
Critically Important Antimicrobial Resistance Trends in Salmonella Derby and Salmonella Typhimurium Isolated from the Pork Production Chain in Brazil: A 16-Year Period

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Resistance Trends
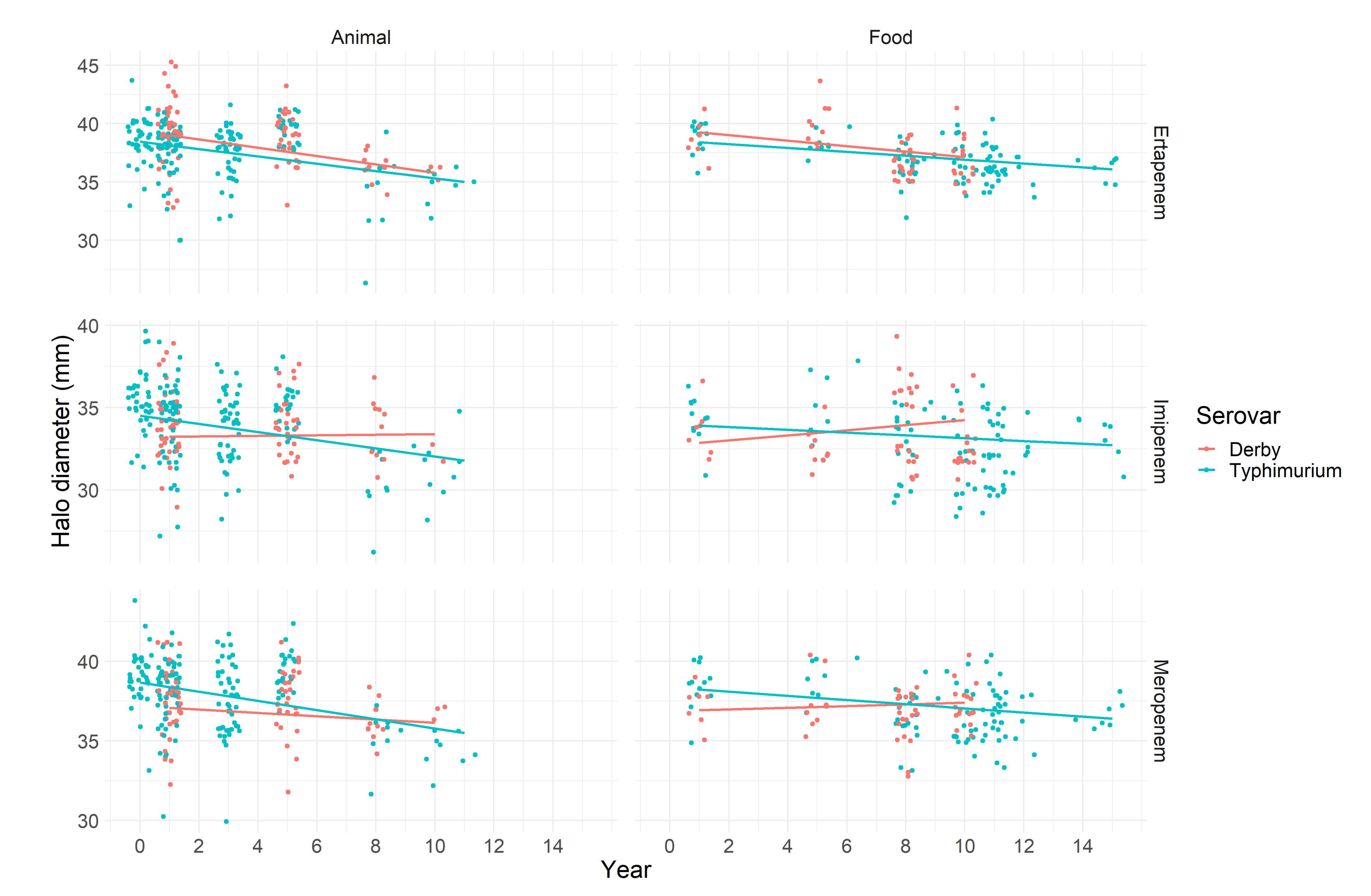
2.1.1. Carbapenems
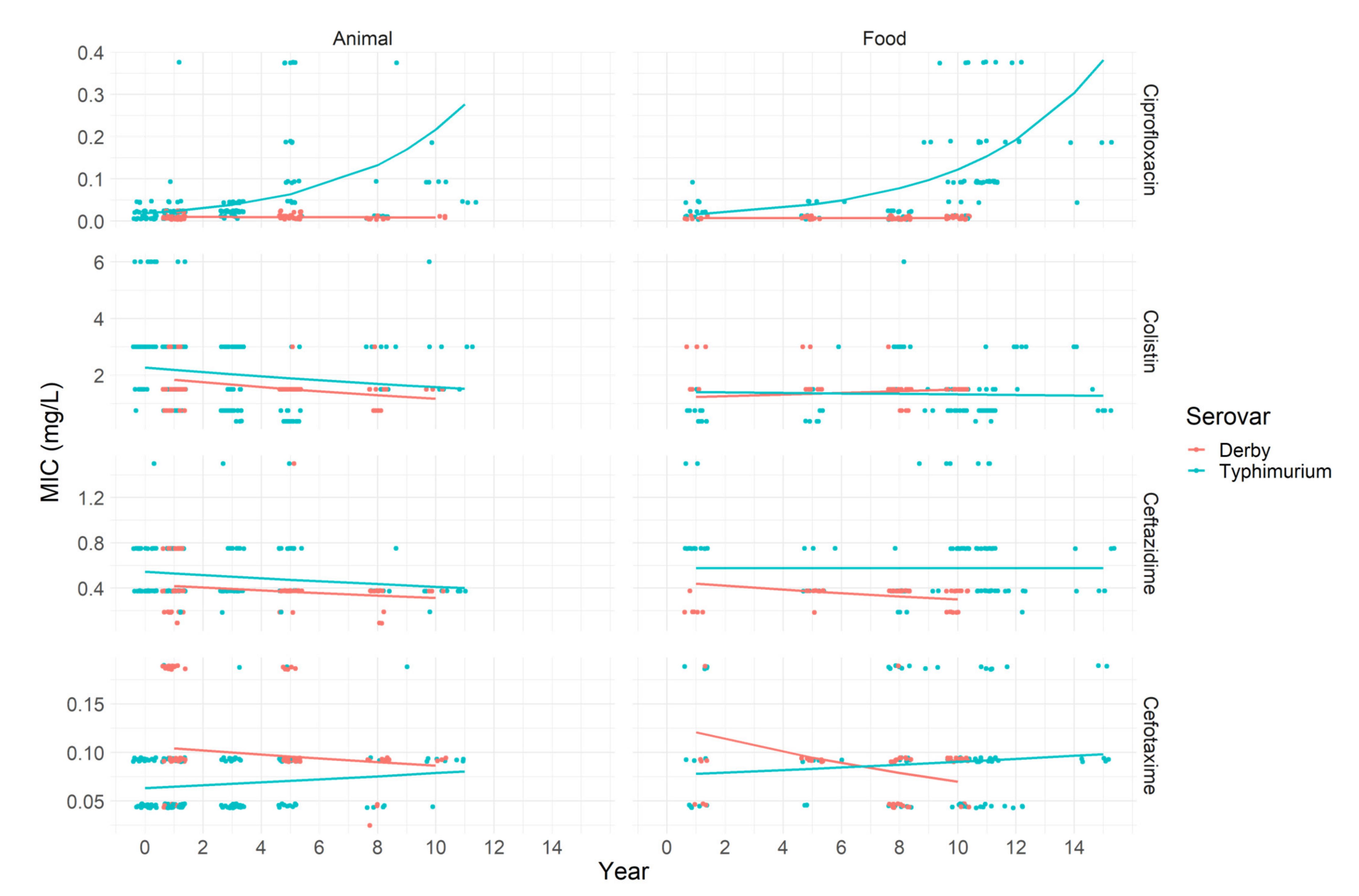
2.1.2. Third-Generation Cephalosporins, Ciprofloxacin and Colistin
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Antimicrobial Resistance Test
4.3. Statistical Analysis
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
References and Notes
- WHO. Critically Important Antimicrobials, 6th ed.; World Health Organization: Geneva, Switzerland, 2019; ISBN 9789241515528. [Google Scholar]
- Palma, E.; Tilocca, B.; Roncada, P. Antimicrobial Resistance in Veterinary Medicine: An Overview. Int. J. Mol. Sci. 2020, 21, 1914. [Google Scholar] [CrossRef]
- Cassini, A.; Colzani, E.; Pini, A.; Mangen, M.J.J.; Plass, D.; McDonald, S.A.; Maringhini, G.; van Lier, A.; Haagsma, J.A.; Havelaar, A.H.; et al. Impact of Infectious Diseases on Population Health Using Incidence-Based Disability-Adjusted Life Years (DALYs): Results from the Burden of Communicable Diseases in Europe Study, European Union and European Economic Countries, 2009 to 2013. Eurosurveillance 2018, 23, 17–00454. [Google Scholar] [CrossRef]
- Evers, E.G.; Pielaat, A.; Smid, J.H.; van Duijkeren, E.; Vennemann, F.B.C.; Wijnands, L.M.; Chardon, J.E. Comparative Exposure Assessment of Esblproducing Escherichia coli through Meat Consumption. PLoS ONE 2017, 12, e0169589. [Google Scholar] [CrossRef]
