
Wild Bird Densities and Landscape Variables Predict Spatial Patterns in HPAI Outbreak Risk across The Netherlands

 , , , , and
, , , , and
Abstract
:1. Introduction
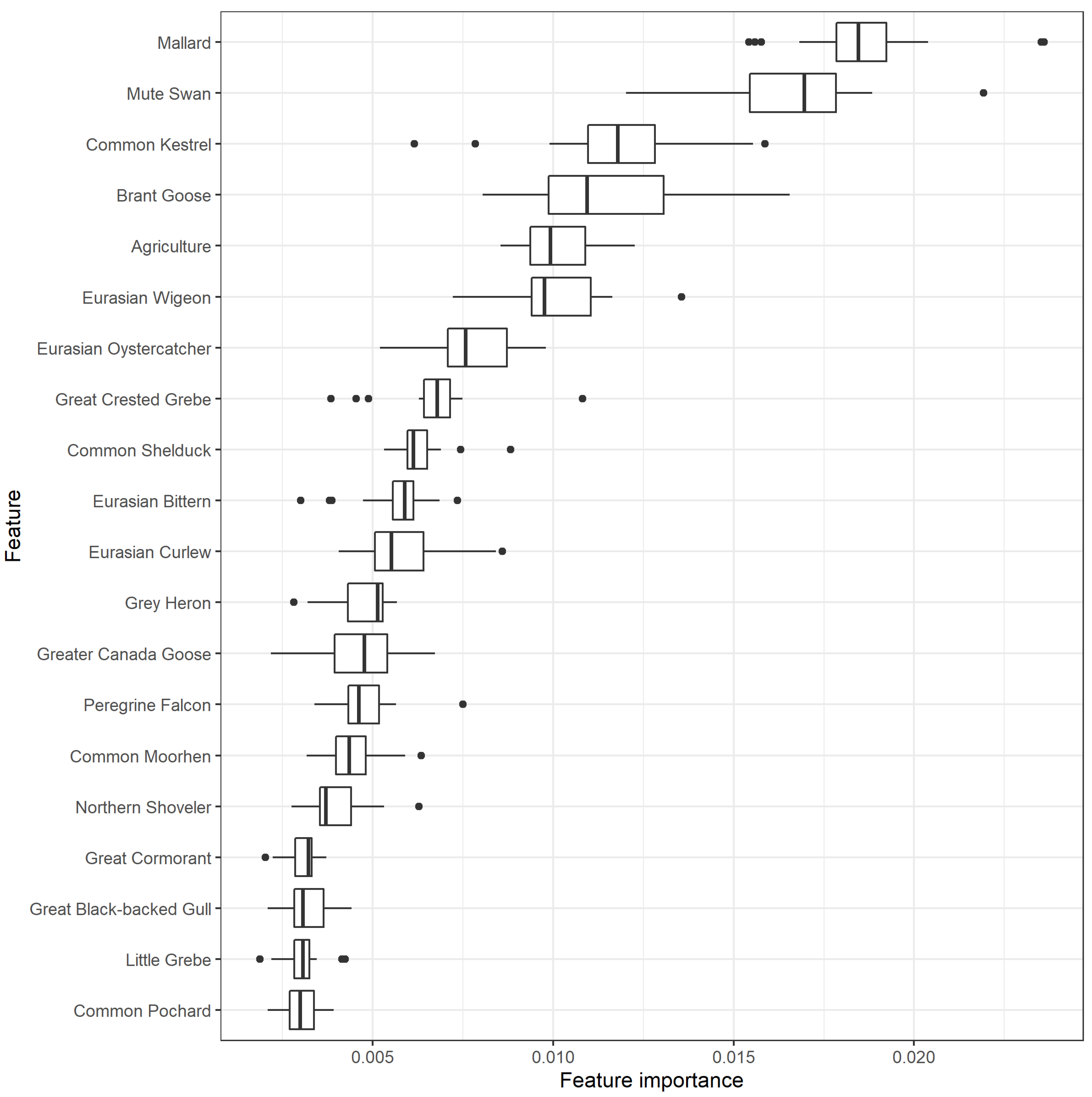
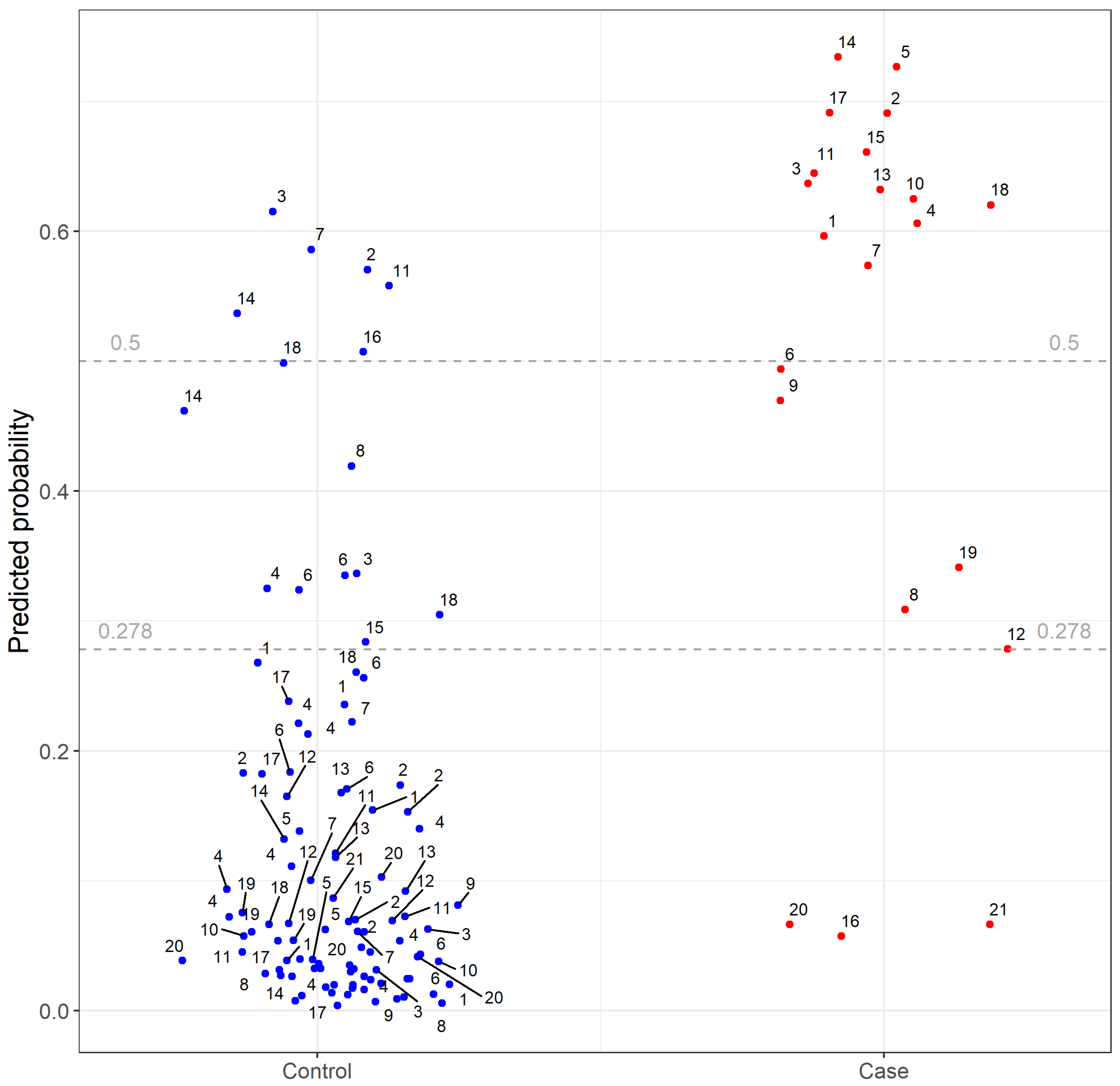
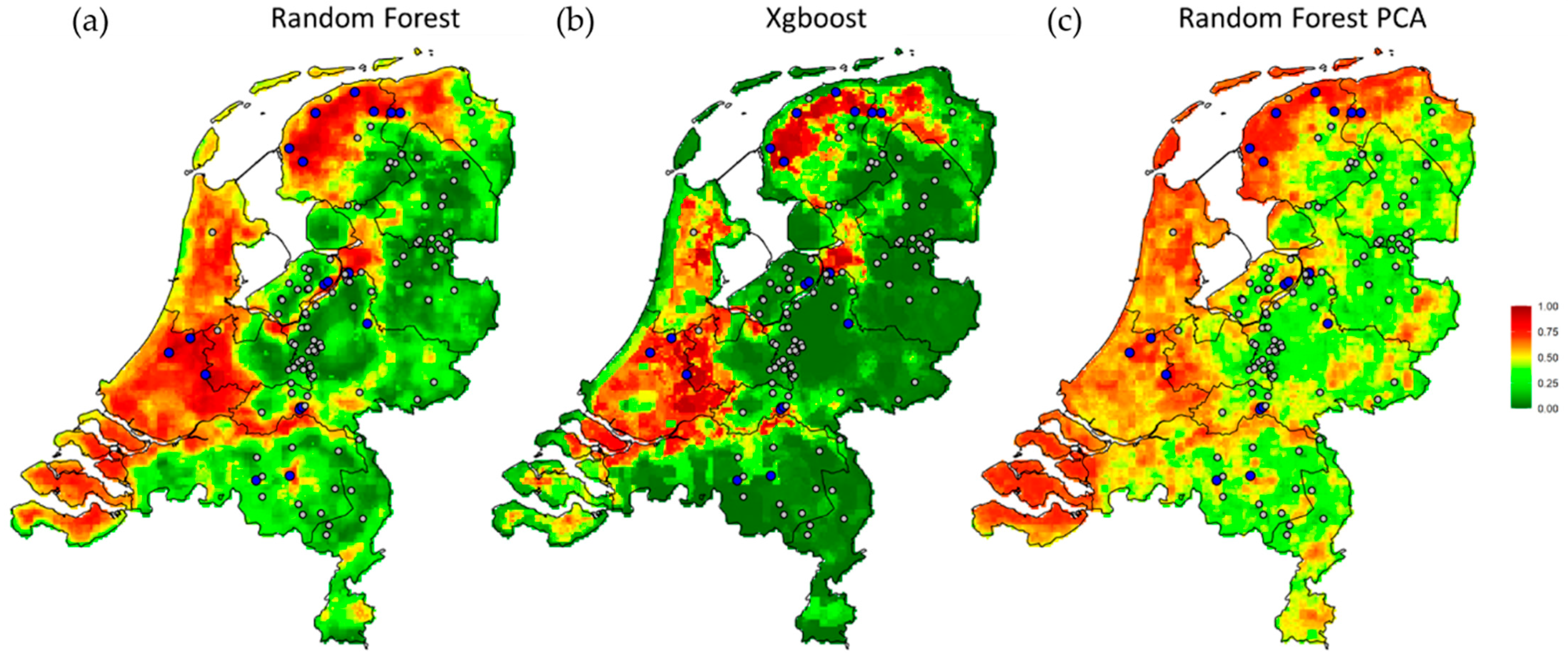
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Design
4.1.1. Case Farms
4.1.2. Control Farms
4.2. Selection of Wild Bird Density Data
4.3. Bird Count Data
4.4. Land Cover Data
4.5. Including Spatial Context
4.6. Cross-Scale Analyses and Dimension Reduction
4.7. Model Training and Evaluation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Bird Count Data Collection and Analyses
References
- Verhagen, J.H.; van der Jeugd, H.P.; Nolet, B.A.; Slaterus, R.; Kharitonov, S.P.; de Vries, P.P.; Vuong, O.; Majoor, F.; Kuiken, T.; Fouchier, R.A. Wild bird surveillance around outbreaks of highly pathogenic avian influenza A(H5N8) virus in the Netherlands, 2014, within the context of global flyways. Euro Surveill. 