
Old and New Insights into Sporothrix schenckii Complex Biology and Identification




{kind=link}
Abstract
:1. Introduction
2. Species-Level Identification
2.1. Microscopy, Culture, and Immunology-Based Methods
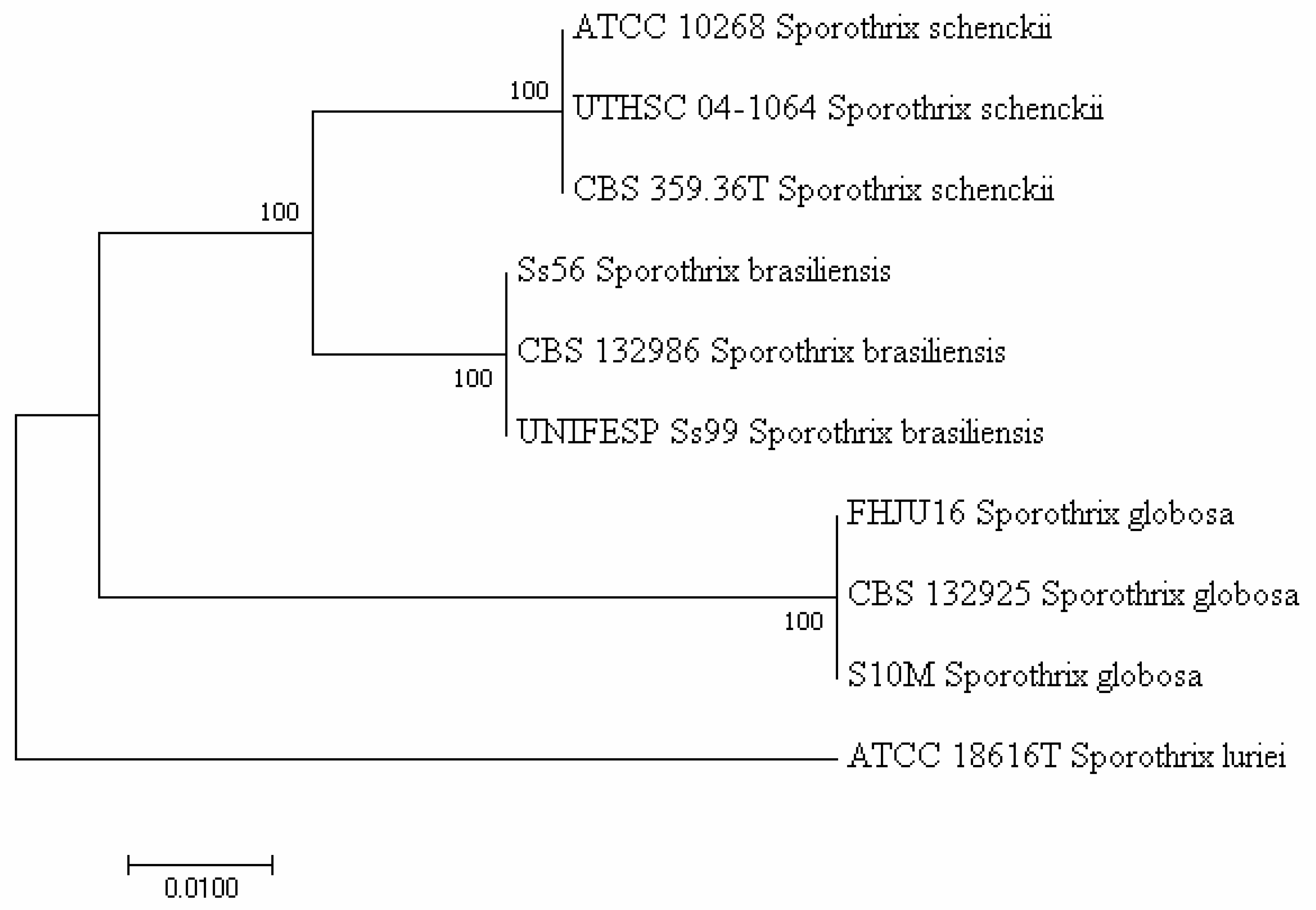
2.2. Molecular Biology-Based Methods
2.3. Matrix-Assisted Laser Desorption Ionization Time-of-Flight-Based Methods
3. Genome and Dimorphism
4. Cell Wall and Surface Components
5. Biofilm and Antifungal Resistance
6. Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- De Lima Barros, M.B.; de Almeida Paes, R.; Schubach, A.O. Sporothrix schenckii and sporotrichosis. Clin. Microbiol. Rev. 2011, 24, 633–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarro, J.; Gené, J.; Stchigel, A.M. Developments in fungal taxonomy. Clin. Microbiol. Rev. 1999, 12, 454–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marimon, R.; Gené, J.; Cano, J.; Trilles, L.; Dos Santos Lazéra, M.; Guarro, J. Molecular phylogeny of Sporothrix schenckii. J. Clin. Microbiol. 2006, 44, 3251–3256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marimon, R.; Cano, J.; Gené, J.; Sutton, D.A.; Kawasaki, M.; Guarro, J. Sporothrix brasiliensis, S. globosa, and S. mexicana, three new Sporothrix species of clinical interest. J. Clin. Microbiol. 2007, 45, 3198–3206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakrabarti, A.; Bonifaz, A.; Gutierrez-Galhardo, M.C.; Mochizuki, T.; Li, S. Global epidemiology of sporotrichosis. Med. Mycol. 2015, 53, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Álvarez, J.A.; Pérez-García, L.A.; Mellado-Mojica, E.; López, M.G.; Martínez-Duncker, I.; Lópes-Bezerra, L.M.; Mora-Montes, H.M. Sporothrix schenckii sensu stricto and Sporothrix brasiliensis are differentially recognized by human peripheral blood mononuclear cells. Front. Microbiol. 2017, 8, 843. [Google Scholar] [CrossRef]
- Arrillaga-Moncrieff, I.; Capilla, J.; Mayayo, E.; Marimon, R.; Mariné, M.; Gené, J.; Cano, J.; Guarro, J. Different virulence levels of the species of Sporothrix in a murine model. Clin. Microbiol. Infect. 2009, 15, 651–655. [Google Scholar] [CrossRef] [Green Version]
- Borba-Santos, L.P.; Rodrigues, A.M.; Gagini, T.B.; Fernandes, G.F.; Castro, R.; de Camargo, Z.P.; Nucci, M.; Lopes-Bezerra, L.M.; Ishida, K.; Rozental, S. Susceptibility of Sporothrix brasiliensis isolates to amphotericin B, azoles, and terbinafine. Med. Mycol. 2015, 53, 178–188. [Google Scholar] [CrossRef]
- Bonifaz, A.; Tirado-Sánchez, A. Cutaneous disseminated and extracutaneous sporotrichosis: Current status of a complex disease. J. Fungi 2017, 3, 6. [Google Scholar] [CrossRef] [Green Version]
- Queiroz-Telles, F.; Buccheri, R.; Benard, G. Sporotrichosis in immunocompromised hosts. J. Fungi 2019, 5, 8. [Google Scholar] [CrossRef] [Green Version]
