
Discovery of a Novel Species Infecting Goats: Morphological and Molecular Characterization of Babesia aktasi n. sp.


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Animal Samples
2.2. Determination of New Babesia sp. Infected Goat in Field Samples
2.3. Experimental Study and Monitoring Animals
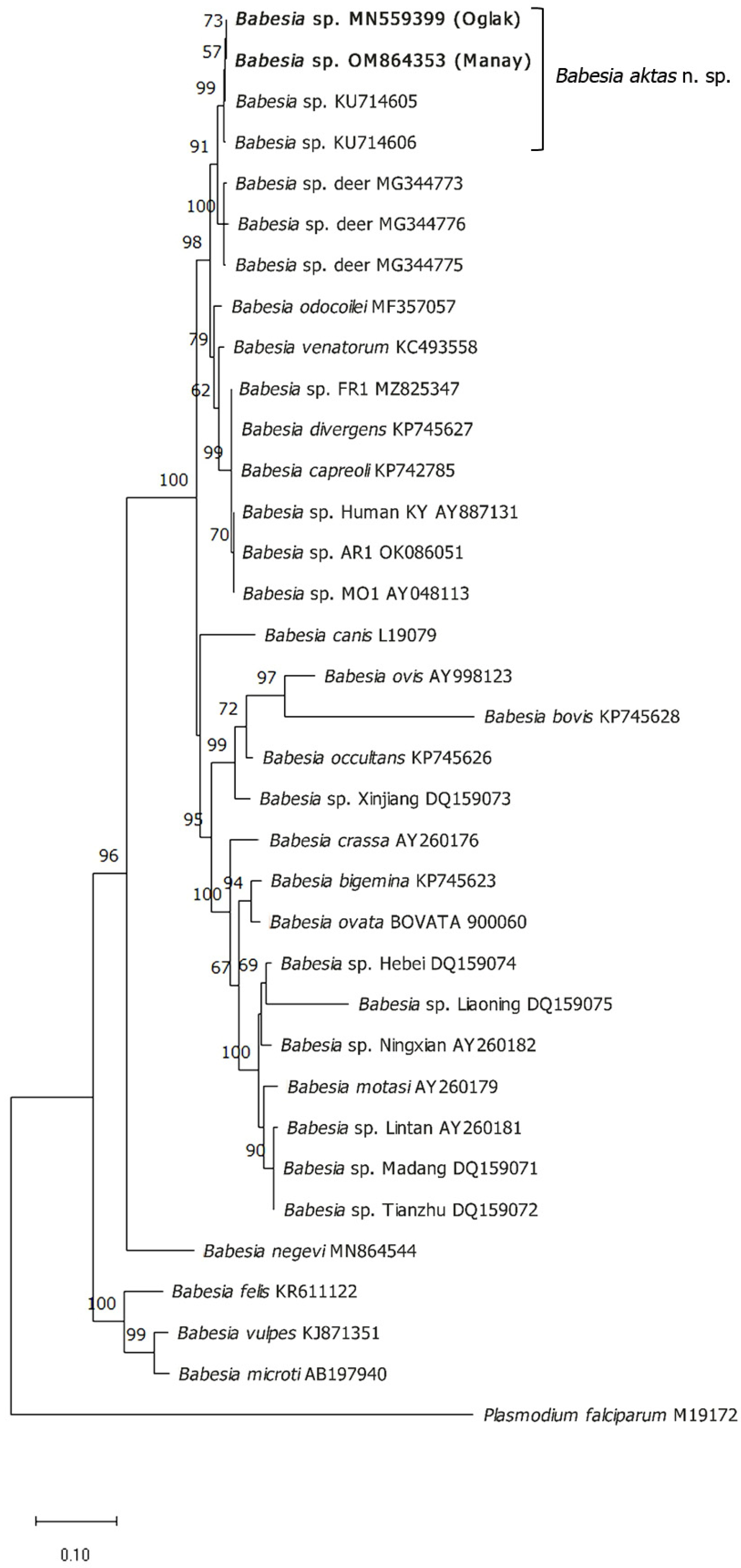
2.4. Phylogenetic and Percent Identity Matrix Analyses

2.5. Ethics Statement
3. Results
3.1. Prevalance Rate of Babesia sp. in Field Samples
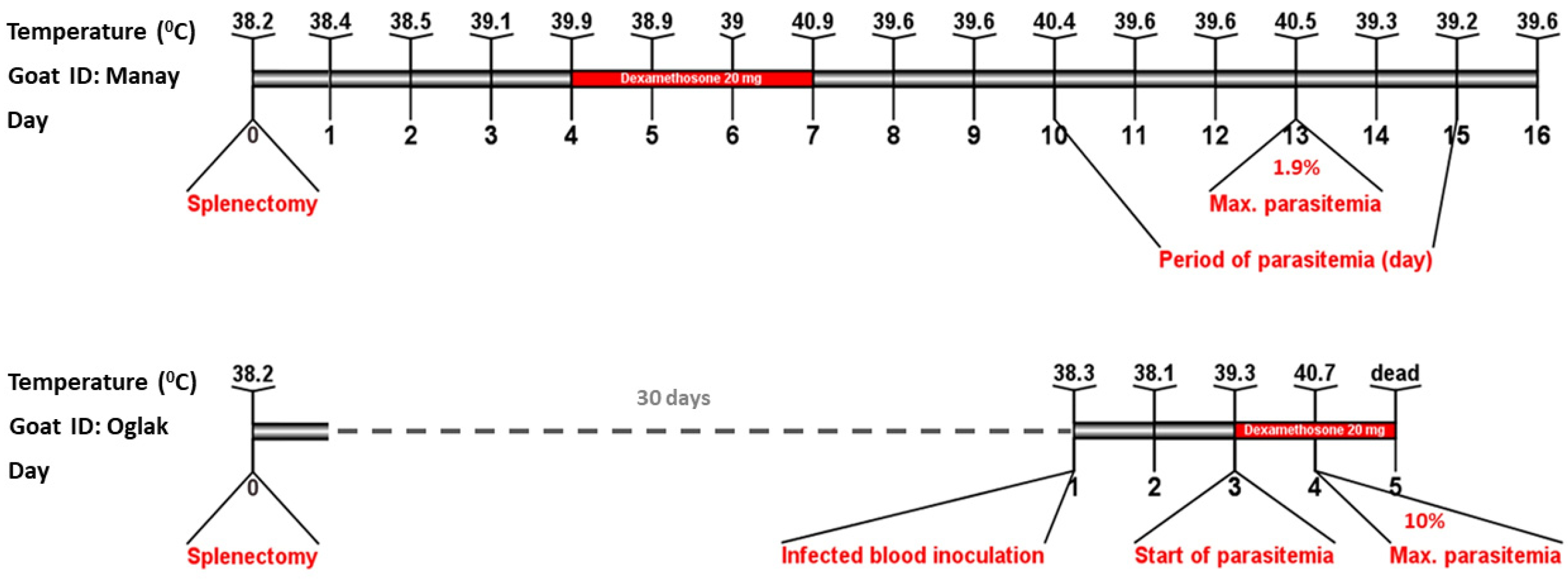
3.2. The Host’s Ability to Control Parasitemia Is Diminished by Splenectomy and Pharmacological Immunosuppression with Dexamethasone
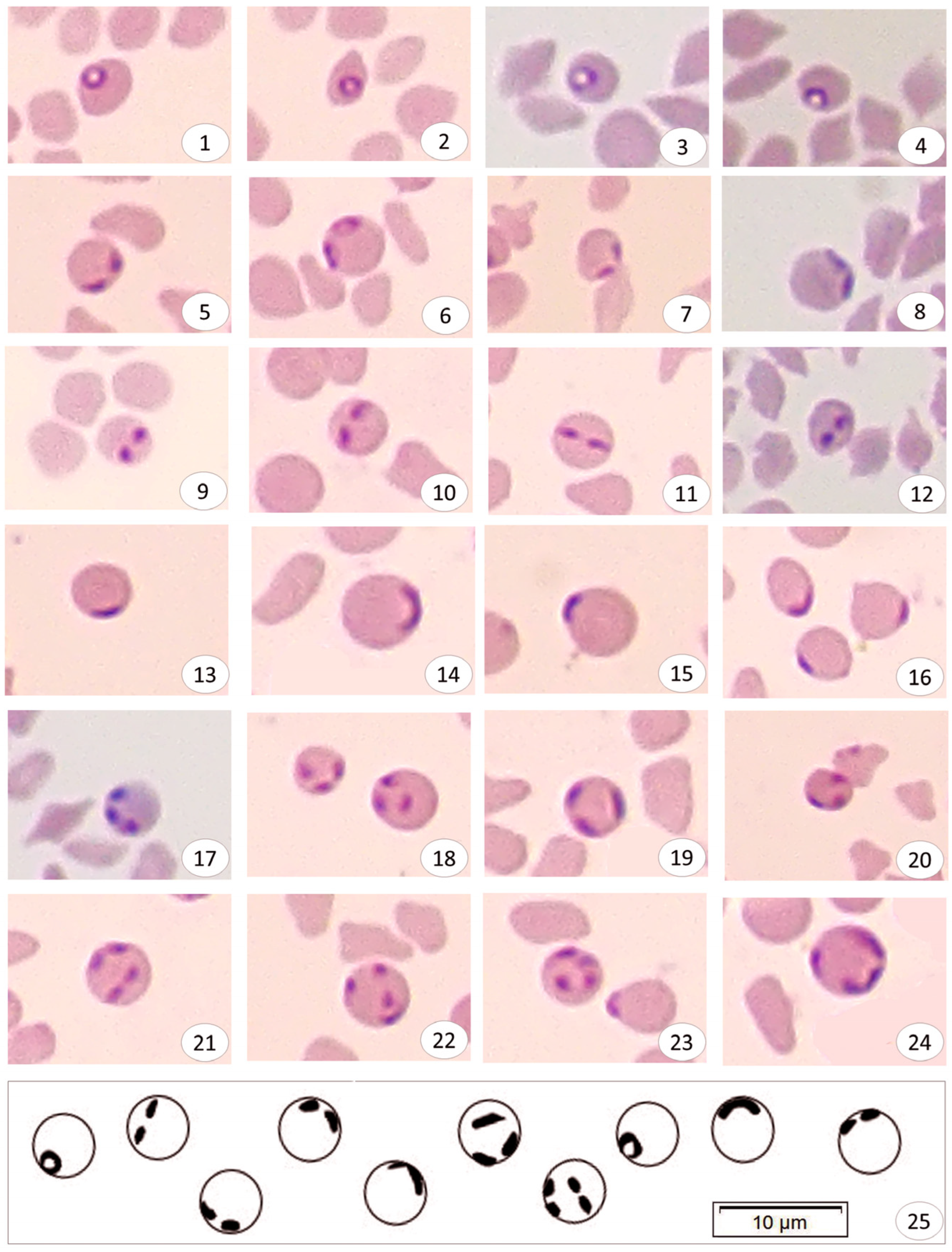
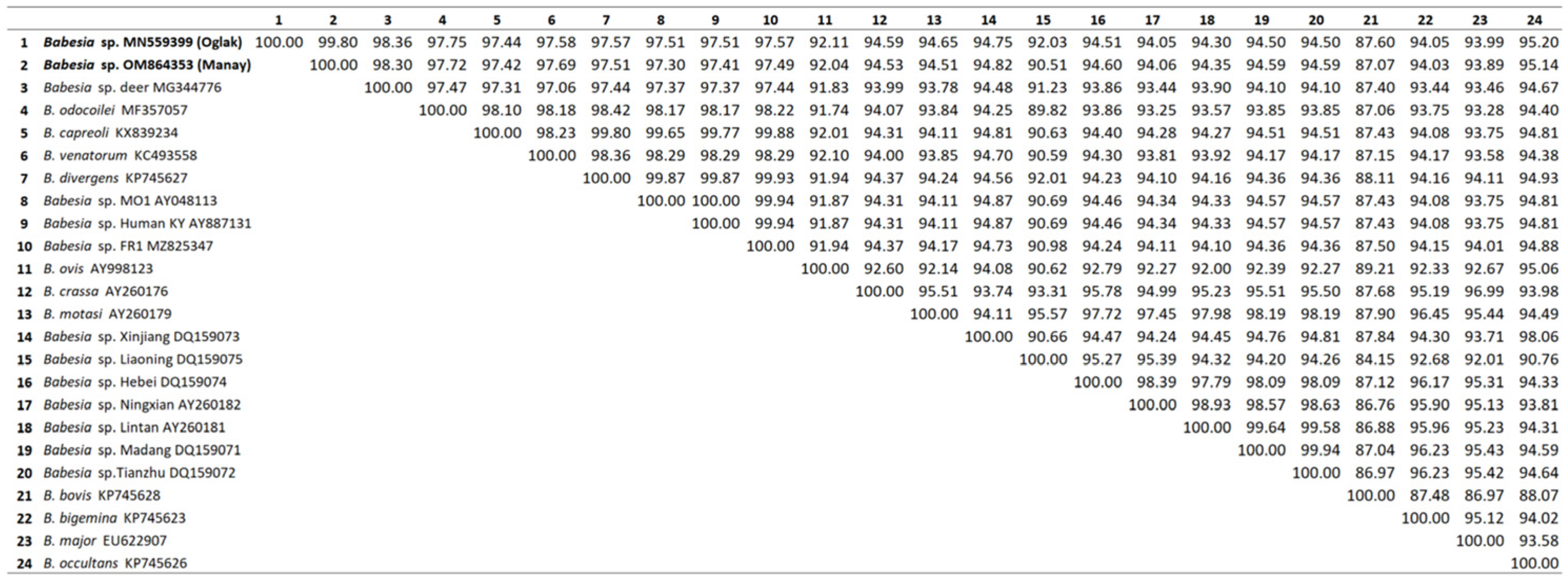
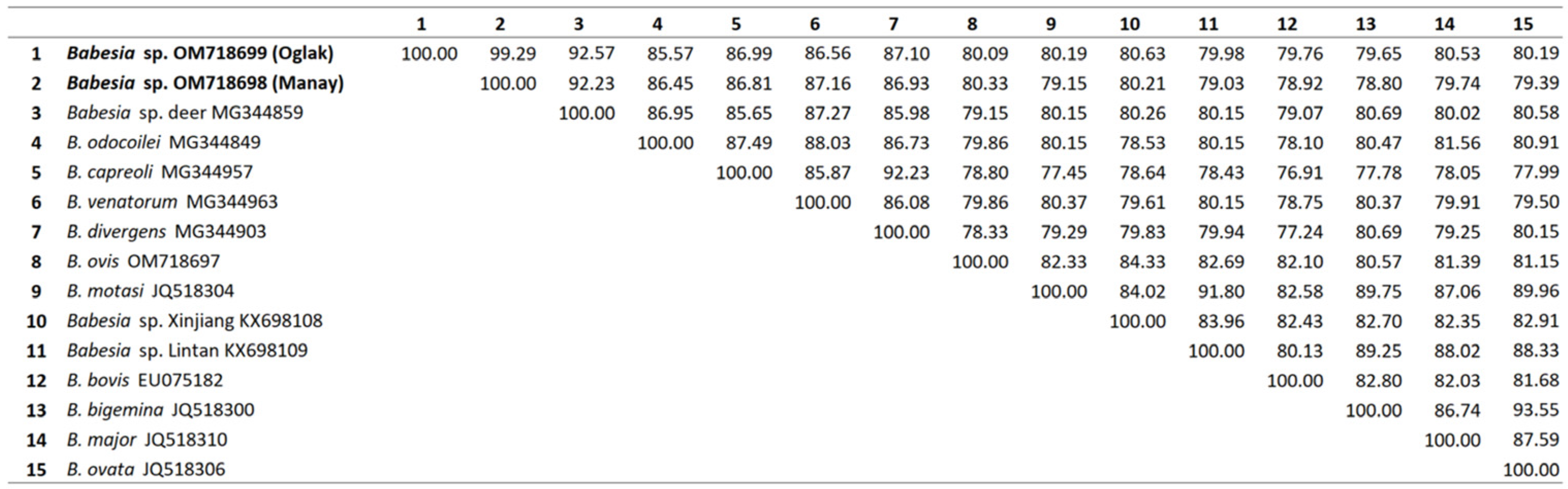
3.3. Sequence Comparisons and Phylogenetic Analysis
4. Discussion
5. Conclusions
6. Taxonomic Summary
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Babes, V. L’étiologie d’une enzootie des moutons, dénommée Carceag en Roumanie. C. R. Hebd. Acad. Sci. 1892, 115, 359–361. (In French) [Google Scholar]
- Uilenberg, G. Babesia—A historical overview. Vet. Parasitol. 2006, 138, 3–10. [Google Scholar] [CrossRef]
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef]
- Schnittger, L.; Ganzinelli, S.; Bhoora, R.; Omondi, D.; Nijhof, A.M.; Florin-Christensen, M. The Piroplasmida Babesia, Cytauxzoon, and Theileria in farm and companion animals: Species compilation, molecular phylogeny, and evolutionary insights. Parasitol. Res. 2022, 121, 1207–1245. [Google Scholar] [CrossRef]
- Yeruham, I.; Hadani, A.; Galker, F. Some epizootiological and clinical aspects of ovine babesiosis caused by Babesia ovis—A review. Vet. Parasitol. 1998, 74, 153–163. [Google Scholar] [CrossRef]
- Smith, M.; Sherman, D. Iodine deficiency. In Goat Medicine, 2nd ed.; Wiley-Blackwell: London, UK, 2009. [Google Scholar]
- Jia, N.; Zheng, Y.-C.; Jiang, J.-F.; Jiang, R.-R.; Jiang, B.-G.; Wei, R.; Liu, H.-B.; Huo, Q.-B.; Sun, Y.; Chu, Y.-L. Human babesiosis caused by a Babesia crassa–like pathogen: A case series. Clin. Infect. Dis. 2018, 67, 1110–1119. [Google Scholar] [CrossRef]
