
Prevalence of Human Intestinal Entamoeba spp. in the Americas: A Systematic Review and Meta-Analysis, 1990–2022

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. The Review Question
2.2. Literature Research
2.3. Data Summary
2.4. GenBank Sequences
3. Results
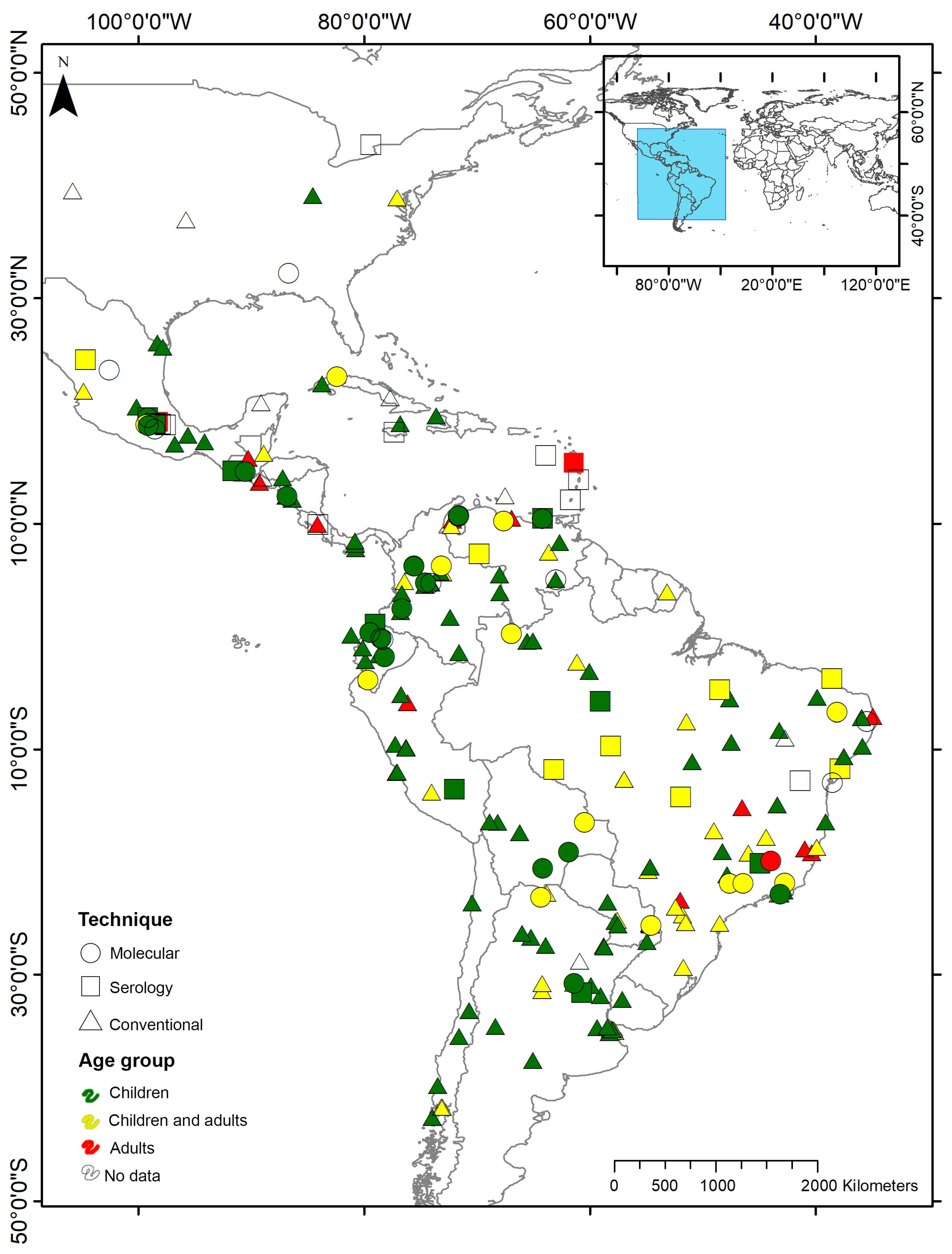
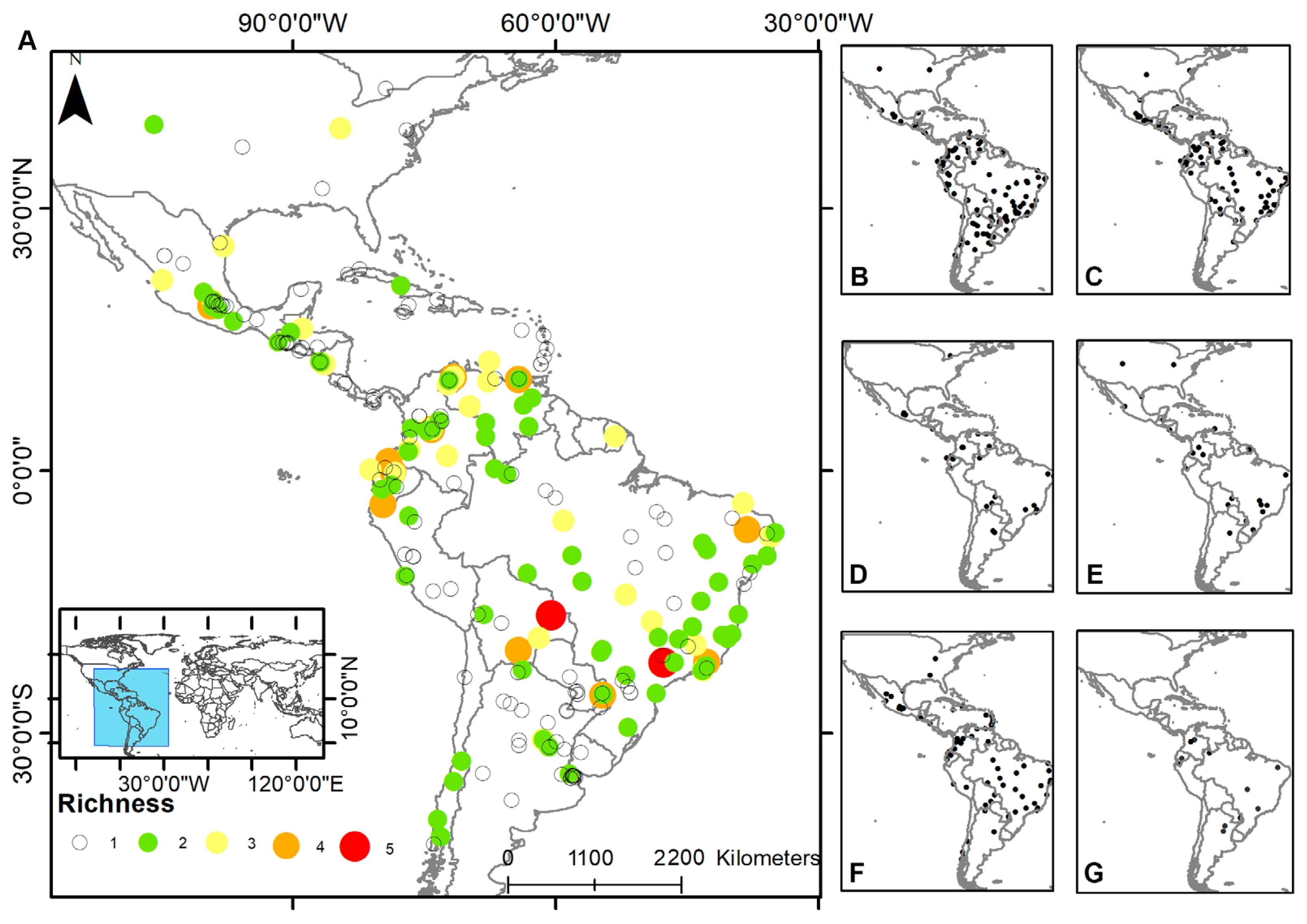
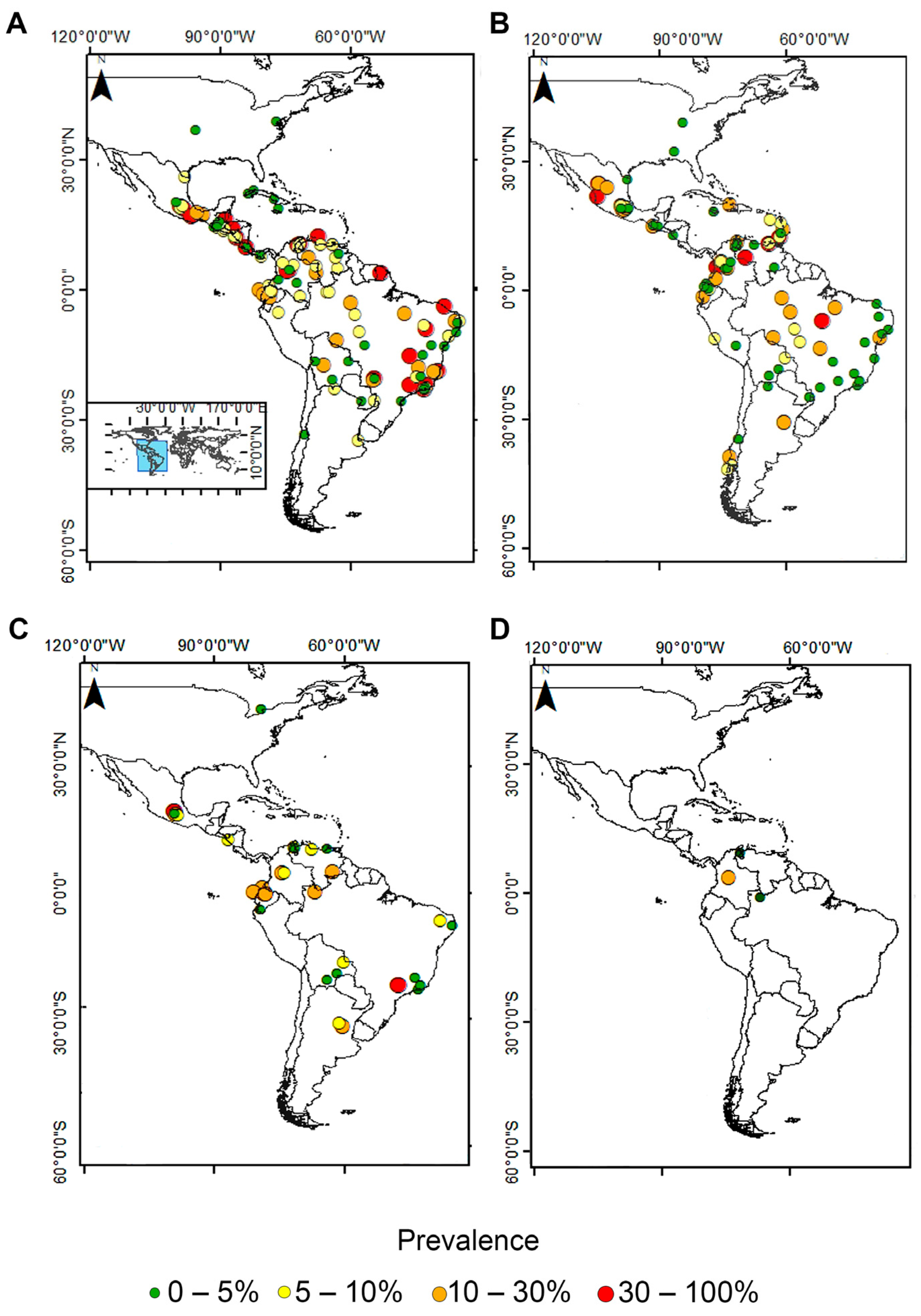
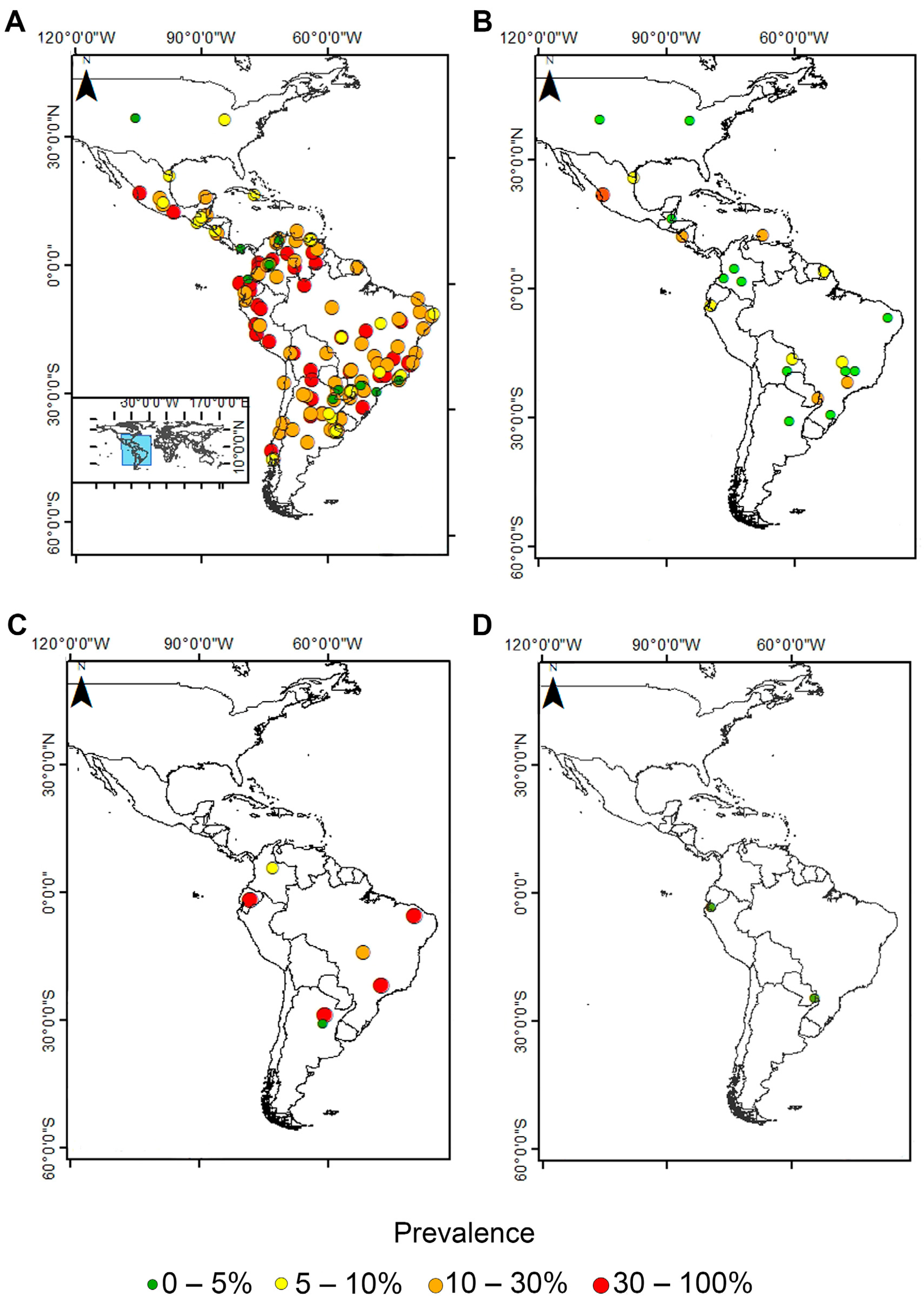
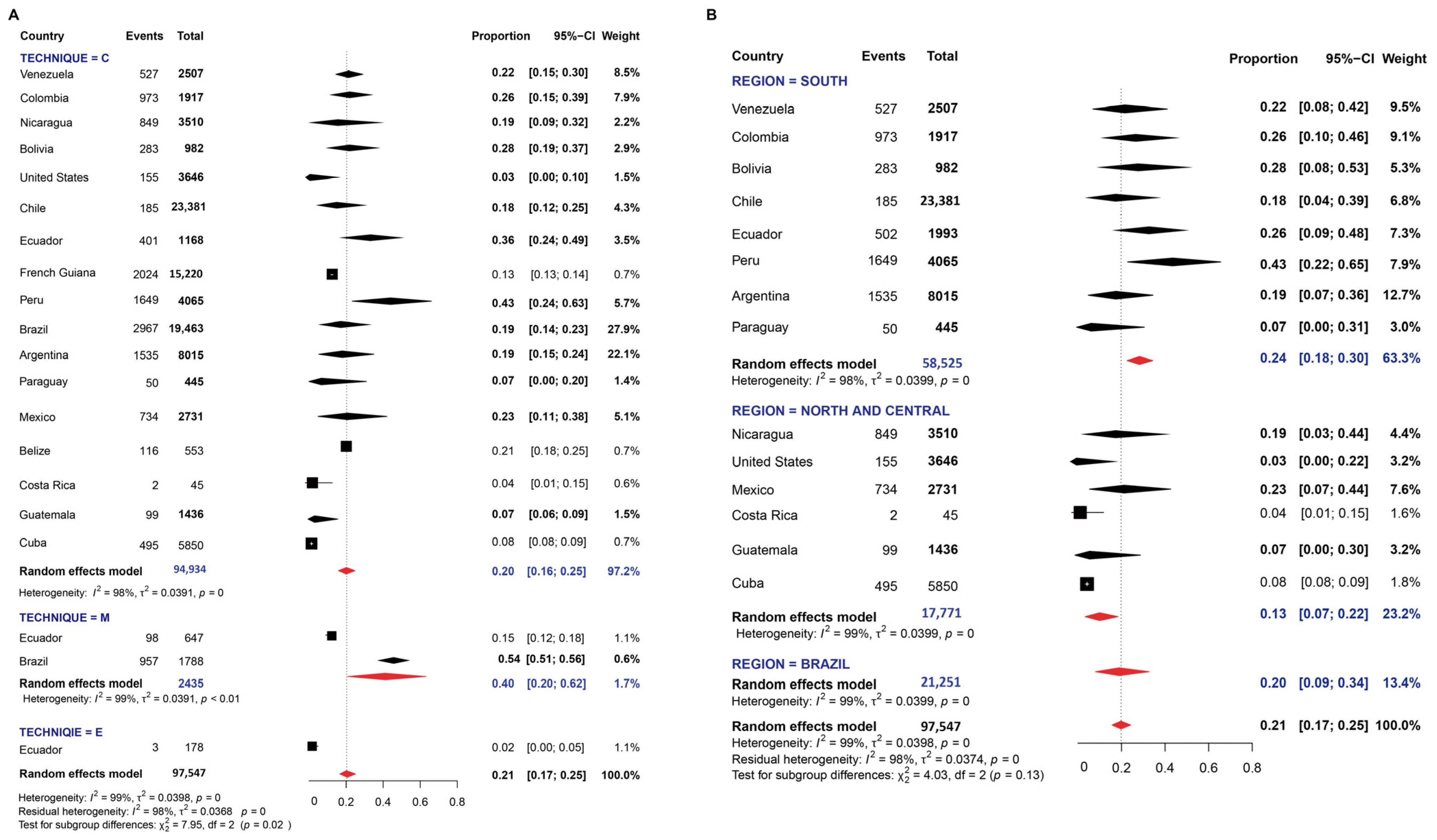
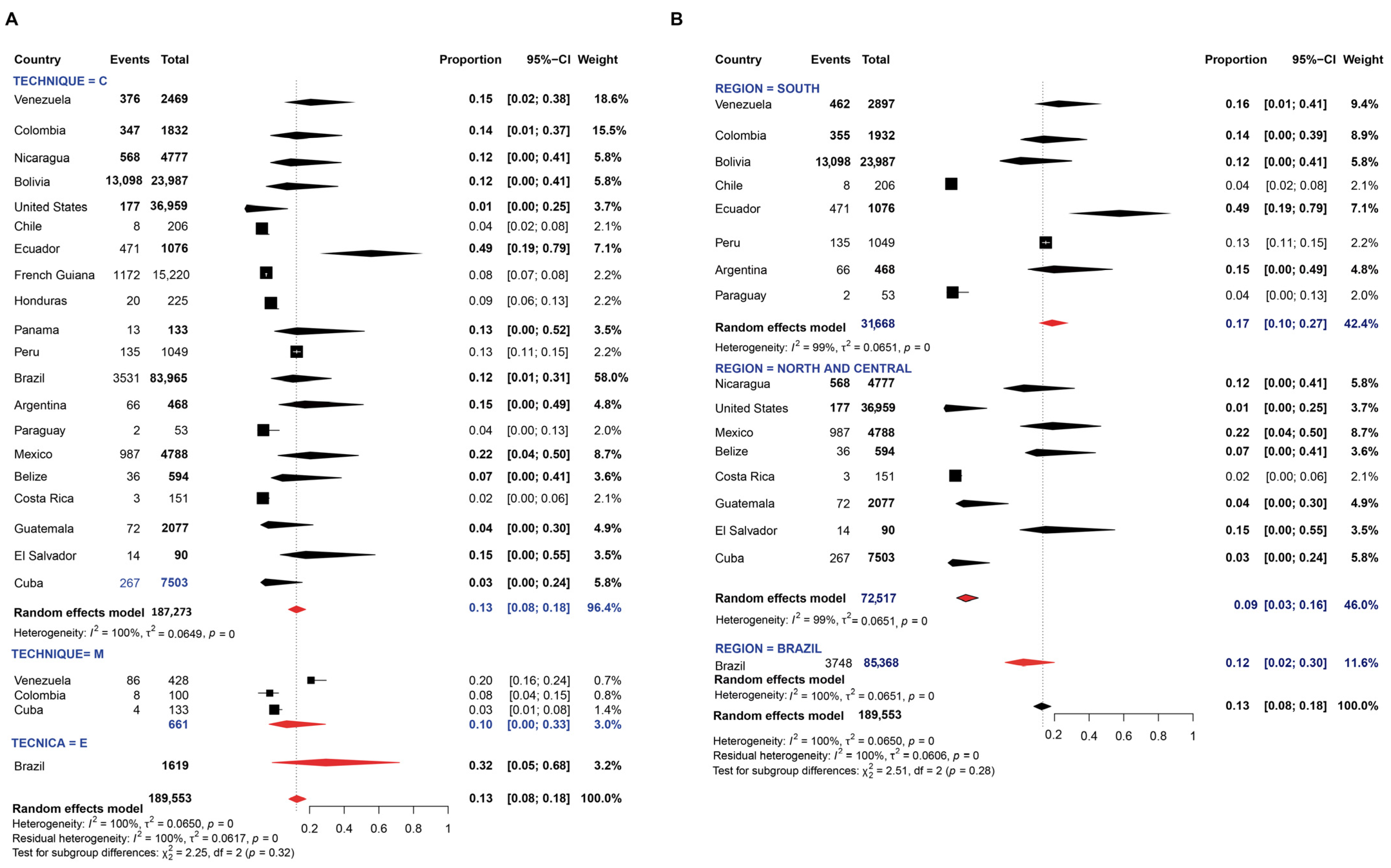
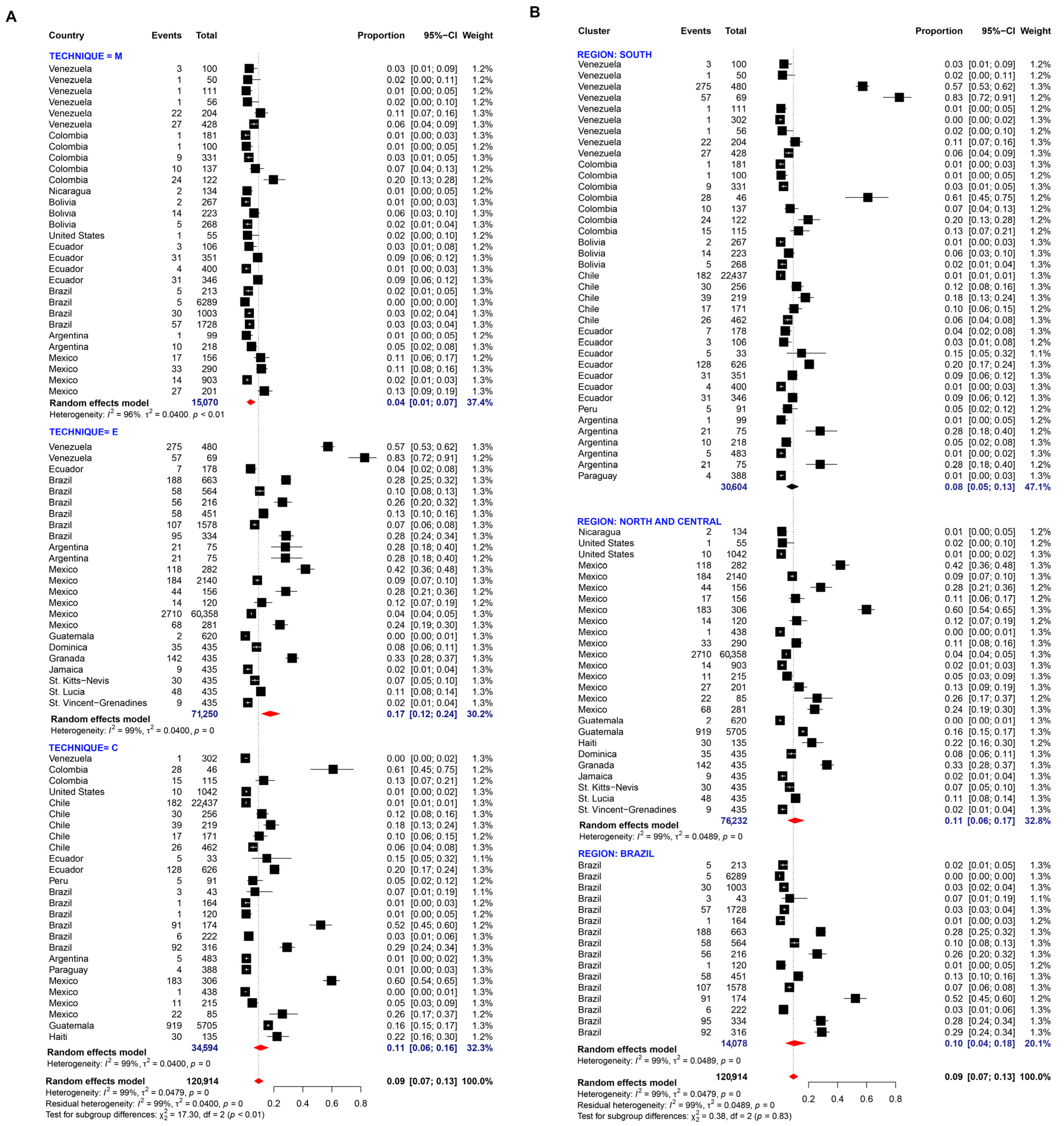
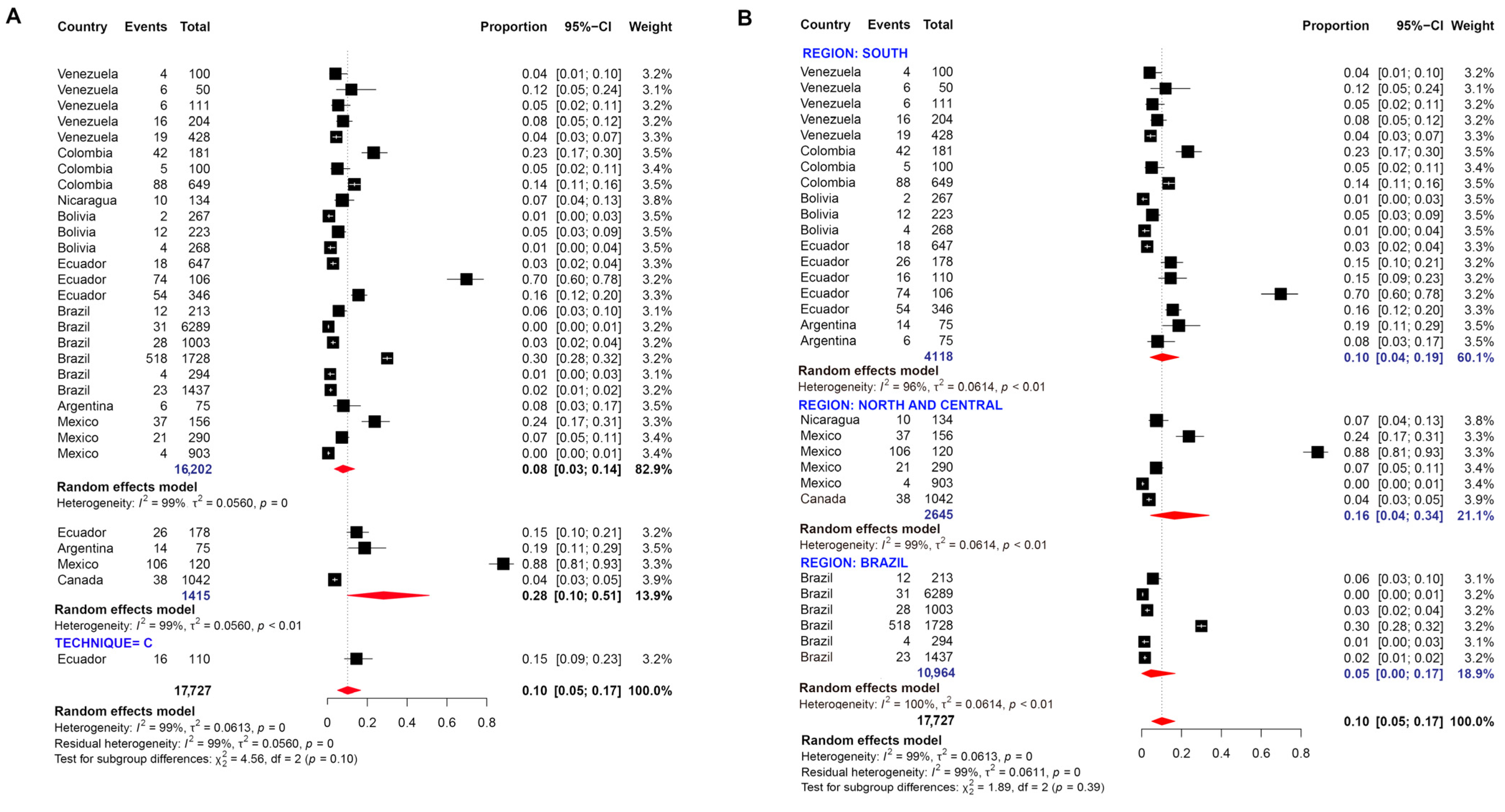
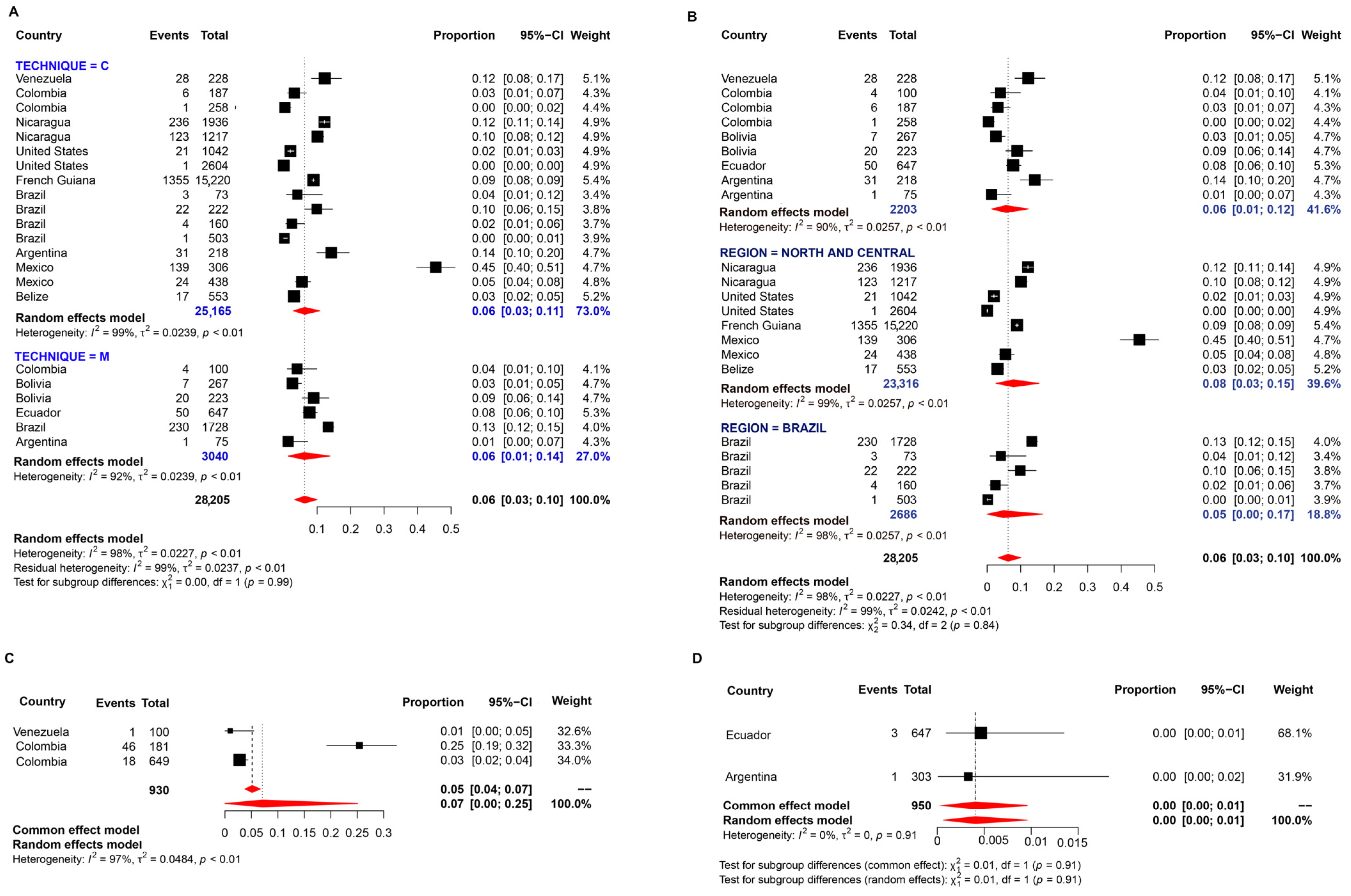
3.1. Pooled Prevalence of Entamoeba Infection
3.2. GenBank Sequences
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stensvold, C.R.; Lebbad, M.; Victory, E.L.; Verweij, J.J.; Tannich, E.; Alfellani, M.; Legarraga, P.; Clark, C.G. Increased sampling reveals novel lineages of Entamoeba: Consequences of genetic diversity and host specificity for taxonomy and molecular detection. Protist 2011, 162, 525–541. [Google Scholar] [CrossRef] [Green Version]
- Ali, I.K. Intestinal amebae. Clin. Lab. Med. 2015, 35, 393–422. [Google Scholar] [CrossRef] [PubMed]