- Do Monte Alves, M.; Pipolo Milan, E.; da Silva-Rocha, W.P.; Soares de Sena da Costa, A.; Araújo Maciel, B.; Cavalcante Vale, P.H.; de Albuquerque, P.R.; Lopes Lima, S.; Salles de Azevedo Melo, A.; Messias Rodrigues, A.; et al. Fatal pulmonary sporotrichosis caused by Sporothrix brasiliensis in Northeast Brazil. PLoS Negl. Trop. Dis. 2020, 14, e0008141. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Bento, A.; de Sena Costa, A.S.; Lima, S.L.; do Monte Alves, M.; de Azevedo Melo, A.S.; Rodrigues, A.M.; da Silva-Rocha, W.P.; Milan, E.P.; Chaves, G.M. The spread of cat-transmitted sporotrichosis due to Sporothrix brasiliensis in Brazil towards the Northeast region. PLoS Negl. Trop. Dis. 2021, 15, e0009693. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.M.; de Melo Teixeira, M.; de Hoog, G.S.; Schubach, T.M.; Pereira, S.A.; Fernandes, G.F.; Bezerra, L.M.; Felipe, M.S.; de Camargo, Z.P. Phylogenetic analysis reveals a high prevalence of Sporothrix brasiliensis in feline sporotrichosis outbreaks. PLoS Negl. Trop. Dis. 2013, 7, e2281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, M.M.; de Almeida, L.G.; Kubitschek-Barreira, P.; Alves, F.L.; Kioshima, E.S.; Abadio, A.K.; Fernandes, L.; Derengowski, L.S.; Ferreira, K.S.; Souza, R.C.; et al. Comparative genomics of the major fungal agents of human and animal sporotrichosis: Sporothrix schenckii and Sporothrix brasiliensis. BMC Genom. 2014, 29, 943. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Hagen, F.; Stielow, B.; Rodrigues, A.M.; Samerpitak, K.; Zhou, X.; Feng, P.; Yang, L.; Chen, M.; Deng, S.; et al. Phylogeography and evolutionary patterns in Sporothrix spanning more than 14 000 human and animal case reports. Persoonia 2015, 35, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Arenas, R.; Sánchez-Cardenas, C.D.; Ramirez-Hobak, L.; Ruíz Arriaga, L.F.; Vega Memije, M.E. Sporotrichosis: From KOH to molecular biology. J. Fungi 2018, 4, 62. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.M.; Almeida-Paes, R.; Gutierrez-Galhardo, M.C.; Zancope-Oliveira, R.M. Molecular identification of the Sporothrix schenckii complex. Rev. Iberoam. Micol. 2014, 31, 2–6. [Google Scholar] [CrossRef]
- Bernardes-Engemann, A.R.; Loureiro y Penha, C.V.; Benvenuto, F.; Braga, J.U.; Barros, M.L.; Orofino-Costa, R.; Lopes-Bezerra, L.M. A comparative serological study of the SsCBF antigenic fraction isolated from three Sporothrix schenckii strains. Med. Mycol. 2009, 47, 874–878. [Google Scholar] [CrossRef] [Green Version]
- Parreiras de Jesus, A.C.; Grossi de Oliveira, A.; Sernizon Guimarães, N.; Mendoza, L.; Toshio Fujiwara, R.; Alves da Silva Menezes, C.; Rocha Vilela, R.V. Serological tests using Sporothrix species antigens for the accurate diagnosis of sporotrichosis: A meta-analysis. Diagn. Microbiol. Infect. Dis. 2020, 98, 115131. [Google Scholar] [CrossRef]
- Orofino-Costa, R.; Macedo, P.M.; Rodrigues, A.M.; Bernardes-Engemann, A.R. Sporotrichosis: An update on epidemiology, etiopathogenesis, laboratory and clinical therapeutics. An. Bras. Dermatol. 2017, 92, 606–620. [Google Scholar] [CrossRef]
