecology, Risk Factors, Host Immunity, Diagnosis and Control. Int. J. Environ. Res. Public Health 2019, 16, 1736. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, J.J. Review on Bovine Babesiosis and Its Economical Importance. Austin J. Vet. Sci. Anim. Husb. 2017, 4, 1035. [Google Scholar]
- Machado, R.Z.; Toledo, C.Z.P.; Teixeira, M.C.A.; André, M.R.; Freschi, C.R.; Sampaio, P.H. Molecular and serological detection of Theileria equi and Babesia caballi in donkeys (Equus asinus) in Brazil. Vet. Parasitol. 2012, 186, 461–465. [Google Scholar] [CrossRef]
- Acici, M.; Umur, S.; Guvenc, T.; Arslan, H.H.; Kurt, M. Seroprevalence of equine babesiosis in the Black Sea region of Turkey. Parasitol Int. 2008, 57, 198–200. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.Y.; Wang, C.H.; Wu, Y.L. Serological survey of equine piroplasmosis, equine granulocytic anaplasmosis, and equine Lyme disease in Taiwan. Taiwan Vet. J. 2010, 36, 261–267. [Google Scholar]
- Butler, C.M.; van Oldruitenborgh-Oosterbaan, M.M.S.; Stout, T.A.; van der Kolk, J.H.; Van den Wollenberg, L.; Nielen, M.; Jongejan, F.; Werners, A.H.; Houwers, D.J. Prevalence of the causative agents of equine piroplasmosis in the South West of The Netherlands and the identification of two autochthonous clinical Theileria equi infections. Vet. J. 2012, 193, 381–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laus, F.; Spaterna, A.; Faillace, V.; Veronesi, F.; Ravagnan, S.; Beribè, F.; Cerquetella, M.; Meligrana, M.C.T.; Tesei, B. Clinical investigation on Theileria equi and Babesia caballi infections in Italian donkeys. BMC Vet. Res. 2015, 28, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scoles, G.A.; Ueti, M.W. Vector ecology of equine piroplasmosis. Annu. Rev. Entomol. 2015, 7, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Fillipova, N.A.; Musatov, S.A.; Panova, I.V.; Lobanov, A.L. Taxonomic structure of the polytypic species Hyalomma asiaticum (Ixodidae). The first experience of using databases on morphometry. Parasitology 1995, 29, 65–82. (In Russian) [Google Scholar]
- Kerbabayev, E.B. Fundamentals of veterinary acarology. Methods and means of combating ticks. Proc. All-Russ. Inst. Helminthol. 1998, 34, 218. (In Russian) [Google Scholar]
- Siuda, K. Ticks of Poland (Acari: Ixodidae). In Systematic and Distribution; Polish Parasitological Society: Warsaw, Poland, 1993; Volume 2, p. 372. (In Polish) [Google Scholar]
- Hoover, T.A.; Vodkin, M.H.; Williams, J.C. A Coxiella burnetti repeated DNA element resembling a bacterial insertion sequence. J. Bacteriol. 1992, 174, 5540–5548. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Scheme | Positive Ticks per Pathogen Species (%) | ||||||
|---|---|---|---|---|---|---|---|
| Tick | Location | Host | No. | LSDV | Coxiella burnetti | Teileria annulata | Babesia caballi |
| D. ma | Bokey Orda district, West Kazakhstan region | Cattle | 49 | 7 (14.28) | - | - | - |
| H. as | Bokey Orda district, West Kazakhstan region | Cattle | 105 | 6 (5.71) | - | - | - |
| D. ma | Otrar district, Turkestan region | Cattle | 47 | - | 15 (31.91) | - | - |
| H. sc | Tolebi district, Turkestan region | Horse | 35 | - | - | - | 6 (17.14) |
| H. sc | Tolebi district, Turkestan region | Cattle | 82 | - | - | 6 (7.32) | - |
| D. ma | Tolebi district, Turkestan region | Cattle | 64 | - | - | 5 (6.10) | - |
| H. an | Taraz city, Zhambyl region | Cattle | 19 | - | 10 (52.63) | - | - |
| Host | Tick Species | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Location | Host Animals | No. | D. ma | D. re | I. ri | I. pe | H. as | H. sc | H. ma | H. an |
| Otrar district, Turkestan region N 42°51′25″ E 68°3′5″ | Cattle | 13 | 47 | 12 | 10 | - | - | 42 | - | 72 |
| Sheep | 15 | 13 | - | - | - | - | 25 | - | 34 | |
| Horse | 6 | 7 | - | - | - | - | - | - | 3 | |
| Tolebi district, Turkestan region N 42°10′59″ E 69°52′57″ | Cattle | 15 | 64 | - | - | - | - | 82 | - | 32 |
| Sheep | 16 | 12 | - | - | - | - | 27 | - | 7 | |
