
Role of a Putative Alkylhydroperoxidase Rv2159c in the Oxidative Stress Response and Virulence of Mycobacterium tuberculosis

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Construction of Mtb∆2159 Mutant in M. tuberculosis
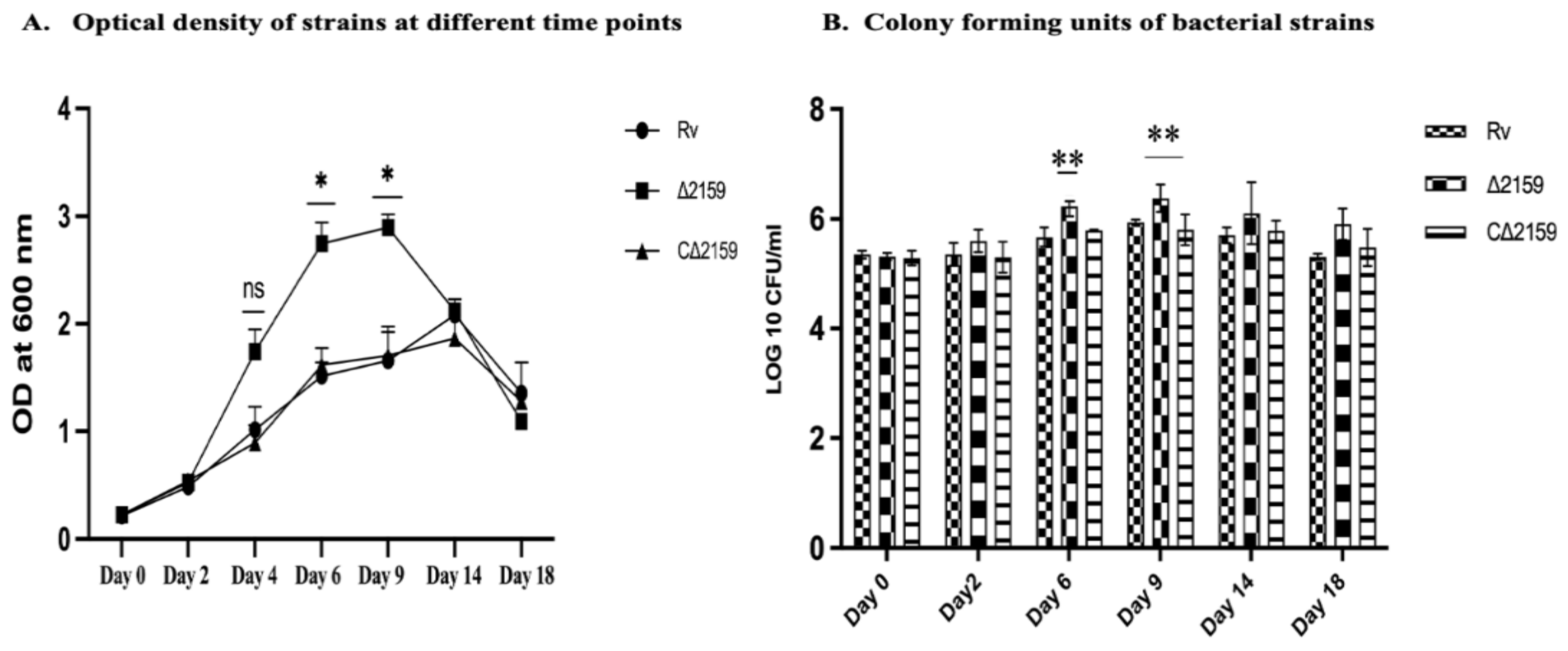
2.2. Growth Kinetics of Mtb∆2159
2.3. Viability of Mtb∆2159 during H2O2 Stress
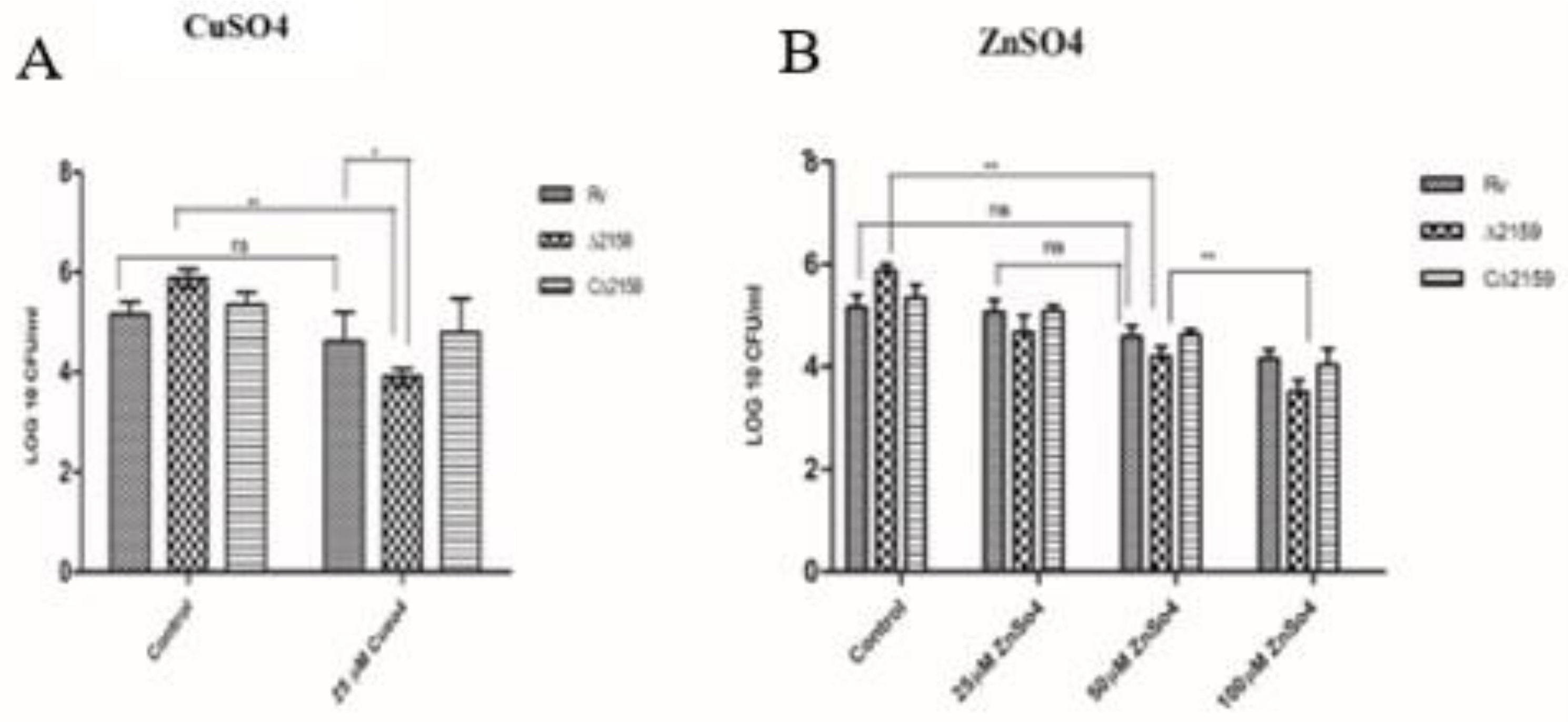
2.4. Effect of CuSO4 and ZnSO4 Treatment in Mtb∆2159
2.5. Survival of Mtb∆2159 in THP-1 Cells
2.6. Cytokine Expression
2.7. Mtb∆2159 Is Attenuated for In Vivo Growth in a Guinea Pig Model of Infection
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plasmids, Bacterial Strains, and Growth Conditions
5.2. Construction of Four Fragment Ligation
5.3. Construction of Transducing Phages Using AES
5.4. Specialized Transduction
5.5. Complementation of Mtb∆2159
5.6. Growth Kinetics Analysis
5.7. Colony Morphology Analysis
5.8. Growth Analysis during Exposure to H2O2, Transition Metals, and SDS
5.9. Exposure to Transition Metals and SDS Stress
5.10. THP-1 Cell Line Infection
5.11. Cytokine ELISA
5.12. Infection Studies in Guinea Pigs
5.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Tuberculosis Report 2021; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Uribe-Quero, E.; Rosales, C. Control of Phagocytosis by Microbial Pathogens. Front. Immunol. 2017, 8, 1368. [Google Scholar]
- Jamwal, S.V.; Mehrotra, P.; Singh, A.; Siddiqui, Z.; Basu, A.; Rao, K.V.S. Mycobacterial Escape from Macrophage Phagosomes to the Cytoplasm Represents an Alternate Adaptation Mechanism. Sci. Rep. 2016, 6, 23089. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxidative Med. Cell. Longev. 2016, 1245049. [Google Scholar] [CrossRef]
- Deretic, V.; Philipp, W.; Dhandayuthapani, S.; Mudd, M.H.; Curcic, R.; Garbe, T.; Heym, B.; Via, L.E.; Cole, S.T. Mycobacterium tuberculosis Is a Natural Mutant with an Inactivated Oxidative-stress Regulatory Gene: Implications for Sensitivity to Isoniazid. Mol. Microbiol. 1995, 17, 889–900. [Google Scholar] [CrossRef] [PubMed]
- Deretic, V.; Song, J.; Pagán-Ramos, E. Loss of OxyR in Mycobacterium tuberculosis. Trends Microbiol. 1997, 5, 367–372. [Google Scholar] [CrossRef]
- Shastri, M.D.; Shukla, S.D.; Chong, W.C.; Dua, K.; Peterson, G.M.; Patel, R.P.; Hansbro, P.M.; Eri, R.; O’Toole, R.F. Role of Oxidative Stress in the Pathology and Management of Human Tuberculosis. Oxidative Med. Cell. Longev. 2018, 2018, 7695364. [Google Scholar] [CrossRef] [Green Version]