- Heredia, R.D.; Fonseca, J.A.; Lopez, M.C. Entamoeba moshkovskii perspectives of a new agent to be considered in the diagnosis of amebiasis. Acta Trop. 2012, 123, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa, C.; Kabir, M.; Taniuchi, M.; Mondal, D.; Kobayashi, S.; Ali, I.K.; Sobuz, S.U.; Senba, M.; Houpt, E.; Haque, R.; et al. Entamoeba moshkovskii is associated with diarrhea in infants and causes diarrhea and colitis in mice. J. Infect. Dis. 2012, 206, 744–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royer, T.L.; Gilchrist, C.; Kabir, M.; Arju, T.; Ralston, K.S.; Haque, R.; Clark, C.G.; Petri, W.A., Jr. Entamoeba bangladeshi nov. sp., Bangladesh. Emerg. Infect. Dis. 2012, 18, 1543–1545. [Google Scholar] [CrossRef]
- Oliveira, F.M.; Neumann, E.; Gomes, M.A.; Caliari, M.V. Entamoeba dispar: Could it be pathogenic. Trop. Parasitol. 2015, 5, 9–14. [Google Scholar]
- Stensvold, C.R.; Winiecka-Krusnell, J.; Lier, T.; Lebbad, M. Evaluation of a PCR method for detection of Entamoeba polecki, with an overview of its molecular epidemiology. J. Clin. Microbiol. 2018, 56, e00154-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levecke, B.; Dorny, P.; Vercammen, F.; Visser, L.G.; Van Esbroeck, M.; Vercruysse, J.; Verweij, J.J. Transmission of Entamoeba nuttalli and Trichuris trichiura from nonhuman primates to humans. Emerg. Infect. Dis. 2015, 21, 1871–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uribe-Querol, E.; Rosales, C. Immune response to the enteric parasite Entamoeba histolytica. Physiology 2020, 35, 244–260. [Google Scholar] [CrossRef]