- Wang, J.; Gao, S.; Zhang, S.; He, X.; Liu, J.; Liu, A.; Li, Y.; Liu, G.; Luo, J.; Guan, G. Rapid detection of Babesia motasi responsible for human babesiosis by cross-priming amplification combined with a vertical flow. Parasites Vectors 2020, 13, 377. [Google Scholar] [CrossRef]
- Oosthuizen, M.C.; Zweygarth, E.; Collins, N.E.; Troskie, M.; Penzhorn, B.L. Identification of a novel Babesia sp. from a sable antelope (Hippotragus niger Harris, 1838). J. Clin. Microbiol. 2008, 46, 2247–2251. [Google Scholar] [CrossRef] [Green Version]
- Oosthuizen, M.C.; Allsopp, B.A.; Troskie, M.; Collins, N.E.; Penzhorn, B.L. Identification of novel Babesia and Theileria species in South African giraffe (Giraffa camelopardalis, Linnaeus, 1758) and roan antelope (Hippotragus equinus, Desmarest 1804). Vet. Parasitol. 2009, 163, 39–46. [Google Scholar] [CrossRef]
- Bajer, A.; Alsarraf, M.; Bednarska, M.; Mohallal, E.M.; Mierzejewska, E.J.; Behnke-Borowczyk, J.; Zalat, S.; Gilbert, F.; Welc-Falęciak, R. Babesia behnkei sp. nov., a novel Babesia species infecting isolated populations of Wagner’s gerbil, Dipodillus dasyurus, from the Sinai Mountains, Egypt. Parasites Vectors 2014, 7, 572. [Google Scholar] [CrossRef] [Green Version]
- Baneth, G.; Florin-Christensen, M.; Cardoso, L.; Schnittger, L. Reclassification of Theileria annae as Babesia vulpes sp. nov. Parasites Vectors 2015, 8, 207. [Google Scholar] [CrossRef]
- Baneth, G.; Nachum-Biala, Y.; Birkenheuer, A.J.; Schreeg, M.E.; Prince, H.; Florin-Christensen, M.; Schnittger, L.; Aroch, I. A new piroplasmid species infecting dogs: Morphological and molecular characterization and pathogeny of Babesia negevi n. sp. Parasites Vectors 2020, 13, 130. [Google Scholar] [CrossRef]
- Knowles, D.P.; Kappmeyer, L.S.; Haney, D.; Herndon, D.R.; Fry, L.M.; Munro, J.B.; Sears, K.; Ueti, M.W.; Wise, L.N.; Silva, M. Discovery of a novel species, Theileria haneyi n. sp., infective to equids, highlights exceptional genomic diversity within the genus Theileria: Implications for apicomplexan parasite surveillance. Int. J. Parasitol. 2018, 48, 679–690. [Google Scholar] [CrossRef]