2015, 20, 21–32. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Avian influenza overview August–December 2020. EFSA J. 2020, 18, e06379. [Google Scholar] [CrossRef]
- Lycett, S.J.; Bodewes, R.; Pohlmann, A.; Banks, J.; Bányai, K.; Boni, M.F.; Bouwstra, R.; Breed, A.C.; Brown, I.H.; Chen, H.; et al. Role for migratory wild birds in the global spread of avian influenza H5N8. Science 2016, 354, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Beerens, N.; Heutink, R.; Bergervoet, S.A.; Harders, F.; Bossers, A.; Koch, G. Multiple reassorted viruses as cause of highly pathogenic avian influenza A(H5N8) virus epidemic, the Netherlands, 2016. Emerg. Infect. Dis. 2017, 23, 1966–1973. [Google Scholar] [CrossRef]
- Beerens, N.; Koch, G.; Heutink, R.; Harders, F.; Vries, D.P.E.E.; Ho, C.; Bossers, A.; Elbers, A.; Nancy, B.; Guus, K.; et al. Novel Highly Pathogenic Avian Influenza A(H5N6) Virus in the Netherlands, December 2017. Emerg. Infect. Dis. 2018, 24, 770. [Google Scholar] [CrossRef]
- Kleyheeg, E.; Slaterus, R.; Bodewes, R.; Rijks, J.M.; Spierenburg, M.A.H.; Beerens, N.; Kelder, L.; Poen, M.J.; Stegeman, J.A.; Fouchier, R.A.M.; et al. Deaths among wild birds during highly pathogenic avian influenza A(H5N8) virus outbreak, the Netherlands. Emerg. Infect. Dis. 2017, 23, 2050–2054. [Google Scholar] [CrossRef]
- Verhagen, J.H.; Herfst, S.; Fouchier, R.A. Infectious disease. How a virus travels the world. Science 2015, 347, 616–617. [Google Scholar] [CrossRef]
- Bouwstra, R.; Heutink, R.; Bossers, A.; Harders, F.; Koch, G.; Elbers, A. Full-genome sequence of influenza A(H5N8) virus in poultry linked to sequences of strains from Asia, the Netherlands, 2014. Emerg. Infect. Dis. 2015, 21, 872–874. [Google Scholar] [CrossRef]
- Napp, S.; Majó, N.; Sánchez-Gónzalez, R.; Vergara-Alert, J. Emergence and spread of highly pathogenic avian influenza A(H5N8) in Europe in 2016-2017. Transbound. Emerg. Dis. 2018, 65, 1217–1226. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Avian influenza overview September–December 2021. EFSA J. 2021, 19, e07108. [Google Scholar] [CrossRef]
- EFSA. Avian influenza overview February–May 2021. EFSA J. 2021, 19, e06951. [Google Scholar] [CrossRef]
- FAO. Global AIV with Zoonotic Potential Situation Update 23 February 2022. Available online: https://www.fao.org/ag/againfo/programmes/en/empres/Global_AIV_Zoonotic_Update/situation_update.html (accessed on 18 March 2022).
- ADIS. European Commission’s Animal Disease Information System-HPAI Positive Events by Country and Poultry Category (2020–2021). Available online: https://www.izsvenezie.com/documents/reference-laboratories/avian-influenza/europe-updates/HPAI/2020-1/total-events.pdf (accessed on 18 March 2022).