- Zhou, X.; Rodrigues, A.M.; Feng, P.; Hoog, G.S. Global ITS diversity in the Sporothrix schenckii complex. Fungal Divers. 2013, 66, 153–165. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; de Hoog, G.S.; de Camargo, Z.P. Genotyping species of the Sporothrix schenckii complex by PCR-RFLP of calmodulin. Diagn. Microbiol. Infect. Dis. 2014, 78, 383–387. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; de Hoog, G.S.; de Camargo, Z.P. Molecular diagnosis of pathogenic Sporothrix species. PLoS Negl. Trop. Dis. 2015, 9, e0004190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Li, F.; Li, R.; Gong, J.; Zhao, F. Fast diagnosis of sporotrichosis caused by Sporothrix globosa, Sporothrix schenckii, and Sporothrix brasiliensis based on multiplex real-time PCR. PLoS Negl. Trop. Dis. 2019, 13, 2, e0007219. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Kaminaga, T.; Baba, A.; Koike, S.; Koike, M.; Kanno, M.; Ishikawa, S.; Tsukada, K.; Suzuki, T.; Hamasaki, Y.; et al. Diagnostic value of a nested polymerase chain reaction for diagnosing cutaneous sporotrichosis from paraffin-embedded skin tissue. Mycoses 2019, 62, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, J.A.; Hagen, F.; Fisher, M.C.; de Camargo, Z.P.; Rodrigues, A.M. Genome-wide mapping using new AFLP markers to explore intraspecific variation among pathogenic Sporothrix species. PLoS Negl. Trop. Dis. 2020, 14, 7, e0008330. [Google Scholar] [CrossRef] [PubMed]
- Sanguinetti, M.; Posteraro, B. Identification of molds by matrix-assisted laser desorption ionization-time of flight mass spectrometry. J. Clin. Microbiol. 2017, 55, 369–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkendorf, L.S.; Bowles, E.; Buil, J.B.; van der Lee, H.A.L.; Posteraro, B.; Sanguinetti, M.; Verweij, P.E. Update on matrix-assisted laser desorption ionization-time of flight mass spectrometry identification of filamentous fungi. J. Clin. Microbiol. 2020, 58, e01263-20. [Google Scholar] [CrossRef]
- Oliveira, M.M.; Santos, C.; Sampaio, P.; Romeo, O.; Almeida-Paes, R.; Pais, C.; Lima, N.; Zancopé-Oliveira, R.M. Development and optimization of a new MALDI-TOF protocol for identification of the Sporothrix species complex. Res. Microbiol. 2015, 166, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Matos, A.M.F.; Moreira, L.M.; Barczewski, B.F.; de Matos, L.X.; de Oliveira, J.B.V.; Pimentel, M.I.F.; Almeida-Paes, R.; Oliveira, M.G.; Pinto, T.C.A.; Lima, N.; et al. Identification by MALDI-TOF MS of Sporothrix brasiliensis isolated from a subconjunctival infiltrative lesion in an immunocompetent patient. Microorganisms 2019, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Lau, A.F. Matrix-assisted laser desorption ionization time-of-flight for fungal identification. Clin. Lab. Med. 2021, 41, 267–283. [Google Scholar] [CrossRef] [PubMed]
- Prakash, H.; Karuppiah, P.A.; Al-Dhabi, N.; Prasad, G.S.; Badapanda, C.; Chakrabarti, A.; Rudramurthy, S.M. Comparative genomics of Sporothrix species and identification of putative pathogenic-gene determinants. Future Microbiol. 2020, 1465–1481. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Ma, Z.; Zhou, X. Comparative genomic data provide new insight on the evolution of pathogenicity in Sporothrix species. Front. Microbiol. 2020, 11, 565439. [Google Scholar] [CrossRef] [PubMed]
- Sil, A.; Andrianopoulos, A. Thermally dimorphic human fungal pathogens--polyphyletic pathogens with a convergent pathogenicity trait. Cold Spring Harb. Perspect. Med. 2014, 5, a019794. [Google Scholar] [CrossRef] [Green Version]