- Hotez, P.J. Could nitazoxanide be added to other essential medicines for integrated neglected tropical disease control and elimination? PLoS Negl. Trop Dis. 2014, 8, e2758. [Google Scholar] [CrossRef] [Green Version]
- Turkeltaub, J.A.; McCarty, T.R., 3rd; Hotez, P.J. The intestinal protozoa: Emerging impact on global health and development. Curr. Opin. Gastroenterol. 2015, 31, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Kantor, M.; Abrantes, A.; Estevez, A.; Schiller, A.; Torrent, J.; Gascon, J.; Hernandez, R.; Ochner, C. Entamoeba Histolytica: Updates in clinical manifestation, pathogenesis, and vaccine development. Can. J. Gastroenterol. Hepatol. 2018, 2018, 4601420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowak, P.; Mastalska, K.; Loster, J. Entamoeba histolytica-pathogenic protozoan of the large intestine in humans. J. Clin. Microbiol. Biochem. Technol. 2015, 1, 17. [Google Scholar] [CrossRef] [Green Version]
- Ngobeni, R.; Samie, A.; Moonah, S.; Watanabe, K.; Petri, W.A., Jr.; Gilchrist, C. Entamoeba species in South Africa: Correlations with the host microbiome, parasite burdens, and first description of Entamoeba bangladeshi outside of Asia. J. Infect. Dis. 2017, 216, 1592–1600. [Google Scholar] [CrossRef]