- Koshkin, A.; Nunn, C.M.; Djordjevic, S.; Ortiz de Montellano, P.R. The Mechanism of Mycobacterium tuberculosis Alkylhydroperoxidase AhpD as Defined by Mutagenesis, Crystallography, and Kinetics. J. Biol. Chem. 2003, 278, 29502–29508. [Google Scholar] [CrossRef] [Green Version]
- Sherman, D.R.; Mdluli, K.; Hickey, M.J.; Barry, C.E.; Stover, C.K. AhpC, oxidative stress and drug resistance in Mycobacterium tuberculosis. BioFactors 1999, 10, 211–217. [Google Scholar] [CrossRef]
- Jaeger, T. Peroxiredoxin Systems in Mycobacteria. Subcell. Biochem. 2007, 44, 207–217. [Google Scholar] [CrossRef]
- Bryk, R.; Lima, C.D.; Bromage, H.E.; Tempst, P.; Nathan, C. Metabolic Enzymes of mycobacteria linked to antioxidant defense by a Thioredoxin-Like Protein. Science 2022, 295, 1073–1077. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.; De Lisle, G.W.; Marcinkeviciene, J.A.; Blanchard, J.S.; Collins, D.M. Antisense RNA to AhpC, an Oxidative Stress Defence Gene Involved in Isoniazid Resistance, Indicates That AhpC of Mycobacterium Bovis Has Virulence Properties. Microbiology 1998, 144, 2687–2695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Master, S.S.; Springer, B.; Sander, P.; Boettger, E.C.; Deretic, V.; Timmins, G.S. Oxidative Stress Response Genes in Mycobacterium tuberculosis: Role of AhpC in Resistance to Peroxynitrite and Stage-Specific Survival in Macrophages. Microbiology 2002, 148, 3139–3144. [Google Scholar] [CrossRef] [Green Version]
- Venkatesan, A.; Palaniyandi, K.; Sharma, D.; Bisht, D.; Narayanan, S. Functional Characterization of PknI-Rv2159c Interaction in Redox Homeostasis of Mycobacterium tuberculosis. Front. Microbiol. 2016, 7, 1654. [Google Scholar] [CrossRef] [Green Version]
- Golby, P.; Hatch, K.A.; Bacon, J.; Cooney, R.; Riley, P.; Allnutt, J.; Hinds, J.; Nunez, J.; Marsh, P.D.; Hewinson, R.G.; et al. Comparative transcriptomics reveals key gene expression differences between the human and bovine pathogens of the Mycobacterium tuberculosis complex. Microbiology 2007, 153, 3323–3336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopalaswamy, R.; Narayanan, S.; Chen, B.; Jacobs, W.R.; Av-Gay, Y. The Serine/Threonine Protein Kinase PknI Controls the Growth of Mycobacterium tuberculosis upon Infection. FEMS Microbiol. Lett. 2009, 295, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Venkatesan, A.; Hassan, S.; Palaniyandi, K.; Narayanan, S. In silico and experimental validation of protein-protein interactions between PknI and Rv2159c from Mycobacterium tuberculosis. J. Mol. Graph. Model. 2015, 62, 283–293. [Google Scholar] [CrossRef]