- Huijbers, P.M.C.; Blaak, H.; de Jong, M.C.M.; Graat, E.A.M.; Vandenbroucke-Grauls, C.M.J.E.; de Roda Husman, A.M. Role of the Environment in the Transmission of Antimicrobial Resistance to Humans: A Review. Environ. Sci. Technol. 2015, 49, 11993–12004. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Neto, W.S.; Leotti, V.B.; Pires, S.M.; Hald, T.; Corbellini, L.G. Non-Typhoidal Human Salmonellosis in Rio Grande Do Sul, Brazil: A Combined Source Attribution Study of Microbial Subtyping and Outbreak Data. Int. J. Food Microbiol. 2021, 338, 108992. [Google Scholar] [CrossRef] [PubMed]
- EFSA. The European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, e05926. [Google Scholar] [CrossRef]
- Finger, J.A.F.F.; Baroni, W.S.G.V.; Maffei, D.F.; Bastos, D.H.M.; Pinto, U.M. Overview of Foodborne Disease Outbreaks in Brazil from 2000 to 2018. Foods 2019, 8, 434. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne Illness Acquired in the United States-Major Pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef]
- de Freitas Costa, E.; Cardoso, M.; Kich, J.D.; Corbellini, L.G. A Qualitative Risk Assessment Approach to Microbial Foodborne Hazards in Brazilian Intensive Pork Production: A Step towards Risk Prioritization. Microb. Risk Anal. 2020, 15, 100105. [Google Scholar] [CrossRef]
- Kich, J.D.; Coldebella, A.; Morés, N.; Nogueira, M.G.; Cardoso, M.; Fratamico, P.M.; Call, J.E.; Fedorka-Cray, P.; Luchansky, J.B. Prevalence, Distribution, and Molecular Characterization of Salmonella Recovered from Swine Finishing Herds and a Slaughter Facility in Santa Catarina, Brazil. Int. J. Food Microbiol. 2011, 151, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Pissetti, C.; Werlang, G.O.; Biesus, L.L.; Kich, J.D.; Cardoso, M.R. Salmonella enterica and Listeria monocytogenes Detection on Pre-Chill Pig Carcasses. Acta Sci. Vet. 2012, 40, 1071. [Google Scholar]
- Silva, L.E.; Dias, V.; Ferronatto, A.; Guerra, P.; Berno, L.; Triches, N.; Kich, J.D.; Corbellini, L.G.; Cardoso, M. Longitudinal Dissemination of Salmonella enterica Clonal Groups through the Slaughter Process of Salmonella-Positive Pig Batches. J. Food Prot. 2012, 75, 1580–1588. [Google Scholar] [CrossRef]