- Ten Hove, R.J.; van Esbroeck, M.; Vervoort, T.; van den Ende, J.; van Lieshout, L.; Verweij, J.J. Molecular diagnostics of intestinal parasites in returning travellers. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 1045–1053. [Google Scholar] [CrossRef] [Green Version]
- Herbinger, K.H.; Fleischmann, E.; Weber, C.; Perona, P.; Loscher, T.; Bretzel, G. Epidemiological, clinical, and diagnostic data on intestinal infections with Entamoeba histolytica and Entamoeba dispar among returning travelers. Infection 2011, 39, 527–535. [Google Scholar] [CrossRef]
- Calegar, D.A.; Nunes, B.C.; Monteiro, K.J.; Santos, J.P.; Toma, H.K.; Gomes, T.F.; Lima, M.M.; Boia, M.N.; Carvalho-Costa, F.A. Frequency and molecular characterisation of Entamoeba histolytica, Entamoeba dispar, Entamoeba moshkovskii, and Entamoeba hartmanni in the context of water scarcity in northeastern Brazil. Mem. Inst. Oswaldo Cruz 2016, 111, 114–119. [Google Scholar] [CrossRef]
- Blessmann, J.; Van Linh, P.; Nu, P.A.; Thi, H.D.; Muller-Myhsok, B.; Buss, H.; Tannich, E. Epidemiology of amebiasis in a region of high incidence of amebic liver abscess in central Vietnam. Am. J. Trop. Med. Hyg. 2002, 66, 578–583. [Google Scholar] [CrossRef] [Green Version]