- Da Cruz Bahiense Rocha, I.; Terra, P.P.D.; Cardoso de Oliveira, R.; Lubianca Zanotti, R.; Falqueto, A.; de Camargo, Z.P.; Rodrigues, A.M.; Gonçalves, S.S. Molecular-based assessment of diversity and population structure of Sporothrix spp. clinical isolates from Espírito Santo-Brazil. Mycoses 2021, 64, 420–427. [Google Scholar] [CrossRef]
- Giosa, D.; Felice, M.R.; Giuffrè, L.; Aiese Cigliano, R.; Paytuví-Gallart, A.; Lo Passo, C.; Barresi, C.; D’Alessandro, E.; Huang, H.; Criseo, G.; et al. Transcriptome-wide expression profiling of Sporothrix schenckii yeast and mycelial forms and the establishment of the Sporothrix Genome DataBase. Microb. Genom. 2020, 6, mgen000445. [Google Scholar] [CrossRef]
- Zheng, F.; Gao, W.; Wang, Y.; Chen, Q.; Zhang, Q.; Jiang, X.; Zhang, Z. Map of dimorphic switching-related signaling pathways in Sporothrix schenckii based on its transcriptome. Mol. Med. Rep. 2021, 24, 646. [Google Scholar] [CrossRef]
- Lopes-Bezerra, L.M.; Walker, L.A.; Niño-Vega, G.; Mora-Montes, H.M.; Neves, G.W.P.; Villalobos-Duno, H.; Barreto, L.; Garcia, K.; Franco, B.; Martínez-Álvarez, J.A.; et al. Cell walls of the dimorphic fungal pathogens Sporothrix schenckii and Sporothrix brasiliensis exhibit bilaminate structures and sloughing of extensive and intact layers. PLoS Negl. Trop. Dis. 2018, 12, e0006169. [Google Scholar] [CrossRef] [Green Version]
- Lopes-Bezerra, L.M. Sporothrix schenckii cell wall peptidorhamnomannans. Front. Microbiol. 2011, 2, 243. [Google Scholar] [CrossRef] [Green Version]
- Mora-Montes, H.M.; da Silva Dantas, A.; Trujillo-Esquivel, E.; de Souza Baptista, A.R.; Lopes-Bezerra, L.M. Current progress in the biology of members of the Sporothrix schenckii complex following the genomic era. FEMS Yeast Res. 2015, 15, 6. [Google Scholar] [CrossRef] [Green Version]
- Castro, R.A.; Kubitschek-Barreira, P.H.; Teixeira, P.A.; Sanchez, G.F.; Teixeira, M.M.; Quintella, L.P.; Almeida, S.R.; Costa, R.O.; Camargo, Z.P.; Felipe, M.S.S.; et al. Differences in cell morphometry, cell wall topography and gp70 expression correlate with the virulence of Sporothrix brasiliensis clinical isolates. PLoS ONE 2013, 8, e75656. [Google Scholar] [CrossRef] [Green Version]
- García-Carnero, L.C.; Salinas-Marín, R.; Lozoya-Pérez, N.E.; Wrobel, K.; Wrobel, K.; Martínez-Duncker, I.; Niño-Vega, G.A.; Mora-Montes, H.M. The heat shock protein 60 and Pap1 participate in the Sporothrix schenckii-host interaction. J. Fungi 2021, 7, 960. [Google Scholar] [CrossRef]
- Almeida-Paes, R.; Frases, S.; Araújo, G.S.; de Oliveira, M.M.; Gerfen, G.J.; Nosanchuk, J.D.; Zancopé-Oliveira, R.M. Biosynthesis and functions of a melanoid pigment produced by species of the Sporothrix complex in the presence of L-tyrosine. Appl. Environ. Microbiol. 2012, 78, 8623–8630. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, P.A.; De Castro, R.A.; Ferreira, F.R.; Cunha, M.M.; Torres, A.P.; Penha, C.V.; Rozental, S.; Lopes-Bezerra, L.M. L-DOPA accessibility in culture medium increases melanin expression and virulence of Sporothrix schenckii yeast cells. Med. Mycol. 