- Bersot, L.D.S.; Cavicchioli, V.Q.; Viana, C.; Burin, R.C.K.; Camargo, A.C.; de Almeida Nogueira Pinto, J.P.; Nero, L.A.; Destro, M.T. Prevalence, Antimicrobial Resistance, and Diversity of Salmonella along the Pig Production Chain in Southern Brazil. Pathogens 2019, 8, 204. [Google Scholar] [CrossRef] [PubMed]
- Paim, D.S.; Pissetti, C.; Vieira, T.R.; Werlang, G.O.; de Freitas Costa, E.; Kich, J.D.; Cardoso, M. Enumeration, Antimicrobial Resistance and Typing of Salmonella enterica: Profile of Strains Carried in the Intestinal Contents of Pigs at Slaughter in Southern Brazil. Acta Sci. Vet. 2019, 47, 1636. [Google Scholar] [CrossRef]
- Rodrigues, G.L.; Panzenhagen, P.; Ferrari, R.G.; Paschoalin, V.M.F.; Conte-Junior, C.A. Antimicrobial Resistance in Nontyphoidal Salmonella Isolates from Human and Swine Sources in Brazil: A Systematic Review of the Past Three Decades. Microb. Drug Resist. 2020, 26, 1260–1270. [Google Scholar] [CrossRef]
- Lopes, G.V.; Pissetti, C.; da Cruz Payão Pellegrini, D.É.B.O.R.A.; Da Silva, L.E.; Cardoso, M. Resistance Phenotypes and Genotypes of Salmonella enterica Subsp. enterica Isolates from Feed, Pigs, and Carcasses in Brazil. Brazil. J. Food Prot. 2015, 78, 407–413. [Google Scholar] [CrossRef]
- Morales, A.S.; Fragoso De Araújo, J.; de Moura Gomes, V.T.; Reis Costa, A.T.; dos Prazeres Rodrigues, D.; Porfida Ferreira, T.S.; de Lima Filsner, P.H.N.; Felizardo, M.R.; Micke Moreno, A. Colistin Resistance in Escherichia coli and Salmonella enterica Strains Isolated from Swine in Brazil. Sci. World J. 2012, 2012, 109795. [Google Scholar] [CrossRef]
- Duarte, A.S.R.; Høg, B.B.; Korsgaard, H.B.; Attauabi, M.; Boel, J.; Dalby, T.; Hammerum, A.M.; Hansen, F.; Hasman, H.; Henius, A.E. DANMAP Use of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Bacteria from Food Animals, Food and Humans in Denmark (DANMAP 2020); Statens Serum Institut and Technical University of Denmark: Copenhagen, Denmark, 2021; pp. 1–174. [Google Scholar]