- Haque, R.; Kress, K.; Wood, S.; Jackson, T.F.H.G.; Lyerly, D.; Wilkins, T.; Petri, W.A. Diagnosis of pathogenic Entamoeba histolytica infection using a stool ELISA based on monoclonal antibodies to the galactose-specific adhesin. J. Infect. Dis. 1993, 167, 247–249. [Google Scholar] [CrossRef]
- Fotedar, R.; Stark, D.; Beebe, N.; Marriott, D.; Ellis, J.; Harkness, J. Laboratory diagnostic techniques for Entamoeba species. Clin. Microbiol. Rev. 2007, 20, 511–532. [Google Scholar] [CrossRef] [Green Version]
- Bahrami, F.; Haghighi, A.; Zamini, G.; Khademerfan, M. Differential detection of Entamoeba histolytica, Entamoeba dispar and Entamoeba moshkovskii in faecal samples using nested multiplex PCR in west of Iran. Epidemiol. Infect. 2019, 147, e96. [Google Scholar] [CrossRef] [PubMed]
- Ximénez, C.; Moran, P.; Rojas, L.; Valadez, A.; Gomez, A. Reassessment of the epidemiology of amebiasis: State of the art. Infect. Genet. Evol. 2009, 9, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- Nair, G.; Rebolledo, M.; White, A.C., Jr.; Crannell, Z.; Richards-Kortum, R.R.; Pinilla, A.E.; Ramirez, J.D.; Lopez, M.C.; Castellanos-Gonzalez, A. Detection of Entamoeba histolytica by recombinase polymerase amplification. Am. J. Trop. Med. Hyg. 2015, 93, 591–595. [Google Scholar] [CrossRef]
- Acuna-Soto, R.; Samuelson, J.; De Girolami, P.; Zarate, L.; Millan-Velasco, F.; Schoolnick, G.; Wirth, D. Application of the polymerase chain reaction to the epidemiology of pathogenic and nonpathogenic Entamoeba histolytica. Am. J. Trop. Med. Hyg. 1993, 48, 58–70. [Google Scholar] [CrossRef]