- Nijhof, A.M.; Penzhorn, B.L.; Lynen, G.; Mollel, J.O.; Morkel, P.; Bekker, C.P.; Jongejan, F. Babesia bicornis sp. nov. and Theileria bicornis sp. nov.: Tick-borne parasites associated with mortality in the black rhinoceros (Diceros bicornis). J. Clin. Microbiol. 2003, 41, 2249–2254. [Google Scholar] [CrossRef] [Green Version]
- Ozubek, S.; Aktas, M. Molecular evidence of a new Babesia sp. in goats. Vet. Parasitol. 2017, 233, 1–8. [Google Scholar] [CrossRef]
- Turan, N. Türkiye’nin Yaban ve Av Hayvanları Memeliler; Ongun Kardeşler Matbaacılık Sanayii: Ankara, Turkiye, 1984. [Google Scholar]
- Kawahara, M.; Rikihisa, Y.; Lin, Q.; Isogai, E.; Tahara, K.; Itagaki, A.; Hiramitsu, Y.; Tajima, T. Novel genetic variants of Anaplasma phagocytophilum, Anaplasma bovis, Anaplasma centrale, and a novel Ehrlichia sp. in wild deer and ticks on two major islands in Japan. Appl. Environ. Microbiol. 2006, 72, 1102–1109. [Google Scholar] [CrossRef] [Green Version]
- Bekker, C.P.; De Vos, S.; Taoufik, A.; Sparagano, O.A.; Jongejan, F. Simultaneous detection of Anaplasma and Ehrlichia species in ruminants and detection of Ehrlichia ruminantium in Amblyomma variegatum ticks by reverse line blot hybridization. Vet. Microbiol. 2002, 89, 223–238. [Google Scholar] [CrossRef]
- Georges, K.; Loria, G.; Riili, S.; Greco, A.; Caracappa, S.; Jongejan, F.; Sparagano, O. Detection of haemoparasites in cattle by reverse line blot hybridisation with a note on the distribution of ticks in Sicily. Vet. Parasitol. 2001, 99, 273–286. [Google Scholar] [CrossRef]
- Pitulle, C.; Citron, D.M.; Bochner, B.; Barbers, R.; Appleman, M.D. Novel bacterium isolated from a lung transplant patient with cystic fibrosis. J. Clin. Microbiol. 1999, 37, 3851–3855. [Google Scholar] [CrossRef] [Green Version]
- Jensen, W.A.; Lappin, M.R.; Kamkar, S.; Reagan, W.J. Use of a polymerase chain reaction assay to detect and differentiate two strains of Haemobartonella felis in naturally infected cats. Am. J. Vet. Res. 2001, 62, 604–608. [Google Scholar] [CrossRef]