- EFSA. The Community Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Animals and Food in the European Union in 2004–2007. EFSA J. 2010, 8, 1309. [Google Scholar] [CrossRef]
- Medalla, F.; Hoekstra, R.M.; Whichard, J.M.; Barzilay, E.J.; Chiller, T.M.; Joyce, K.; Rickert, R.; Krueger, A.; Stuart, A.; Griffin, P.M. Increase in Resistance to Ceftriaxone and Nonsusceptibility to Ciprofloxacin and Decrease in Multidrug Resistance among Salmonella Strains, United States, 1996–2009. Foodborne Pathog. Dis. 2013, 10, 302–309. [Google Scholar] [CrossRef]
- Angelo, K.M.; Reynolds, J.; Karp, B.E.; Hoekstra, R.M.; Scheel, C.M.; Friedman, C. Antimicrobial Resistance among Nontyphoidal Salmonella Isolated from Blood in the United States, 2003–2013. J. Infect. Dis. 2016, 214, 1565–1570. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Biswas, S.; Paudyal, N.; Pan, H.; Li, X.; Fang, W.; Yue, M. Antibiotic Resistance in Salmonella Typhimurium Isolates Recovered from the Food Chain through National Antimicrobial Resistance Monitoring System between 1996 and 2016. Front. Microbiol. 2019, 10, 985. [Google Scholar] [CrossRef] [PubMed]
- MAPA. Ministério da Agricultura, Pecuária e Abastecimento. Programa de Vigilância e Monitoramento Da Resistência Aos Antimicrobianos No Âmbito Da Agropecuária; Ministério da Agricultura, Pecuária e Abastecimento: Brasília, Brasil, 2021. [Google Scholar]
- EUCAST. European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. 2022, 110p. Available online: http://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_5.0_Breakpoint_Table_01.pdf (accessed on 5 July 2022).
- EUCAST. MIC and Zone Diameter Distributions and ECOFFs. Available online: https://www.eucast.org/mic_distributions_and_ecoffs/ (accessed on 5 July 2022).