- ADIS. European Commission’s Animal Disease Information System-HPAI Positive Events by Country and Poultry Category (2021–2022). Available online: https://www.izsvenezie.com/documents/reference-laboratories/avian-influenza/europe-updates/HPAI/2021-1/total-events.pdf (accessed on 18 March 2022).
- ADIS. European Commission’s Animal Disease Information System-Wild Birds: Species Affected 2020–2021. Available online: https://izsvenezie.com/documents/reference-laboratories/avian-influenza/europe-updates/HPAI/2020-1/wild-birds.pdf (accessed on 18 March 2022).
- ADIS. European Commission’s Animal Disease Information System-Wild Birds: Species Affected 2021–2022. Available online: https://izsvenezie.com/documents/reference-laboratories/avian-influenza/europe-updates/HPAI/2021-1/wild-birds.pdf (accessed on 18 March 2022).
- Belkhiria, J.; Hijmans, R.J.; Boyce, W.; Crossley, B.M.; Martínez-López, B. Identification of high risk areas for avian influenza outbreaks in California using disease distribution models. PLoS ONE 2018, 13, e0190824. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.; Pfeiffer, D.U. Risk factor modelling of the spatio-temporal patterns of highly pathogenic avian influenza (HPAIV) H5N1: A review. Spat. Spatiotemporal Epidemiol. 2012, 3, 173–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, Y.; de Boer, W.F.; Gong, P. Different Environmental Drivers of Highly Pathogenic Avian Influenza H5N1 Outbreaks in Poultry and Wild Birds. PLoS ONE 2013, 8, e53362. [Google Scholar] [CrossRef]
- Velkers, F.C.; Manders, T.T.M.; Vernooij, J.C.M.; Stahl, J.; Slaterus, R.; Stegeman, J.A. Association of wild bird densities around poultry farms with the risk of highly pathogenic avian influenza virus subtype H5N8 outbreaks in the Netherlands, 2016. Transbound. Emerg. Dis. 2020, 68, 76–87. [Google Scholar] [CrossRef]
- Beerens, N.; Heutink, R.; Pritz-Verschuren, S.; Germeraad, E.A.; Bergervoet, S.A.; Harders, F.; Bossers, A.; Koch, G. Genetic relationship between poultry and wild bird viruses during the highly pathogenic avian influenza H5N6 epidemic in the Netherlands, 2017–2018. Transbound. Emerg. Dis. 2019, 66, 1370–1378. [Google Scholar] [CrossRef]
- Elbers, A.R.W.; Gonzales, J.L. Quantification of visits of wild fauna to a commercial free-range layer farm in the Netherlands located in an avian influenza hot-spot area assessed by video-camera monitoring. Transbound. Emerg. Dis. 2020, 67, 661–677. [Google Scholar] [CrossRef] [Green Version]
- Velkers, F.C.; Blokhuis, S.J.; Veldhuis Kroeze, E.J.B.; Burt, S.A. The role of rodents in avian influenza outbreaks in poultry farms: A review. Vet. Q. 2017, 37, 182–194. [Google Scholar] [CrossRef]
- Root, J.; Shriner, S.A. Avian influenza A virus associations in wild, terrestrial mammals: A review of potential synanthropic vectors to poultry facilities. Viruses 2021, 12, 1352. [Google Scholar] [CrossRef]