- Gou, H.; Guan, G.; Liu, A.; Ma, M.; Xu, Z.; Liu, Z.; Ren, Q.; Li, Y.; Yang, J.; Chen, Z. A DNA barcode for Piroplasmea. Acta Trop. 2012, 124, 92–97. [Google Scholar] [CrossRef]
- Bai, Q.; Liu, G.; Liu, D.; Ren, J.; Li, X. Isolation and preliminary characterization of a large Babesia sp. from sheep and goats in the eastern part of Gansu Province, China. Parasitol. Res. 2002, 88, S16–S21. [Google Scholar] [CrossRef]
- Sevinc, F.; Turgut, K.; Sevinc, M.; Ekici, O.D.; Coskun, A.; Koc, Y.; Erol, M.; Ica, A. Therapeutic and prophylactic efficacy of imidocarb dipropionate on experimental Babesia ovis infection of lambs. Vet. Parasitol. 2007, 149, 65–71. [Google Scholar] [CrossRef]
- Luo, J.; Chen, F.; Lu, W.; Guan, G.; Ma, M.; Yin, H. Experimental transmission of an unnamed bovine Babesia by Hyalomma spp., Haemaphysalis longicornis and Boophilus microplus. Vet. Parasitol. 2003, 116, 115–124. [Google Scholar] [CrossRef]
- Uilenberg, G.; Rombach, M.; Perié, N.; Zwart, D. Blood parasites of sheep in the Netherlands. II. Babesia motasi (Sporozoa, Babesiidae). Vet. Q. 1980, 2, 3–14. [Google Scholar]
- Guan, G.; Ma, M.; Moreau, E.; Liu, J.; Lu, B.; Bai, Q.; Luo, J.; Jorgensen, W.; Chauvin, A.; Yin, H. A new ovine Babesia species transmitted by Hyalomma anatolicum anatolicum. Exp. Parasitol. 2009, 122, 261–267. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Liu, W.; Xie, Y.; Ma, J.; Luo, X.; Nie, P.; Zuo, Z.; Lahrmann, U.; Zhao, Q.; Zheng, Y.; Zhao, Y. IBS: An illustrator for the presentation and visualization of biological sequences. Bioinformatics 2015, 31, 3359–3361. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Uilenberg, G. Encyclopedia of Arthropod-Transmitted Infections of Man and Domesticated Animals; CABI Publishing: New York, NY, USA, 2001. [Google Scholar]
- Ceylan, O.; Byamukama, B.; Ceylan, C.; Galon, E.M.; Liu, M.; Masatani, T.; Xuan, X.; Sevinc, F. Tick-borne hemoparasites of sheep: A molecular research in Turkey. Pathogens 2021, 10, 162. [Google Scholar] [CrossRef]