- Bhargavi, G.; Hassan, S.; Balaji, S.; Tripathy, S.P.; Palaniyandi, K. Protein-Protein Interaction of Rv0148 with Htdy and Its Predicted Role towards Drug Resistance in Mycobacterium tuberculosis. BMC Microbiol. 2020, 20, 93. [Google Scholar] [CrossRef] [Green Version]
- Bhargavi, G.; Singh, A.K.; Patil, S.A.; Palaniyandi, K. A putative short-chain dehydrogenase Rv0148 of Mycobacterium tuberculosis affects bacterial survival and virulence. Curr. Res. Microb. Sci. 2021, 3, 100113. [Google Scholar] [CrossRef]
- Rodriguez, M.G.; Neyrolles, O. Metallobiology of Tuberculosis. Microbiol. Spectr. 2014, 2, 2–3. [Google Scholar] [CrossRef] [Green Version]
- Neyrolles, O.; Mintz, E.; Catty, P. Zinc and Copper Toxicity in Host Defense against Pathogens: Mycobacterium tuberculosis as a Model Example of an Emerging Paradigm. Front. Cell. Infect. Microbiol. 2013, 3, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neyrolles, O.; Wolschendorf, F.; Mitra, A.; Niederweis, M. Mycobacteria, Metals, and the Macrophage. Immunol. Rev. 2015, 264, 249–263. [Google Scholar] [CrossRef]
- Nambi, S.; Long, J.K.; Mishra, B.B.; Baker, R.; Murphy, K.C.; Olive, A.J.; Nguyen, H.P.; Shaffer, S.A.; Sassetti, C.M. The oxidative stress network of Mycobacterium tuberculosis reveals coordination between radical detoxification systems. Cell Host Microbe 2016, 17, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Tullius, M.V.; Harth, G.; Horwitz, M.A. High Extracellular Levels of Mycobacterium tuberculosis Glutamine Synthetase and Superoxide Dismutase in Actively Growing Cultures Are Due to High Expression and Extracellular Stability Rather than to a Protein-Specific Export Mechanism. Infect. Immun. 2001, 69, 6348–6363. [Google Scholar] [CrossRef] [Green Version]
- Liao, D.; Fan, Q.; Bao, L. The role of superoxide dismutase in the survival of Mycobacterium tuberculosis in macrophages. Jpn. J. Infect. Dis. 2003, 66, 480–488. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Liu, H.; Shi, L.; Gan, M.; Zhao, X.; Lyu, L.D.; Takiff, H.E.; Wan, K.; Gao, Q. Local adaptation of Mycobacterium tuberculosis on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2021, 118, e2017831118. [Google Scholar] [CrossRef]