| Horse | 5 | 8 | - | - | - | - | 35 | - | - | |
| Bokey Orda district, West Kazakhstan region N 48°57′22″ E 47°36′46″ | Cattle | 14 | 49 | - | - | - | 105 | - | 75 | 28 |
| Sheep | 20 | 23 | - | - | - | 56 | - | 27 | 46 | |
| Horse | 6 | 10 | - | - | - | 14 | - | - | 3 | |
| Zhanibek district, West Kazakhstan region N 49°27’00″ E 46°53’24″ | Cattle | 15 | 22 | - | - | - | 89 | - | 83 | 32 |
| Sheep | 20 | 21 | - | - | - | 47 | - | 25 | 25 | |
| Horse | 6 | - | - | - | - | 9 | - | - | - | |
| Korday district, Zhambyl region N 43°2′42″ E 74°42′27″ | Cattle | 12 | 92 | 21 | - | - | 38 | 34 | 12 | 37 |
| Sheep | 16 | 34 | - | - | - | 35 | 31 | - | 25 | |
| Horse | 5 | 4 | - | - | - | 12 | - | 2 | - | |
| Taraz city, Zhambyl region N 42°52′48″ E 71°21′47″ | Cattle | 24 | 78 | - | - | - | 67 | 22 | 34 | 19 |
| Sheep | 17 | 46 | - | - | - | 35 | 30 | 23 | 12 | |
| Horse | 6 | - | - | - | - | - | 5 | 2 | - | |
| Timiryazev district, North Kazakhstan region N 53°48′00″ E 66°32′24″ | Cattle | 12 | 103 | - | 40 | 4 | - | - | 30 | 27 |
| Sheep | 33 | 52 | - | 17 | - | - | - | 25 | 19 | |
| Horse | 7 | - | - | - | - | - | - | - | - | |
| Taiynsha district, North Kazakhstan region N 53°52′48″ E 69°43′48″ | Cattle | 10 | 95 | - | 15 | 7 | - | - | 40 | 35 |
| Sheep | 22 | 37 | - | - | - | - | - | 25 | 18 | |
| Horse | 5 | - | - | - | - | - | - | - | - | |
| Zhambyl district, Almaty region N 43°16′17″ E 76°40′15″ | Cattle | 15 | 34 | 28 | - | - | 56 | - | - | 28 |
| Sheep | 25 | - | 6 | - | - | 32 | - | - | 15 | |
| Horse | 6 | - | - | - | - | - | - | - | 9 | |
| Zhalagash district, Kyzylorda region N 45°4′49″ E 64°40′43″ | Cattle | 10 | - | - | 30 | 34 | 15 | 50 | 25 | |
| Sheep | 16 | - | - | - | 46 | 22 | 45 | 23 | ||
| Horse | 5 | - | - | - | - | - | - | |||
| Munaily district, Mangystau region N 43°41′44″ E 51°19′34” | Cattle | 15 | 10 | - | - | - | 15 | 23 | - | - |
| Sheep | 18 | - | - | - | - | - | 11 | - | - | |
| Horse | 6 | - | - | - | - | - | - | - | - | |
| Mangystau district, Mangystau region N 43°41′44″ E 51°19′34″ | Cattle | 10 | 12 | - | - | - | 13 | 14 | - | - |
| Sheep | 14 | - | - | - | - | 8 | - | - | - | |
| Horse | 7 | - | - | - | - | - | - | - | - | |
| Mugalzhar district, Aktobe region N 48°35′9″ E 58°27′44″ | Cattle | 10 | 34 | - | - | - | - | 12 | - | - |
| Sheep | 15 | 25 | - | - | - | - | - | - | - | |
| Horse | 6 | 6 | - | - | - | - | - | - | - | |
| 938 (28.08%) | 67 (2.01%) | 112 (3.35%) | 11 (0.33%) | 711 (21.28%) | 430 (12.87%) | 498 (14.90%) | 574 (17.18%) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sultankulova, K.T.; Shynybekova, G.O.; Issabek, A.U.; Mukhami, N.N.; Melisbek, A.M.; Chervyakova, O.V.; Kozhabergenov, N.S.; Barmak, S.M.; Bopi, A.K.; Omarova, Z.D.; et al. The Prevalence of Pathogens among Ticks Collected from Livestock in Kazakhstan. Pathogens 2022, 11, 1206. https://doi.org/10.3390/pathogens11101206
Sultankulova KT, Shynybekova GO, Issabek AU, Mukhami NN, Melisbek AM, Chervyakova OV, Kozhabergenov NS, Barmak SM, Bopi AK, Omarova ZD, et al. The Prevalence of Pathogens among Ticks Collected from Livestock in Kazakhstan. Pathogens. 2022; 11(10):1206. https://doi.org/10.3390/pathogens11101206
Chicago/Turabian StyleSultankulova, Kulyaisan T., Gaukhar O. Shynybekova, Aisha U. Issabek, Nazym N. Mukhami, Aibarys M. Melisbek, Olga V. Chervyakova, Nurlan S. Kozhabergenov, Sabyrkhan M. Barmak, Arailym K. Bopi, Zamira D. Omarova, and et al. 2022. "The Prevalence of Pathogens among Ticks Collected from Livestock in Kazakhstan" Pathogens 11, no. 10: 1206. https://doi.org/10.3390/pathogens11101206
APA StyleSultankulova, K. T., Shynybekova, G. O., Issabek, A. U., Mukhami, N. N., Melisbek, A. M., Chervyakova, O. V., Kozhabergenov, N. S., Barmak, S. M., Bopi, A. K., Omarova, Z. D., Alibekova, D. A., Argimbayeva, T. U., Namet, A. M., Zuban, I. A., & Orynbayev, M. B. (2022). The Prevalence of Pathogens among Ticks Collected from Livestock in Kazakhstan. Pathogens, 11(10), 1206. https://doi.org/10.3390/pathogens11101206