- Uilenberg, G.; Gray, J.; Kahl, O. Research on Piroplasmorida and other tick-borne agents: Are we going the right way? Ticks Tick-Borne Dis. 2018, 9, 860–863. [Google Scholar] [CrossRef]
- Mans, B.J. The basis of molecular diagnostics for piroplasmids: Do the sequences lie? Ticks Tick-Borne Dis. 2022, 13, 101907. [Google Scholar] [CrossRef] [PubMed]
- Sears, K.P.; Knowles, D.P.; Fry, L.M. Clinical Progression of Theileria haneyi in Splenectomized Horses Reveals Decreased Virulence Compared to Theileria equi. Pathogens 2022, 11, 254. [Google Scholar] [CrossRef] [PubMed]
- Buffet, P.A.; Safeukui, I.; Deplaine, G.; Brousse, V.; Prendki, V.; Thellier, M.; Turner, G.D.; Mercereau-Puijalon, O. The pathogenesis of Plasmodium falciparum malaria in humans: Insights from splenic physiology. Blood J. Am. Soc. Hematol. 2011, 117, 381–392. [Google Scholar]
- Sevinc, F.; Sevinc, M.; Koc, Y.; Alkan, F.; Ekici, O.D.; Yildiz, R.; Isik, N.; Aydogdu, U. The effect of 12 successive blood passages on the virulence of Babesia ovis in splenectomized lambs: A preliminary study. Small Rumin. Res. 2014, 116, 66–70. [Google Scholar] [CrossRef]
- Chauvin, A.; Valentin, A.; Malandrin, L.; L’Hostis, M. Sheep as a new experimental host for Babesia divergens. Vet. Res. 2002, 33, 429–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laha, R.; Das, M.; Sen, A. Morphology, epidemiology, and phylogeny of Babesia: An overview. Trop. Parasitol. 2015, 5, 94. [Google Scholar] [CrossRef] [Green Version]
- Shayan, P.; Hooshmand, E.; Nabian, S.; Rahbari, S. Biometrical and genetical characterization of large Babesia ovis in Iran. Parasitol. Res. 2008, 103, 217–221. [Google Scholar] [CrossRef]
- Hrazdilová, K.; Rybářová, M.; Široký, P.; Votýpka, J.; Zintl, A.; Burgess, H.; Steinbauer, V.; Žákovčík, V.; Modrý, D. Diversity of Babesia spp. in cervid ungulates based on the 18S rDNA and cytochrome c oxidase subunit I phylogenies. Infect. Genet. Evol. 2020, 77, 104060. [Google Scholar] [CrossRef]