- Yoo, D.S.; Lee, K.N.; Chun, B.C.; Lee, H.S.; Park, H.; Kim, J.K. Preventive effect of on-farm biosecurity practices against highly pathogenic avian influenza (HPAI) H5N6 infection on commercial layer farms in the Republic of Korea during the 2016-17 epidemic: A case-control study. Prev. Vet. Med. 2022, 199, 105556. [Google Scholar] [CrossRef]
- Beerens, N.; Heutink, R.; Harders, F.; Roose, M.; Pritz-Verschuren, S.B.E.; Germeraad, E.A.; Engelsma, M. Incursion of novel highly pathogenic avian influenza A(H5N8) virus, the Netherlands, October 2020. Emerg. Infect. Dis. 2021, 27, 1750–1753. [Google Scholar] [CrossRef] [PubMed]
- Caron, A.; Cappelle, J.; Gaidet, N. Challenging the conceptual framework of maintenance hosts for influenza A viruses in wild birds. J. Appl. Ecol. 2017, 54, 681–690. [Google Scholar] [CrossRef]
- Lisovski, S.; van Dijk, J.G.B.; Klinkenberg, D.; Nolet, B.A.; Fouchier, R.A.M.; Klaassen, M. The roles of migratory and resident birds in local avian influenza infection dynamics. J. Appl. Ecol. 2018, 55, 2963–2975. [Google Scholar] [CrossRef] [PubMed]
- Yoo, D.S.; Kang, S.I.; Lee, Y.N.; Lee, E.K.; Kim, W.Y.; Lee, Y.J. Bridging the Local Persistence and Long-Range Dispersal of Highly Pathogenic Avian Influenza Virus (HPAIv): A Case Study of HPAIv-Infected Sedentary and Migratory Wildfowls Inhabiting Infected Premises. Viruses 2022, 14, 116. [Google Scholar] [CrossRef]
- Alarcon, P.; Brouwer, A.; Venkatesh, D.; Duncan, D.; Dovas, C.I.; Georgiades, G.; Monne, I.; Fusaro, A.; Dan, A.; Śmietanka, K.; et al. Comparison of 2016-17 and Previous Epizootics of Highly Pathogenic Avian Influenza H5 Guangdong Lineage in Europe. Emerg. Infect. Dis. 2018, 24, 2270–2283. [Google Scholar] [CrossRef]
- Hill, N.J.; Takekawa, J.Y.; Ackerman, J.T.; Hobson, K.A.; Herring, G.; Cardona, C.J.; Runstadler, J.A.; Boyce, W.M. Migration strategy affects avian influenza dynamics in mallards (Anas platyrhynchos). Mol. Ecol. 2012, 21, 5986–5999. [Google Scholar] [CrossRef]
- Yoo, D.S.; Lee, K.; Beatriz, M.L.; Chun, B.C.; Belkhiria, J.; Lee, K.N. Spatiotemporal risk assessment for avian influenza outbreak based on the dynamics of habitat suitability for wild birds. Transbound. Emerg. Dis. 2021, 1–15. [Google Scholar] [CrossRef]
- Scolamacchia, F.; Mulatti, P.; Mazzucato, M.; Barbujani, M.; Harvey, W.T.; Fusaro, A.; Monne, I.; Marangon, S. Different environmental gradients associated to the spatiotemporal and genetic pattern of the H5N8 highly pathogenic avian influenza outbreaks in poultry in Italy. Transbound. Emerg. Dis. 2021, 68, 152–167. [Google Scholar] [CrossRef]
- Huang, Z.Y.X.; Xu, C.; van Langevelde, F.; Ma, Y.; Langendoen, T.; Mundkur, T.; Si, Y.; Tian, H.; Kraus, R.H.S.; Gilbert, M.; et al. Contrasting effects of host species and phylogenetic diversity on the occurrence of HPAI H5N1 in European wild birds. J. Anim. Ecol. 2019, 88, 1044–1053. [Google Scholar] [CrossRef]
- McDuie, F.; Matchett, E.L.; Prosser, D.J.; Takekawa, J.Y.; Pitesky, M.E.; Lorenz, A.A.; McCuen, M.M.; Overton Cory, T.; Ackerman, J.T.; De La Cruz, S.E.W.; et al. Pathways for avian influenza virus spread: GPS reveals wild waterfowl in commercial livestock facilities and connectivity with the natural wetland landscape. Transbound. Emerg. Dis. 2022, 1–15. [Google Scholar] [CrossRef]