- Sanchez-Guillen Mdel, C.; Perez-Fuentes, R.; Salgado-Rosas, H.; Ruiz-Arguelles, A.; Ackers, J.; Shire, A.; Talamas-Rohana, P. Differentiation of Entamoeba histolytica/Entamoeba dispar by PCR and their correlation with humoral and cellular immunity in individuals with clinical variants of amoebiasis. Am. J. Trop. Med. Hyg. 2002, 66, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Ramos, F.; Garcia, G.; Valadez, A.; Moran, P.; Gonzalez, E.; Gomez, A.; Melendro, E.I.; Valenzuela, O.; Ximenez, C.E. dispar strain: Analysis of polymorphism as a tool for study of geographic distribution. Mol. Biochem. Parasitol. 2005, 141, 175–177. [Google Scholar] [CrossRef]
- Rojas, L.; Moran, P.; Valadez, A.; Gomez, A.; Gonzalez, E.; Hernandez, E.; Partida, O.; Nieves, M.; Gudino, M.; Magana, U.; et al. Entamoeba histolytica and Entamoeba dispar infection in Mexican school children: Genotyping and phylogenetic relationship. BMC Infect. Dis. 2016, 16, 485. [Google Scholar] [CrossRef] [Green Version]
- Gunther, J.; Shafir, S.; Bristow, B.; Sorvillo, F. Short report: Amebiasis-related mortality among United States residents, 1990–2007. Am. J. Trop. Med. Hyg. 2011, 85, 1038–1040. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 3 September 2022).