- EUCAST. Guidelines for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance. 2013, 1, 1–40
- EUCAST. Guidelines for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance. 2017, 2, 1–43
- Mürmann, L.; dos Santos, M.C.; Cardoso, M. Prevalence, Genetic Characterization and Antimicrobial Resistance of Salmonella Isolated from Fresh Pork Sausages in Porto Alegre, Brazil. Food Control 2009, 20, 191–195. [Google Scholar] [CrossRef]
- Viana, C.; Sereno, M.J.; Pegoraro, K.; Yamatogi, R.S.; Call, D.R.; dos Santos Bersot, L.; Nero, L.A. Distribution, Diversity, Virulence Genotypes and Antibiotic Resistance for Salmonella Isolated from a Brazilian Pork Production Chain. Int. J. Food Microbiol. 2019, 310, 108310. [Google Scholar] [CrossRef] [PubMed]
- Almeida, F.; Medeiros, M.I.C.; Kich, J.D.; Falcão, J.P. Virulence-Associated Genes, Antimicrobial Resistance and Molecular Typing of Salmonella Typhimurium Strains Isolated from Swine from 2000 to 2012 in Brazil. J. Appl. Microbiol. 2016, 120, 1677–1690. [Google Scholar] [CrossRef] [PubMed]
- Diallo, O.O.; Baron, S.A.; Abat, C.; Colson, P.; Chaudet, H.; Rolain, J.M. Antibiotic Resistance Surveillance Systems: A Review. J. Glob. Antimicrob. Resist. 2020, 23, 430–438. [Google Scholar] [CrossRef]
- Hoinville, L.J.; Alban, L.; Drewe, J.A.; Gibbens, J.C.; Gustafson, L.; Häsler, B.; Saegerman, C.; Salman, M.; Stärk, K.D.C. Proposed Terms and Concepts for Describing and Evaluating Animal-Health Surveillance Systems. Prev. Vet. Med. 2013, 112, 1–12. [Google Scholar] [CrossRef]
- Brasil Instrução Normativa N° 45, 22 November 2016. Ministério da Agricultura, Pecuária e Abastecimento: 2016
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Schwarz, S.; Johnson, A.P. Transferable Resistance to Colistin: A New but Old Threat. J. Antimicrob. Chemother. 2016, 71, 2066–2070. [Google Scholar] [CrossRef]
- Leite, E.L.; Araújo, W.J.; Vieira, T.R.; Zenato, K.S.; Vasconcelos, P.C.; Cibulski, S.; Givisiez, P.E.N.; Cardoso, M.R.I.; Oliveira, C.J.B. First Reported Genome of an Mcr-9-Mediated Colistin-Resistant Salmonella Typhimurium Isolate from Brazilian Livestock. J. Glob. Antimicrob. Resist. 2020, 23, 394–397. [Google Scholar] [CrossRef]
- Rhouma, M.; Beaudry, F.; Thériault, W.; Letellier, A. Colistin in Pig Production: Chemistry, Mechanism of Antibacterial Action, Microbial Resistance Emergence, and One Health Perspectives. Front. Microbiol. 2016, 7, 1789. [Google Scholar] [CrossRef] [PubMed]
- Gomes, V.T.M.; Moreno, L.Z.; Silva, A.P.S.; Thakur, S.; la Ragione, R.M.; Mather, A.E.; Moreno, A.M. Characterization of Salmonella enterica Contamination in Pork and Poultry Meat from São Paulo/Brazil: Serotypes, Genotypes and Antimicrobial Resistance Profiles. Pathogens 2022, 11, 358. [Google Scholar] [CrossRef] [PubMed]
- Sato, J.; Vannucci, F.; Gava, D.; Barcellos, D. O Uso Prudente e Eficaz de Antibióticos Na Suinocultura: Uma Abordagem Integrada; ABCS: Brasília, Brazil, 2022; ISBN 9788568384121. [Google Scholar]
- Zimmerman, J.; Karriker, L.; Ramirez, A.; Schwartz, K.; Stevenson, G.Z.J. Disease of Swine; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; Volume 11, ISBN 9781119350903. [Google Scholar]
- Fleury, M.A.; Mourand, G.; Jouy, E.; Touzain, F.; le Devendec, L.; de Boisseson, C.; Eono, F.; Cariolet, R.; Guérin, A.; le Goff, O.; et al. Impact of Ceftiofur Injection on Gut Microbiota and Escherichia coli Resistance in Pigs. Antimicrob. Agents Chemother. 2015, 59, 5171–5180. [Google Scholar] [CrossRef] [PubMed]
- Pissetti, C.; Kich, J.D.; Allen, H.K.; Navarrete, C.; de Freitas Costa, E.; Morés, N.; Cardoso, M. Antimicrobial Resistance in Commensal Escherichia coli and Enterococcus Spp. Isolated from Pigs Subjected to Different Antimicrobial Administration Protocols. Res. Vet. Sci. 2021, 137, 174–185. [Google Scholar] [CrossRef]
- Fernández, J.; Guerra, B.; Rodicio, M.R. Resistance to Carbapenems in Non-Typhoidal Salmonella enterica Serovars from Humans, Animals and Food. Vet. Sci. 2018, 5, 40. [Google Scholar] [CrossRef]
- Abraham, S.; O’Dea, M.; Trott, D.J.; Abraham, R.J.; Hughes, D.; Pang, S.; McKew, G.; Cheong, E.Y.L.; Merlino, J.; Saputra, S.; et al. Isolation and Plasmid Characterization of Carbapenemase (IMP-4) Producing Salmonella enterica Typhimurium from Cats. Sci. Rep. 2016, 6, 35527. [Google Scholar] [CrossRef]
- Associação Brasileira de Proteína Animal (ABPA). Relatório Anual 2022; S;ão Paulo, Brazil, 2022.