- Andersson, M.O.; Bergvall, U.A.; Chirico, J.; Christensson, M.; Lindgren, P.-E.; Nordström, J.; Kjellander, P. Molecular detection of Babesia capreoli and Babesia venatorum in wild Swedish roe deer, Capreolus capreolus. Parasites Vectors 2016, 9, 221. [Google Scholar] [CrossRef] [Green Version]
- Pattullo, K.M.; Wobeser, G.; Lockerbie, B.P.; Burgess, H.J. Babesia odocoilei infection in a Saskatchewan elk (Cervus elaphus canadensis) herd. J. Vet. Diagn. Investig. 2013, 25, 535–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathieu, A.; Pastor, A.R.; Berkvens, C.N.; Gara-Boivin, C.; Hébert, M.; Léveillé, A.N.; Barta, J.R.; Smith, D.A. Babesia odocoilei as a cause of mortality in captive cervids in Canada. Can. Vet. J. 2018, 59, 52. [Google Scholar] [PubMed]
- Scott, J.D.; Pascoe, E.L.; Sajid, M.S.; Foley, J.E. Detection of Babesia odocoilei in Ixodes scapularis ticks collected in southern Ontario, Canada. Pathogens 2021, 10, 327. [Google Scholar] [CrossRef]
- Michel, A.O.; Mathis, A.; Ryser-Degiorgis, M.-P. Babesia spp. in European wild ruminant species: Parasite diversity and risk factors for infection. Vet. Res. 2014, 45, 65. [Google Scholar] [CrossRef] [PubMed]
- Häselbarth, K.; Tenter, A.M.; Brade, V.; Krieger, G.; Hunfeld, K.-P. First case of human babesiosis in Germany–clinical presentation and molecular characterisation of the pathogen. Int. J. Med. Microbiol. 2007, 297, 197–204. [Google Scholar] [CrossRef]
- Sun, Y.; Li, S.-G.; Jiang, J.-F.; Wang, X.; Zhang, Y.; Wang, H.; Cao, W.-C. Babesia venatorum infection in child, China. Emerg. Infect. Dis. 2014, 20, 896. [Google Scholar] [CrossRef]
- Gray, A.; Capewell, P.; Loney, C.; Katzer, F.; Shiels, B.R.; Weir, W. Sheep as host species for zoonotic Babesia venatorum, United Kingdom. Emerg. Infect. Dis. 2019, 25, 2257. [Google Scholar] [CrossRef] [Green Version]
- Overzier, E.; Pfister, K.; Herb, I.; Mahling, M.; Böck Jr, G.; Silaghi, C. Detection of tick-borne pathogens in roe deer (Capreolus capreolus), in questing ticks (Ixodes ricinus), and in ticks infesting roe deer in southern Germany. Ticks Tick-Borne Dis. 2013, 4, 320–328. [Google Scholar] [CrossRef]