- Schwarzer, G. meta: An R package for meta-analysis. R News 2007, 7, 40–45. [Google Scholar]
- Higgins, J.P.; Thompson, S.G.; Deeks, J.J.; Altman, D.G. Measuring inconsistency in meta-analyses. BMJ 2003, 327, 557–560. [Google Scholar] [CrossRef] [Green Version]
- Quantum GIS Development Team. QGIS geographic information system. Open Source Geospatial Foundation Project. 2020. Available online: https://qgis.org/es/site/ (accessed on 21 September 2022).
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zonta, M.L.; Servián, A.; Panisse, G.; Oyhenart, E.E.; Navone, G.T. Nutritional status, intestinal parasitic infections, and socio-environmental conditions in Mbyá-Guaraní children: The current situation in communities in Central Misiones, Argentina. Am. J. Hum. Biol. 2022, 34, e23749. [Google Scholar] [CrossRef] [PubMed]
- López Arias, L.; Guillemi, E.; Bordoni, N.; Farber, M.; Garbossa, G. Diagnóstico de Entamoeba polecki y su potencial impacto en las condiciones sanitarias de la producción porcina. Rev. Investig. Agropecu. 2019, 45, 373–377. [Google Scholar]
- Calegar, D.A.; Monteiro, K.J.L.; Bacelar, P.A.A.; Evangelista, B.B.C.; Almeida, M.M.; dos Santos, J.P.; Boia, M.N.; Coronato-Nunes, B.; Jaeger, L.H.; Carvalho-Costa, F.A. Epidemiology, species composition and genetic diversity of tetra- and octonucleated Entamoeba spp. in different Brazilian biomes. Parasites Vectors 2021, 14, 160. [Google Scholar] [CrossRef]
- Ximénez, C.; Cerritos, R.; Rojas, L.; Dolabella, S.; Morán, P.; Shibayama, M.; González, E.; Valadez, A.; Hernández, E.; Valenzuela, O.; et al. Human amebiasis: Breaking the paradigm? Int. J. Environ. Res. Public Health 2010, 7, 1105–1120. [Google Scholar] [CrossRef] [PubMed]
- Tawari, B.; Ali, I.K.M.; Scott, C.; Quail, M.A.; Berriman, M.; Hall, N.; Clark, C.G. Patterns of evolution in the unique tRNA gene arrays of the genus entamoeba. Mol. Biol. Evol. 2007, 25, 187–198. [Google Scholar] [CrossRef] [Green Version]
- Lopez, M.C.; Leon, C.M.; Fonseca, J.; Reyes, P.; Moncada, L.; Olivera, M.J.; Ramirez, J.D. Molecular epidemiology of Entamoeba: First description of Entamoeba moshkovskii in a rural area from Central Colombia. PLoS ONE 2015, 10, e0140302. [Google Scholar] [CrossRef] [Green Version]
- Morán, P.; González, E.; Portillo, T.; Reyes, M.Á.; Hernandez, E.; Ramiro, M.; Nieves, M.; Partida, O.; Cerritos, R.; Gudiño, M.; et al. Cutaneous amebiasis: The importance of molecular diagnosis of an emerging parasitic disease. Am. J. Trop. Med. Hyg. 2013, 88, 186–190. [Google Scholar] [CrossRef] [Green Version]
- Kebede, A.; Verweij, J.J.; Petros, B.; Polderman, A.M. Short communication: Misleading microscopy in amoebiasis. Trop. Med. Int. Health 2004, 9, 651–652. [Google Scholar] [CrossRef]
- Pinheiro, S.M.; Carneiro, R.M.; Aca, I.S.; Irmao, J.I.; Morais, M.A., Jr.; Coimbra, M.R.; Carvalho, L.B., Jr. Determination of the prevalence of Entamoeba histolytica and E. dispar in the pernambuco state of northeastern Brazil by a polymerase chain reaction. Am. J. Trop. Med. Hyg. 2004, 70, 221–224. [Google Scholar] [CrossRef]
- Haque, R.; Ali, I.K.; Akther, S.; Petri, W.A., Jr. Comparison of PCR, isoenzyme analysis, and antigen detection for diagnosis of Entamoeba histolytica infection. J. Clin. Microbiol. 1998, 36, 449–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. WHO/PAHO/UNESCO report. A consultation with experts on amoebiasis. Mexico City, Mexico 28–29 January, 1997. Epidemiol. Bull. 1997, 18, 13–14. [Google Scholar]
- Cui, Z.; Li, J.; Chen, Y.; Zhang, L. Molecular epidemiology, evolution, and phylogeny of Entamoeba spp. Infect. Genet. Evol. 2019, 75, 104018. [Google Scholar] [CrossRef] [PubMed]
- Nesbitt, R.A.; Mosha, F.W.; Katki, H.A.; Ashraf, M.; Assenga, C.; Lee, C.M. Amebiasis and comparison of microscopy to ELISA technique in detection of Entamoeba histolytica and Entamoeba dispar. J. Natl. Med. Assoc. 2004, 96, 671–677. [Google Scholar]
- Ngui, R.; Angal, L.; Fakhrurrazi, S.A.; Lian, Y.L.; Ling, L.Y.; Ibrahim, J.; Mahmud, R. Differentiating Entamoeba histolytica, Entamoeba dispar and Entamoeba moshkovskii using nested polymerase chain reaction (PCR) in rural communities in Malaysia. Parasites Vectors 2012, 5, 187. [Google Scholar] [CrossRef]
- Morsy, S.M.; Elmatrawy, O.M.; Rubio, J.M.; El-Badry, A.A.; Hassan, M.A. Enteric pathogenic protozoa from misdiagnosis to overmedication in Egypt: A need for molecular diagnosis. Comp. Clin. Pathol. 2022, 5, 765–772. [Google Scholar] [CrossRef]
- Leiva, B.; Lebbad, M.; Winiecka-Krusnell, J.; Altamirano, I.; Tellez, A.; Linder, E. Overdiagnosis of Entamoeba histolytica and Entamoeba dispar in Nicaragua: A microscopic, triage parasite panel and PCR study. Arch. Med. Res. 2006, 37, 529–534. [Google Scholar] [CrossRef]
- Shnawa, B. Molecular diagnosis of Entamoeba histolytica, Entamoeba dispar, and Entamoeba moshkovskii: An update review. Annu. Res. Rev. Biol. 2017, 21, 1–12. [Google Scholar] [CrossRef]
- La Hoz, R.M.; Morris, M.I.; AST Infectious Diseases Community of Practice. Intestinal parasites including Cryptosporidium, Cyclospora, Giardia, and Microsporidia, Entamoeba histolytica, Strongyloides, Schistosomiasis, and Echinococcus: Guidelines from the American Society of Transplantation Infectious Diseases Community of Practice. Clin. Transpl. 2019, 33, e13618. [Google Scholar]
- Yanagawa, Y.; Shimogawara, R.; Endo, T.; Fukushima, R.; Gatanaga, H.; Hayasaka, K.; Kikuchi, Y.; Kobayashi, T.; Koga, M.; Koibuchi, T.; et al. Utility of the rapid antigen detection test, E. histolytica quik chek for the diagnosis of Entamoeba histolytica infection in nonendemic situations. J. Clin. Microbiol. 2020, 58, e01991-20. [Google Scholar] [CrossRef]