- Humphreys, J.M.; Douglas, D.C.; Ramey, A.M.; Mullinax, J.M.; Soos, C.; Link, P.; Walther, P.; Prosser, D.J. The spatial–temporal relationship of blue-winged teal to domestic poultry: Movement state modelling of a highly mobile avian influenza host. J. Appl. Ecol. 2021, 58, 2040–2052. [Google Scholar] [CrossRef]
- Gonzales, J.L.; Pritz-Verschuren, S.; Bouwstra, R.; Wiegel, J.; Elbers, A.R.W.; Beerens, N. Seasonal risk of low pathogenic avian influenza virus introductions into free-range layer farms in the Netherlands. Transbound. Emerg. Dis. 2020, 68, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.C.; Lee, Y.J.; Song, B.M.; Kang, H.M.; Lee, E.K.; Hanna, A.; Gilbert, M.; Brown, I.H.; Pybus, O.G. Wild waterfowl migration and domestic duck density shape the epidemiology of highly pathogenic H5N8 influenza in the Republic of Korea. Infect. Genet. Evol. 2015, 34, 267–277. [Google Scholar] [CrossRef] [PubMed]
- de Knegt, H.J.; van Langevelde, F.; Skidmore, A.K.; Delsink, A.; Slotow, R.; Henley, S.; Bucini, G.; de Boer, W.F.; Coughenour, M.B.; Grant, C.C.; et al. The spatial scaling of habitat selection by African elephants. J. Anim. Ecol. 2011, 80, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Verhagen, J.H.; Fouchier, R.A.M.; Lewis, N. Highly pathogenic avian influenza viruses at the wild–domestic bird interface in Europe: Future directions for research and surveillance. Viruses 2021, 13, 212. [Google Scholar] [CrossRef]
- Animal and Plant Health Agency (UK); Erasmus Medical Centre (NL); Friedrich Loeffler Institute (DE); Istituto Zooprofilattico Sperimentale delle Venezie (IT); Linnaeus University (SE); Wageningen University (NL). Report about HPAI introduction into Europe, HPAI detection in wild birds and HPAI spread between European holdings in the period 2005–2015. EFSA Supporting Publ. 2017, 14, 1284E. [Google Scholar] [CrossRef]
- Bouwstra, R.J.; Koch, G.; Heutink, R.; Harders, F.; Van Der Spek, A.; Elbers, A.R.; Bossers, A. Phylogenetic analysis of highly pathogenic avian influenza A(H5N8) virus outbreak strains provides evidence for four separate introductions and one between-poultry farm transmission in the Netherlands, November 2014. Euro Surveill. 2015, 20, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Poen, M.J.; Bestebroer, T.M.; Vuong, O.; Scheuer, R.D.; van der Jeugd, H.P.; Kleyheeg, E.; Eggink, D.; Lexmond, P.; van den Brand, J.M.A.; Begeman, L.; et al. Local amplification of highly pathogenic avian influenza H5N8 viruses in wild birds in the Netherlands, 2016 to 2017. Euro Surveill. 2018, 23, 17-00449. [Google Scholar] [CrossRef]
- Sovon Dutch Center for Field Ornithology. Bird Atlas of the Netherlands, 1st ed.; Kosmos Publishers: Utrecht, The Netherlands; Antwerp, Belgium, 2018; pp. 1–640. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 10 August 2021).