- Zintl, A.; Mulcahy, G.; Skerrett, H.E.; Taylor, S.M.; Gray, J.S. Babesia divergens, a bovine blood parasite of veterinary and zoonotic importance. Clin. Microbiol. Rev. 2003, 16, 622–636. [Google Scholar] [CrossRef] [Green Version]
- Wiegmann, L.; Silaghi, C.; Obiegala, A.; Karnath, C.; Langer, S.; Ternes, K.; Kämmerling, J.; Osmann, C.; Pfeffer, M. Occurrence of Babesia species in captive reindeer (Rangifer tarandus) in Germany. Vet. Parasitol. 2015, 211, 16–22. [Google Scholar] [CrossRef]
- Criado-Fornelio, A.; Martinez-Marcos, A.; Buling-Sarana, A.; Barba-Carretero, J. Molecular studies on Babesia, Theileria and Hepatozoon in southern Europe: Part II. Phylogenetic analysis and evolutionary history. Vet. Parasitol. 2003, 114, 173–194. [Google Scholar] [CrossRef] [PubMed]
- Bonsergent, C.; de Carné, M.-C.; de la Cotte, N.; Moussel, F.; Perronne, V.; Malandrin, L. The new human Babesia sp. FR1 Is a European member of the Babesia sp. MO1 Clade. Pathogens 2021, 10, 1433. [Google Scholar] [CrossRef]
- Azagi, T.; Jaarsma, R.I.; Docters van Leeuwen, A.; Fonville, M.; Maas, M.; Franssen, F.F.; Kik, M.; Rijks, J.M.; Montizaan, M.G.; Groenevelt, M. Circulation of Babesia species and their exposure to humans through Ixodes ricinus. Pathogens 2021, 10, 386. [Google Scholar] [CrossRef] [PubMed]
- Gubbels, J.; De Vos, A.; Van der Weide, M.; Viseras, J.; Schouls, L.; De Vries, E.; Jongejan, F. Simultaneous detection of bovine Theileria and Babesia species by reverse line blot hybridization. J. Clin. Microbiol. 1999, 37, 1782–1789. [Google Scholar] [CrossRef]
- Nagore, D.; Garcıa-Sanmartın, J.; Garcıa-Pérez, A.L.; Juste, R.A.; Hurtado, A. Identification, genetic diversity and prevalence of Theileria and Babesia species in a sheep population from Northern Spain. Int. J. Parasitol. 2004, 34, 1059–1067. [Google Scholar] [CrossRef]
- Schnittger, L.; Yin, H.; Qi, B.; Gubbels, M.J.; Beyer, D.; Niemann, S.; Jongejan, F.; Ahmed, J.S. Simultaneous detection and differentiation of Theileria and Babesia parasites infecting small ruminants by reverse line blotting. Parasitol. Res. 2004, 92, 189–196. [Google Scholar] [CrossRef]