2010, 48, 687–695. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, M.A.K.; de Almeida, J.R.F.; Jannuzzi, G.P.; Cronemberger-Andrade, A.; Torrecilhas, A.C.T.; Moretti, N.S.; da Cunha, J.P.C.; de Almeida, S.R.; Ferreira, K.S. Extracellular vesicles from Sporothrix brasiliensis are an important virulence factor that induce an increase in fungal burden in experimental sporotrichosis. Front. Microbiol. 2018, 9, 2286. [Google Scholar] [CrossRef]
- Silva-Bailão, M.G.; Lima, P.S.; Oliveira, M.; Oliveira, L.C.; Almeida-Paes, R.; Borges, C.L.; Bailão, A.M.; Coelho, A.; Soares, C.; Zancopé-Oliveira, R.M. Comparative proteomics in the three major human pathogenic species of the genus Sporothrix. Microbes Infect. 2021, 23, 104762. [Google Scholar] [CrossRef]
- Ortega, I.; Soares Felipe, M.S.; Vasconcelos, A.T.; Lopes Bezerra, L.M.; Da Silva Dantas, A. Peroxide sensing and signaling in the Sporothrix schenckii complex: An in silico analysis to uncover putative mechanisms regulating the Hog1 and AP-1 like signaling pathways. Med. Mycol. 2015, 53, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Herrera, R.; Flores-Villavicencio, L.L.; Pichardo-Molina, J.L.; Castruita-Domínguez, J.P.; Aparicio-Fernández, X.; Sabanero López, M.; Villagómez-Castro, J.C. Analysis of biofilm formation by Sporothrix schenckii. Med. Mycol. 2021, 59, 31–40. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Chandra, J. Candida biofilm resistance. Drug Resist. Updates 2004, 7, 301–309. [Google Scholar] [CrossRef]
- Bao, F.; Pan, Q.; Wang, Z.; Liu, H.; Zhang, F. Susceptibility testing of clinical isolates of Sporothrix globosa in Shandong, China. Mycoses 2020, 63, 1191–1194. [Google Scholar] [CrossRef]
- Waller, S.B.; Dalla Lana, D.F.; Quatrin, P.M.; Ferreira, M.R.A.; Fuentefria, A.M.; Mezzari, A. Antifungal resistance on Sporothrix species: An overview. Braz. J. Microbiol. 2021, 52, 73–80. [Google Scholar] [CrossRef]
- Flórez-Muñoz, S.V.; Alzate, J.F.; Mesa-Arango, A.C. Molecular identification and antifungal susceptibility of clinical isolates of Sporothrix schenckii complex in Medellin, Colombia. Mycopathologia 2019, 184, 53–63. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; de Hoog, G.S.; de Cássia Pires, D.; Brihante, R.S.; Sidrim, J.J.; Gadelha, M.F.; Colombo, A.L.; de Camargo, Z.P. Genetic diversity and antifungal susceptibility profiles in causative agents of sporotrichosis. BMC Infect. Dis. 2014, 14, 219. [Google Scholar] [CrossRef] [Green Version]
- Brilhante, R.S.N.; de Aguiar, F.R.M.; da Silva, M.L.Q.; de Oliveira, J.S.; de Camargo, Z.P.; Rodrigues, A.M.; Pereira, V.S.; Serpa, R.; Castelo-Branco, D.S.C.M.; Correia, E.E.M.; et al. Antifungal susceptibility of Sporothrix schenckii complex biofilms. Med. Mycol. 2018, 56, 297–306. [Google Scholar] [CrossRef]
- Matowane, R.G.; Wieteska, L.; Bamal, H.D.; Kgosiemang, I.K.R.; van Wyk, M.; Manume, N.A.; Abdalla, S.M.H.; Mashele, S.S.; Gront, D.; Syed, K. In silico analysis of cytochrome P450 monooxygenases in chronic granulomatous infectious fungus Sporothrix schenckii: Special focus on CYP51. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 166–177. [Google Scholar] [CrossRef]