- Clark, J.S.; Bjornstad, O. Population Time Series with Errors, Missing Values, and Time Lags. Ecol. Soc. Am. Annu. Meet. Abstr. 2004, 89, 96. [Google Scholar]
- Olvera Astivia, O.L.; Gadermann, A.; Guhn, M. The Relationship between Statistical Power and Predictor Distribution in Multilevel Logistic Regression: A Simulation-Based Approach. BMC Med. Res. Methodol. 2019, 19, 97. [Google Scholar] [CrossRef]
- Bessa, M.C.; da Costa, M.; Cardoso, M. Prevalência de Salmonella Sp. Em Suínos Abatidos Em Frígoríficos Do Rio Grande Do Sul. Pesqui. Veterinária Bras. 2004, 24, 80–84. [Google Scholar] [CrossRef]
- Castagna, S.M.F.; Schwarz, P.; Canal, C.W.; Cardoso, M. Presença de Salmonella Sp. No Trato Intestinal e Em Tonsilas/Linfonodos Submandibulares de Suínos Ao Abate. Arq. Bras. De Med. Veterinária E Zootec. 2004, 56, 300–306. [Google Scholar] [CrossRef]
- Castagna, S.M.F.; Schwarz, P.; Canal, C.W.; Cardoso, M.R. DeItapema Prevalência de Suínos Portadores de Salmonella Sp. Ao Abate e Contaminação de Embutidos Tipo Frescal. Acta Sci. Vet. 2004, 32, 141. [Google Scholar] [CrossRef]
- Michael, G.B.; Simoneti, R.; Cardoso, M.R.D.I.; Costa, M.D. Sorotipos de Salmonella Isolados Em Uma Propriedade de Suínos de Terminação No Sul Do Brasil. Ciência Rural 2002, 32, 525–527. [Google Scholar] [CrossRef]
- Silva, L.E.; Gotardi, C.P.; Vizzotto, R.; Kich, J.D.; Cardoso, M.R.I. Infecção Por Salmonella enterica Em Suínos Criados Em Um Sistema Integrado de Produção Do Sul Do Brasil Salmonella Infection in Pigs Raised in a Multiple-Site Swine Production System from Southern Brazil. Arq. Bras. Med. Vet. Zootec 2006, 58, 455–461. [Google Scholar] [CrossRef]
- Schwarz, P.; Kich, J.D.; Kolb, J.; Cardoso, M. Use of an Avirulent Live Salmonella choleraesuis Vaccine to Reduce the Prevalence of Salmonella Carrier Pigs at Slaughter. Vet. Rec. 2011, 169, 553. [Google Scholar] [CrossRef] [PubMed]
- Werlang, G.O.; Haubert, L.; Peter, C.M.; Cardoso, M. Isolation of Salmonella Typhimurium, Listeria Monocytogenes and Coagulase-Positive Staphylococcus from Salami Sold at Street Fairs in Porto Alegre, Brazil. Arq. Inst. Biol. 2019, 86, 1–6. [Google Scholar] [CrossRef]
- Werlang, G.O.; Kich, J.D.; Lopes, G.V.; Coldebella, A.; Feddern, V.; Cardoso, M. Effect of Gaseous Ozone Application during Chilling on Microbial and Quality Attributes of Pig Carcasses. Food Sci. Technol. Int. 2021, 28, 366–376. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- Bjork, K.E.; Kopral, C.A.; Wagner, B.A.; Dargatz, D.A. Comparison of Mixed Effects Models of Antimicrobial Resistance Metrics of Livestock and Poultry Salmonella Isolates from a National Monitoring System. Prev. Vet. Med. 2015, 122, 265–272. [Google Scholar] [CrossRef]
- Kalbfleisch, J.D.; Prentice, R.L. The Statistical Analysis of Failure Time Data; Wiley: Hoboken, NJ, USA, 2002. [Google Scholar]
- R Core Team, R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model; Springer: New York, NY, USA, 2000; ISBN 0-387-98784-3. [Google Scholar]
- Therneau, T. A Package for Survival Analysis in R. R Package Version 3.4-0. Available online: https://cran.r-project.org/package=survival (accessed on 30 June 2022).