- Altay, K.; Dumanli, N.; Aktas, M. Molecular identification, genetic diversity and distribution of Theileria and Babesia species infecting small ruminants. Vet. Parasitol. 2007, 147, 161–165. [Google Scholar] [CrossRef]
- Adamu, M.; Troskie, M.; Oshadu, D.O.; Malatji, D.P.; Penzhorn, B.L.; Matjila, P.T. Occurrence of tick-transmitted pathogens in dogs in Jos, Plateau State, Nigeria. Parasites Vectors 2014, 7, 119. [Google Scholar] [CrossRef] [Green Version]
- Schouls, L.M.; Van De Pol, I.; Rijpkema, S.G.; Schot, C.S. Detection and identification of Ehrlichia, Borrelia burgdorferi sensu lato, and Bartonella species in Dutch Ixodes ricinus ticks. J. Clin. Microbiol. 1999, 37, 2215–2222. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. Positive | Identified Pathogens | ||||
|---|---|---|---|---|---|
| Babesia sp. | B. ovis | T. ovis | A. ovis | Mycoplasma spp. | |
| 2 | + | − | − | − | − |
| 1 | − | − | + | − | − |
| 3 | + | + | − | − | − |
| 4 | + | − | + | − | + |
| 3 | + | − | + | + | + |
| 1 | − | + | + | + | + |
| 14 (28%) | 12 (24%) | 4 (8%) | 9 (18%) | 4 (8%) | 8 (16%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozubek, S.; Ulucesme, M.C.; Aktas, M. Discovery of a Novel Species Infecting Goats: Morphological and Molecular Characterization of Babesia aktasi n. sp. Pathogens 2023, 12, 113. https://doi.org/10.3390/pathogens12010113
Ozubek S, Ulucesme MC, Aktas M. Discovery of a Novel Species Infecting Goats: Morphological and Molecular Characterization of Babesia aktasi n. sp. Pathogens. 2023; 12(1):113. https://doi.org/10.3390/pathogens12010113
Chicago/Turabian StyleOzubek, Sezayi, Mehmet Can Ulucesme, and Munir Aktas. 2023. "Discovery of a Novel Species Infecting Goats: Morphological and Molecular Characterization of Babesia aktasi n. sp." Pathogens 12, no. 1: 113. https://doi.org/10.3390/pathogens12010113
APA StyleOzubek, S., Ulucesme, M. C., & Aktas, M. (2023). Discovery of a Novel Species Infecting Goats: Morphological and Molecular Characterization of Babesia aktasi n. sp. Pathogens, 12(1), 113. https://doi.org/10.3390/pathogens12010113