- Hillas, P.J.; Alba, F.S.; Oyarzabal, J.; Wilks, A.; de Montellano, P.R.O. The AhpC and AhpD Antioxidant Defense System of Mycobacterium tuberculosis. J. Biol. Chem. 2000, 275, 18801–18809. [Google Scholar] [CrossRef] [Green Version]
- Voskuil, M.I.; Bartek, I.L.; Visconti, K.; Schoolnik, G.K. The Response of Mycobacterium tuberculosis to Reactive Oxygen and Nitrogen Species. Front. Microbiol. 2011, 2, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.N.; Lee, N.O.; Han, S.J.; Ko, I.J.; Oh, J. Il Regulation of the AhpC Gene Encoding Alkyl Hydroperoxide Reductase in Mycobacterium Smegmatis. PLoS ONE 2014, 9, e111680. [Google Scholar] [CrossRef]
- Seaver, L.C.; Imlay, J.A. Alkyl Hydroperoxide Reductase Is the Primary Scavenger of Endogenous Hydrogen Peroxide in Escherichia Coli. J. Bacteriol. 2001, 183, 7173–7181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alan, R.; Katritzky, C.W.R. (Eds.) Comprehensive Organic Functional Group Transformations ScienceDirect. In Comprehensive Organic Functional Group Transformations; Elsevier: Amsterdam, The Netherlands, 1995; ISBN 978-0-08-044705-6. [Google Scholar]
- Agranoff, D.; Krishna, S. Metal Ion Transport and Regulation in Mycobacterium tuberculosis. Front. Biosci. 2004, 9, 2996–3006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- León-Torres, A.; Arango, E.; Castillo, E.; Soto, C.Y. CtpB Is a Plasma Membrane Copper (I) Transporting P-Type ATPase of Mycobacterium tuberculosis. Biol. Res. 2020, 53, 6. [Google Scholar] [CrossRef] [PubMed]
- Achard, M.E.S.; Stafford, S.L.; Bokil, N.J.; Chartres, J.; Bernhardt, P.V.; Schembri, M.A.; Sweet, M.J.; McEwan, A.G. Copper Redistribution in Murine Macrophages in Response to Salmonella Infection. Biochem. J. 2012, 444, 51–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.R.; Liu, J.; Wang, Y.; Xian, Q.Y.; Shao, L.Y.; Yang, Z.; Wang, X.N. Immunotherapy Using IL-2 and GM-CSF Is a Potential Treatment for Multidrug-Resistant Mycobacterium tuberculosis. Sci. China Life Sci. 2012, 55, 800–806. [Google Scholar] [CrossRef]
- Benmerzoug, S.; Marinho, F.V.; Rose, S.; Mackowiak, C.; Gosset, D.; Sedda, D.; Poisson, E.; Uyttenhove, C.; Van Snick, J.; Jacobs, M.; et al. GM-CSF Targeted Immunomodulation Affects Host Response to M. tuberculosis Infection. Sci. Rep. 2018, 8, 8652. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, R.D.; Prince, O.; Cooper, A.; Khader, S. Cytokines and Chemokines in Mycobacterium tuberculosis Infection. Microbiol. Spectr. 2016, 4, 4–5. [Google Scholar] [CrossRef] [Green Version]