{kind=link}
{kind=link}
| Variable 1 | Estimate | 95% CI 2 | p-Value | ICC 3 |
|---|---|---|---|---|
| Ertapenem: | 19.41% | |||
| 39.36 | 38.16; 40.53 | - | ||
| 0.91 | 0.17; 1.7 | 0.02 | ||
| 0.13 | −1.04; 1.36 | 0.83 | ||
| −0.36 | −0.57; −0.14 | 0.003 | ||
| −0.31 | −0.48; −0.14 | 0.001 | ||
| −0.23 | −0.43; −0.04 | 0.03 | ||
| −0.16 | −0.32; −0.01 | 0.04 | ||
| Imipenem: | 16.8% | |||
| 33.21 | 32.00; 34.39 | - | ||
| −1.3 | −2.03; −0.47 | 0.001 | ||
| −0.50 | −1.70; 0.66 | 0.41 | ||
| 0.02 | −0.21; 0.23 | 0.87 | ||
| −0.24 | −0.42; −0.08 | 0.01 | ||
| 0.15 | −0.05; 0.34 | 0.13 | ||
| −0.08 | −0.23; 0.06 | 0.28 | ||
| Meropenem: | 15.20% | |||
| 37.18 | 36.18; 38.17 | - | ||
| −1.48 | −2.15; −0.77 | <0.001 | ||
| −0.31 | −1.35; 0.75 | 0.57 | ||
| −0.10 | −0.29; 0.08 | 0.29 | ||
| −0.29 | −0.43; −0.14 | <0.001 | ||
| 0.05 | −0.11; 0.22 | 0.54 | ||
| −0.13 | −0.26; −0.002 | 0.06 | ||
| ATM | Source | Serovar | Number of Isolates with MIC (mg/L) Same to: | ECOFF a | R b | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.008 | 0.015 | 0.03 | 0.06 | 0.125 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | (%) | (%) | |||
| Ciprofloxacin | Animal | Derby | 40 | 30 | 12 | 0 | 0 | |||||||||
| Typhimurium | 31 | 29 | 66 | 28 | 11 | 5 | 8 | 2 | 30 | 14.44 | ||||||
| Food | Derby | 37 | 16 | 0 | 0 | |||||||||||
| Typhimurium | 12 | 10 | 13 | 12 | 25 | 13 | 8 | 5 c | 64.29 | 52.04 | ||||||
| Colistin | Animal | Derby | 11 | 61 | 10 | * | 12.20 | |||||||||
| Typhimurium | 15 | 27 | 42 | 85 | 10 | 1 | * | 53.33 | ||||||||
| Food | Derby | 5 | 42 | 6 | * | 11.32 | ||||||||||
| Typhimurium | 10 | 46 | 24 | 17 | 1 | * | 18.37 | |||||||||
| Ceftazidime | Animal | Derby | 4 | 12 | 48 | 17 | 1 | 1.22 | 0 | |||||||
| Typhimurium | 5 | 107 | 64 | 3 | 1 | 2.22 | 0.56 | |||||||||
| Food | Derby | 14 | 39 | 0 | 0 | |||||||||||
| Typhimurium | 5 | 41 | 44 | 8 | 8.16 | 0 | ||||||||||
| Cefotaxime | Animal | Derby | 1 | 7 | 55 | 17 | 1 | 1 | 2.44 | 0 | ||||||
| Typhimurium | 94 | 81 | 5 | 0 | 0 | |||||||||||
| Food | Derby | 16 | 35 | 2 | 0 | 0 | ||||||||||
| Typhimurium | 31 | 47 | 19 | 1 | 1.02 | 0 | ||||||||||
| Variable 1 | Estimate | 95% CI 2 | p-Value |
|---|---|---|---|
| Ciprofloxacin: | |||
| −4 | −4.41; −3.64 | - | |
| −0.56 | −0.81; −0.31 | <0.001 | |
| −0.36 | −0.99; 0.26 | 0.27 | |
| −0.02 | −0.09; 0.05 | 0.62 | |
| 0.25 | 0.06; 0.43 | 0.01 | |
| 0 | −0.07; 0.07 | 0.99 | |
| 0.23 | 0.16; 0.29 | <0.001 | |
| Colistin: | |||
| 0.82 | 0.55; 1.09 | - | |
| −0.16 | −0.73; 0.41 | 0.57 | |
| −0.47 | −0.97; −0.02 | 0.06 | |
| −0.05 | −0.11; 0.01 | 0.1 | |
| −0.03 | −0.11; 0.04 | 0.35 | |