- Espinel-Ingroff, A.; Abreu, D.P.B.; Almeida-Paes, R.; Brilhante, R.S.N.; Chakrabarti, A.; Chowdhary, A.; Hagen, F.; Córdoba, S.; Gonzalez, G.M.; Govender, N.P.; et al. Multicenter, international study of MIC/MEC distributions for definition of epidemiological cutoff values for Sporothrix species identified by molecular methods. Antimicrob. Agents Chemother. 2017, 61, e01057-17. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Fourth Informational Supplement. CLSI document M27-S4; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi; Approved Standard—Second Edition. CLSI document M38-A2; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]
- Kohler, L.M.; Soares, B.M.; de Assis, S.D.; Da Silva Barros, M.E.; Hamdan, J.S. In vitro susceptibility of isolates of Sporothrix schenckii to amphotericin B, itraconazole, and terbinafine: Comparison of yeast and mycelial forms. Can. J. Microbiol. 2006, 52, 843–847. [Google Scholar] [CrossRef]
- Lopes-Bezerra, L.M.; Mora-Montes, H.M.; Zhang, Y.; Nino-Vega, G.; Rodrigues, A.M.; de Camargo, Z.P.; de Hoog, S. Sporotrichosis between 1898 and 2017: The evolution of knowledge on a changeable disease and on emerging etiological agents. Med. Mycol. 2018, 56, S126–S143. [Google Scholar] [CrossRef]
- Friedman, D.Z.P.; Schwartz, I.S. Emerging fungal infections: New patients, new patterns, and new pathogens. J. Fungi 2019, 5, 67. [Google Scholar] [CrossRef] [Green Version]
- Rossow, J.A.; Queiroz-Telles, F.; Caceres, D.H.; Beer, K.D.; Jackson, B.R.; Pereira, J.G.; Ferreira Gremião, I.D.; Pereira, S.A. A one health approach to combatting Sporothrix brasiliensis: Narrative review of an emerging zoonotic fungal pathogen in South America. J. Fungi 2020, 6, 247. [Google Scholar] [CrossRef]
- Waller, S.B.; Cleff, M.B.; Dalla Lana, D.F.; de Mattos, C.B.; Guterres, K.A.; Freitag, R.A.; Sallis, E.S.V.; Fuentefria, A.M.; de Mello, J.R.B.; de Faria, R.O.; et al. Can the essential oil of rosemary (Rosmarinus officinalis Linn.) protect rats infected with itraconazole-resistant Sporothrix brasiliensis from fungal spread? J. Mycol. Med. 2021, 31, 101199. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Carolis, E.; Posteraro, B.; Sanguinetti, M. Old and New Insights into Sporothrix schenckii Complex Biology and Identification. Pathogens 2022, 11, 297. https://doi.org/10.3390/pathogens11030297
De Carolis E, Posteraro B, Sanguinetti M. Old and New Insights into Sporothrix schenckii Complex Biology and Identification. Pathogens. 2022; 11(3):297. https://doi.org/10.3390/pathogens11030297
Chicago/Turabian StyleDe Carolis, Elena, Brunella Posteraro, and Maurizio Sanguinetti. 2022. "Old and New Insights into Sporothrix schenckii Complex Biology and Identification" Pathogens 11, no. 3: 297. https://doi.org/10.3390/pathogens11030297
APA StyleDe Carolis, E., Posteraro, B., & Sanguinetti, M. (2022). Old and New Insights into Sporothrix schenckii Complex Biology and Identification. Pathogens, 11(3), 297. https://doi.org/10.3390/pathogens11030297