- Ruhwald, M.; Bodmer, T.; Maier, C.; Jepsen, M.; Haaland, M.B.; Eugen-Olsen, J.; Ravn, P. Evaluating the Potential of IP-10 and MCP-2 as Biomarkers for the Diagnosis of Tuberculosis. Eur. Respir. J. 2008, 32, 1607–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruhwald, M.; Aabye, M.G.; Ravn, P. IP-10 Release Assays in the Diagnosis of Tuberculosis Infection: Current Status and Future Directions. Expert Rev. Mol. Diagn. 2012, 12, 175–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilda, J.N.; Narasimhan, M.; Das, S.D. Neutrophils from Pulmonary Tuberculosis Patients Show Augmented Levels of Chemokines MIP-1α, IL-8 and MCP-1 Which Further Increase upon in Vitro Infection with Mycobacterial Strains. Hum. Immunol. 2014, 75, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Mehta, N.S.; Benoit, S.L.; Mysore, J.; Maier, R.J. In Vitro and in Vivo Characterization of Alkyl Hydroperoxide Reductase Mutant Strains of Helicobacter hepaticus. Biochim. Biophys. Acta -Gen. Subj. 2007, 1770, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Charoenlap, N.; Shen, Z.; Mcbee, M.E.; Muthupalani, S.; Wogan, G.N.; Fox, J.G.; Schauer, D.B. Alkyl Hydroperoxide Reductase Is Required for Helicobacter cinaedi Intestinal Colonization and Survival under Oxidative Stress in Balb/c and Balb/c Interleukin-10-/- Mice. Infect. Immun. 2012, 80, 921–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Springer, B.; Master, S.; Sander, P.; Zahrt, T.; McFalone, M.; Song, J.; Papavinasasundaram, K.G.; Colston, M.J.; Boettger, E.; Deretic, V. Silencing of oxidative stress response in Mycobacterium tuberculosis: Expression patterns of ahpC in virulent and avirulent strains and effect of ahpC inactivation. Infect Immun. 2001, 69, 5967–5973. [Google Scholar] [CrossRef] [Green Version]
- Clark, S.; Hall, Y.; Williams, A. Animals Models of Tuberculosis: Guinea Pigs. Cold Spring Harb. Perspect. Med. 2015, 5, a018572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, P.; Reddy, P.V.; Singh, R.; Jaisinghani, N.; Gandotra, S.; Tyagi, A.K. Secretory Phosphatases Deficient Mutant of Mycobacterium tuberculosis Imparts Protection at the Primary Site of Infection in Guinea Pigs. PLoS ONE 2013, 8, e77930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chawla, M.; Parikh, P.; Saxena, A.; Munshi, M.; Mehta, M.; Mai, D.; Srivastava, A.K.; Narasimhulu, K.V.; Redding, K.E.; Vashi, N.; et al. Mycobacterium tuberculosis WhiB4 Regulates Oxidative Stress Response to Modulate Survival and Dissemination in Vivo. Mol. Microbiol. 2012, 85, 1148–1165. [Google Scholar] [CrossRef]