| 0.02 | −0.02; 0.07 | 0.37 | |
| −0.007 | −0.07; 0.06 | 0.83 | |
| Ceftazidime: | |||
| −0.6 | −0.72; −0.52 | - | |
| −0.23 | −0.72; 0.29 | 0.36 | |
| −0.05 | −0.18; 0.32 | 0.38 | |
| −0.03 | −0.11; 0.04 | 0.4 | |
| −0.02 | −0.05; 0.13 | 0.23 | |
| −0.04 | −0.08; 0.001 | 0.06 | |
| −0.0002 | −0.03; 0.03 | 0.99 | |
| Cefotaxime: | |||
| −2.7 | −2.8; −2.6 | - | |
| 0.53 | 0.2; 0.96 | 0.002 | |
| 0.2 | 0.04; 0.35 | 0.017 | |
| −0.02 | −0.08; 0.053 | 0.49 | |
| 0.03 | 0.011; 0.04 | 0.001 | |
| −0.06 | −0.1; −0.02 | 0.003 | |
| 0.017 | −0.001; 0.04 | 0.11 | |
| Collect Year | Number of Isolates—Salmonella | Reference | |
|---|---|---|---|
| Derby | Typhimurium | ||
| 2000 and 2001 | 25 | 37 | Bessa et al., 2004 [50] |
| 2001 | 14 | 30 | Castagna et al., 2004a [51] |
| 2001 | 9 | 29 | Castagna et al., 2004b [52] |
| 2001 | 0 | 7 | Michael et al., 2002 [53] |
| 2003 | 0 | 46 | Silva et al., 2006 [54] |
| 2005 | 28 | 25 | Schwarz et al., 2011 [55] |
| 2005 and 2006 | 9 | 8 | Murmann et al., 2009 [30] |
| 2008 and 2009 | 31 | 30 | Silva et al., 2012 [14] |
| 2010 and 2011 | 19 | 53 | Pissetti et al., 2012 [13] |
| 2012 | 0 | 5 | Werlang et al., 2019 [56] |
| 2014 | 0 | 2 | Werlang et al., 2021 [57] |
| 2015 | 0 | 6 | Paim et al., 2019 [16] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pissetti, C.; de Freitas Costa, E.; Zenato, K.S.; de Itapema Cardoso, M.R. Critically Important Antimicrobial Resistance Trends in Salmonella Derby and Salmonella Typhimurium Isolated from the Pork Production Chain in Brazil: A 16-Year Period. Pathogens 2022, 11, 905. https://doi.org/10.3390/pathogens11080905
Pissetti C, de Freitas Costa E, Zenato KS, de Itapema Cardoso MR. Critically Important Antimicrobial Resistance Trends in Salmonella Derby and Salmonella Typhimurium Isolated from the Pork Production Chain in Brazil: A 16-Year Period. Pathogens. 2022; 11(8):905. https://doi.org/10.3390/pathogens11080905
Chicago/Turabian StylePissetti, Caroline, Eduardo de Freitas Costa, Karoline Silva Zenato, and Marisa Ribeiro de Itapema Cardoso. 2022. "Critically Important Antimicrobial Resistance Trends in Salmonella Derby and Salmonella Typhimurium Isolated from the Pork Production Chain in Brazil: A 16-Year Period" Pathogens 11, no. 8: 905. https://doi.org/10.3390/pathogens11080905
APA StylePissetti, C., de Freitas Costa, E., Zenato, K. S., & de Itapema Cardoso, M. R. (2022). Critically Important Antimicrobial Resistance Trends in Salmonella Derby and Salmonella Typhimurium Isolated from the Pork Production Chain in Brazil: A 16-Year Period. Pathogens, 11(8), 905. https://doi.org/10.3390/pathogens11080905