- de Knegt, H.J.; van Langevelde, F.; Coughenour, M.B.; Skidmore, A.K.; de Boer, W.F.; Heitkönig, I.M.A.; Knox, N.M.; Slotow, R.; van der Waal, C.; Prins, H.H.T. Spatial autocorrelation and the scaling of species–environment relationships. Ecology 2010, 91, 2455–2465. [Google Scholar] [CrossRef] [Green Version]
- Holland, J.D.; Bert, D.G.; Fahrig, L. Determining the Spatial Scale of Species’ Response to Habitat. Bioscience 2004, 54, 227–233. [Google Scholar] [CrossRef]
- Therneau, T. A Package for Survival Analysis in R, 2020. R Package Version 3.2-7. 2020. Available online: https://cran.r-project.org/package=survival (accessed on 10 August 2021).
- Wright, M.N.; Ziegler, A. ranger: A Fast Implementation of Random Forests for High Dimensional Data in C++ and R. J. Stat. Softw. 2017, 77, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Altmann, A.; Toloşi, L.; Sander, O.; Lengauer, T. Permutation importance: A corrected feature importance measure. Bioinformatics 2010, 26, 1340–1347. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Algorithm | Accuracy | Recall/Sensitivity | AUC-ROC |
|---|---|---|---|
| RF | 0.86 | 0.88 | 0.92 |
| GBT | 0.94 | 0.88 | 0.94 |
| RF-PCA | 0.87 | 0.81 | 0.88 |
| Case-ID | Poultry Type | 2014-H5N8 | 2016-H5N8 | 2017-H5N6 | 2018-H5N6 | 2020-H5N8 1 |
|---|---|---|---|---|---|---|
| 1 | Layer | x | x | x | ||
| 2 | Layer | x | ||||
| 3 | Layer | x | ||||
| 4 | Pekin Duck | x | x | x | ||
| 5 | Broiler Breeder | x | ||||
| 6 | Pekin Duck | x | x | |||
| 7 | Pekin Duck | x | ||||
| 8 | Pekin Duck | x | ||||
| 9 | Layer | x | ||||
| 10 | Layer | x | ||||
| 11 | Broiler Breeder | x | ||||
| 12 | Broiler Breeder | x | ||||
| 13 | Broiler Breeder | x 1 | ||||
| 14 | Layer | x | ||||
| 15 | Layer | x | ||||
| 16 | Pekin Duck | x | ||||
| 17 | Broiler | x | ||||
| 18 | Broiler | x | ||||
| 19 | Broiler Breeder | x | ||||
| 20 | Turkey | x | ||||
| 21 | Layer | x |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schreuder, J.; de Knegt, H.J.; Velkers, F.C.; Elbers, A.R.W.; Stahl, J.; Slaterus, R.; Stegeman, J.A.; de Boer, W.F. Wild Bird Densities and Landscape Variables Predict Spatial Patterns in HPAI Outbreak Risk across The Netherlands. Pathogens 2022, 11, 549. https://doi.org/10.3390/pathogens11050549
Schreuder J, de Knegt HJ, Velkers FC, Elbers ARW, Stahl J, Slaterus R, Stegeman JA, de Boer WF. Wild Bird Densities and Landscape Variables Predict Spatial Patterns in HPAI Outbreak Risk across The Netherlands. Pathogens. 2022; 11(5):549. https://doi.org/10.3390/pathogens11050549
Chicago/Turabian StyleSchreuder, Janneke, Henrik J. de Knegt, Francisca C. Velkers, Armin R. W. Elbers, Julia Stahl, Roy Slaterus, J. Arjan Stegeman, and Willem F. de Boer. 2022. "Wild Bird Densities and Landscape Variables Predict Spatial Patterns in HPAI Outbreak Risk across The Netherlands" Pathogens 11, no. 5: 549. https://doi.org/10.3390/pathogens11050549
APA StyleSchreuder, J., de Knegt, H. J., Velkers, F. C., Elbers, A. R. W., Stahl, J., Slaterus, R., Stegeman, J. A., & de Boer, W. F. (2022). Wild Bird Densities and Landscape Variables Predict Spatial Patterns in HPAI Outbreak Risk across The Netherlands. Pathogens, 11(5), 549. https://doi.org/10.3390/pathogens11050549