- Jain, P.; Hsu, T.; Arai, M.; Biermann, K.; Thaler, D.S.; Nguyen, A.; González, P.A.; Tufariello, J.M.; Kriakov, J.; Chen, B.; et al. Specialized transduction designed for precise high-throughput unmarked deletions in Mycobacterium tuberculosis. mBio 2014, 5, 01245-14. [Google Scholar] [CrossRef] [Green Version]
- Vandal, O.H.; Roberts, J.A.; Odaira, T.; Schnappinger, D.; Nathan, C.F.; Ehrt, S. Acid-Susceptible mutants of Mycobacterium tuberculosis shares hypersusceptibility to cell wall and oxidative stress and to the host environment. J. Bacteriol. 2009, 191, 625–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, K.N.; Verma, A.K.; Gopalaswamy, R.; Adikesavalu, H.; Singhal, D.K.; Tripathy, S.; Ranganathan, U.D.; Sherman, D.R.; Urdahl, K.B.; Ramakrishnan, L.; et al. Diverse Clinical Isolates of Mycobacterium tuberculosis Develop Macrophage-Induced Rifampin Tolerance. J. Infect. Dis. 2019, 219, 1554–1558. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhargavi, G.; Singh, A.K.; Deenadayalan, A.; Ponnuraja, C.; Patil, S.A.; Palaniyandi, K. Role of a Putative Alkylhydroperoxidase Rv2159c in the Oxidative Stress Response and Virulence of Mycobacterium tuberculosis. Pathogens 2022, 11, 684. https://doi.org/10.3390/pathogens11060684
Bhargavi G, Singh AK, Deenadayalan A, Ponnuraja C, Patil SA, Palaniyandi K. Role of a Putative Alkylhydroperoxidase Rv2159c in the Oxidative Stress Response and Virulence of Mycobacterium tuberculosis. Pathogens. 2022; 11(6):684. https://doi.org/10.3390/pathogens11060684
Chicago/Turabian StyleBhargavi, Gunapati, Amit Kumar Singh, Anbarasu Deenadayalan, Chinnaiyan Ponnuraja, Shripad A. Patil, and Kannan Palaniyandi. 2022. "Role of a Putative Alkylhydroperoxidase Rv2159c in the Oxidative Stress Response and Virulence of Mycobacterium tuberculosis" Pathogens 11, no. 6: 684. https://doi.org/10.3390/pathogens11060684
APA StyleBhargavi, G., Singh, A. K., Deenadayalan, A., Ponnuraja, C., Patil, S. A., & Palaniyandi, K. (2022). Role of a Putative Alkylhydroperoxidase Rv2159c in the Oxidative Stress Response and Virulence of Mycobacterium tuberculosis. Pathogens, 11(6), 684. https://doi.org/10.3390/pathogens11060684