- Dos Santos Zanetti, A.; Malheiros, A.F.; de Matos, T.A.; Dos Santos, C.; Battaglini, P.F.; Moreira, L.M.; Lemos, L.M.S.; Castrillon, S.K.I.; da Costa Boamorte Cortela, D.; Ignotti, E.; et al. Diversity, geographical distribution, and prevalence of Entamoeba spp. in Brazil: A systematic review and meta-analysis. Parasite 2021, 28, 17. [Google Scholar] [CrossRef] [PubMed]
- Villamizar, X.; Higuera, A.; Herrera, G.; Vasquez, A.L.R.; Buitron, L.; Muñoz, L.M.; Gonzalez, C.F.E.; Lopez, M.C.; Giraldo, J.C.; Ramírez, J.D. Molecular and descriptive epidemiology of intestinal protozoan parasites of children and their pets in Cauca, Colombia: A cross-sectional study. BMC Infect. Dis. 2019, 19, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreto, S.M.; Miranda, J.J.; Figueroa, J.P.; Schmidt, M.I.; Munoz, S.; Kuri-Morales, P.P.; Silva, J.B., Jr. Epidemiology in Latin America and the Caribbean: Current situation and challenges. Int. J. Epidemiol. 2012, 41, 557–571. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecular Marker | Species | Accession No. | Country | Host | Isolation Source | References |
|---|---|---|---|---|---|---|
| SSU rRNA gene | Entamoeba dispar | MK541026 | Argentina | Homo sapiens | Stool | Direct submission |
| Entamoeba dispar | OM985615 | Argentina | Homo sapiens | Stool | Direct submission | |
| Entamoeba dispar | MZ787761 | Argentina | Homo sapiens | Stool | Direct submission | |
| Entamoeba dispar | OM985618 | Argentina | Homo sapiens | Stool | Direct submission | |
| Entamoeba coli | ON713469 | Argentina | Homo sapiens | Stool | Direct submission | |
| Entamoeba coli | OM985619 | Argentina | Homo sapiens | Stool | Direct submission | |
| Entamoeba coli | MZ787759 | Argentina | Homo sapiens | Stool | Direct submission | |
| Entamoeba coli | OM985620 | Argentina | Homo sapiens | Stool | Direct submission | |
| Entamoeba coli | MZ787760 | Argentina | Homo sapiens | Stool | Direct submission | |
| Entamoeba coli | OM985617 | Argentina | Homo sapiens | Stool | Direct submission | |
| Entamoeba coli | MK541024 | Argentina | Homo sapiens | Stool | Direct submission | |
| Entamoeba coli | OM985616 | Argentina | Homo sapiens | Stool | Direct submission | |
| Entamoeba coli | OM985619 | Argentina | Homo sapiens | Stool | Direct submission | |
| Entamoeba hartmanni | MT703882 | Argentina | Homo sapiens | Stool | [34] | |
| Entamoeba polecki | MH348163-MH348175 | Argentina | Sus scrofa domestica | Stool | [35] | |
| SSU rRNA gene | Entamoeba dispar | MW026767-MW026784 | Brazil | Homo sapiens | Stool | [36] |
| Entamoeba histolytica | MW026793 MW026794 | Brazil | Homo sapiens | Stool | [36] | |
| Entamoeba hartmanni | MW026785-MW026792 | Brazil | Homo sapiens | Stool | [36] | |
| Entamoeba coli | MW026735-MW026766 | Brazil | Homo sapiens | Stool | [36] | |
| Entamoeba coli | FR686423 | Brazil | Homo sapiens | Stool | [1] | |
| tRNA | Entamoeba dispar | GU324326 | Brazil | Homo sapiens | Stool | [37] |
| Entamoeba histolytica | EF421375 | Brazil | Homo sapiens | Stool | [38] | |
| SSU rRNA gene | Entamoeba histolytica | KT825974 | Colombia | Homo sapiens | Stool | [39] |
| Entamoeba moshkovskii | KT825984-KT825993 | Colombia | Homo sapiens | Stool | [39] | |
| Entamoeba dispar | KT825975-KT825983 | Colombia | Homo sapiens | Stool | [39] | |
| SSU rRNA gene | Entamoeba coli | FR686443 | Peru | Homo sapiens | Stool | [1] |
| SSU rRNA gene | Entamoeba coli | FR686446 | Ecuador | Homo sapiens | Stool | [1] |
| SSU rRNA gene | Entamoeba hartmanni | MK541027 | Mexico | Homo sapiens | Stool | Direct submission |
| Entamoeba coli | MK541025 | Mexico | Homo sapiens | Stool | Direct submission | |
| tRNA | Entamoeba dispar | GU324327-GU324329 GU324333- GU324337 | Mexico | Homo sapiens | Mixed liver abscess | [37] |
| Entamoeba histolytica | GU324330-GU324332 | Mexico | Homo sapiens | Mixed liver abscess | [37] | |
| Entamoeba histolytica | JN191598 JN191599 JQ828978 | Mexico | Homo sapiens | Cutaneous amoebiasis | [40] | |
| Entamoeba histolytica | KC791705-KC791758 | Mexico | Homo sapiens | Amoebic liver abscess | [40] | |
| Entamoeba dispar | KX461938-KX461956 | Mexico | Homo sapiens | Stool | [27] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Servián, A.; Helman, E.; Iglesias, M.d.R.; Panti-May, J.A.; Zonta, M.L.; Navone, G.T. Prevalence of Human Intestinal Entamoeba spp. in the Americas: A Systematic Review and Meta-Analysis, 1990–2022. Pathogens 2022, 11, 1365. https://doi.org/10.3390/pathogens11111365
Servián A, Helman E, Iglesias MdR, Panti-May JA, Zonta ML, Navone GT. Prevalence of Human Intestinal Entamoeba spp. in the Americas: A Systematic Review and Meta-Analysis, 1990–2022. Pathogens. 2022; 11(11):1365. https://doi.org/10.3390/pathogens11111365
Chicago/Turabian StyleServián, Andrea, Elisa Helman, María del Rosario Iglesias, Jesús Alonso Panti-May, María Lorena Zonta, and Graciela Teresa Navone. 2022. "Prevalence of Human Intestinal Entamoeba spp. in the Americas: A Systematic Review and Meta-Analysis, 1990–2022" Pathogens 11, no. 11: 1365. https://doi.org/10.3390/pathogens11111365
APA StyleServián, A., Helman, E., Iglesias, M. d. R., Panti-May, J. A., Zonta, M. L., & Navone, G. T. (2022). Prevalence of Human Intestinal Entamoeba spp. in the Americas: A Systematic Review and Meta-Analysis, 1990–2022. Pathogens, 11(11), 1365. https://doi.org/10.3390/pathogens11111365