
Correlation between the Biodistribution of Bovine Leukemia Virus in the Organs and the Proviral Load in the Peripheral Blood during Early Stages of Experimentally Infected Cattle

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Collection of Blood and Organs
2.3. DNA Extraction from the Blood and Organs
2.4. Detection and Quantification of the Bovine Leukemia Virus Proviral Load
2.5. Detection of the Anti-Bovine Leukemia Virus Antibodies in the Serum Samples
2.6. Statistical Analysis
3. Results
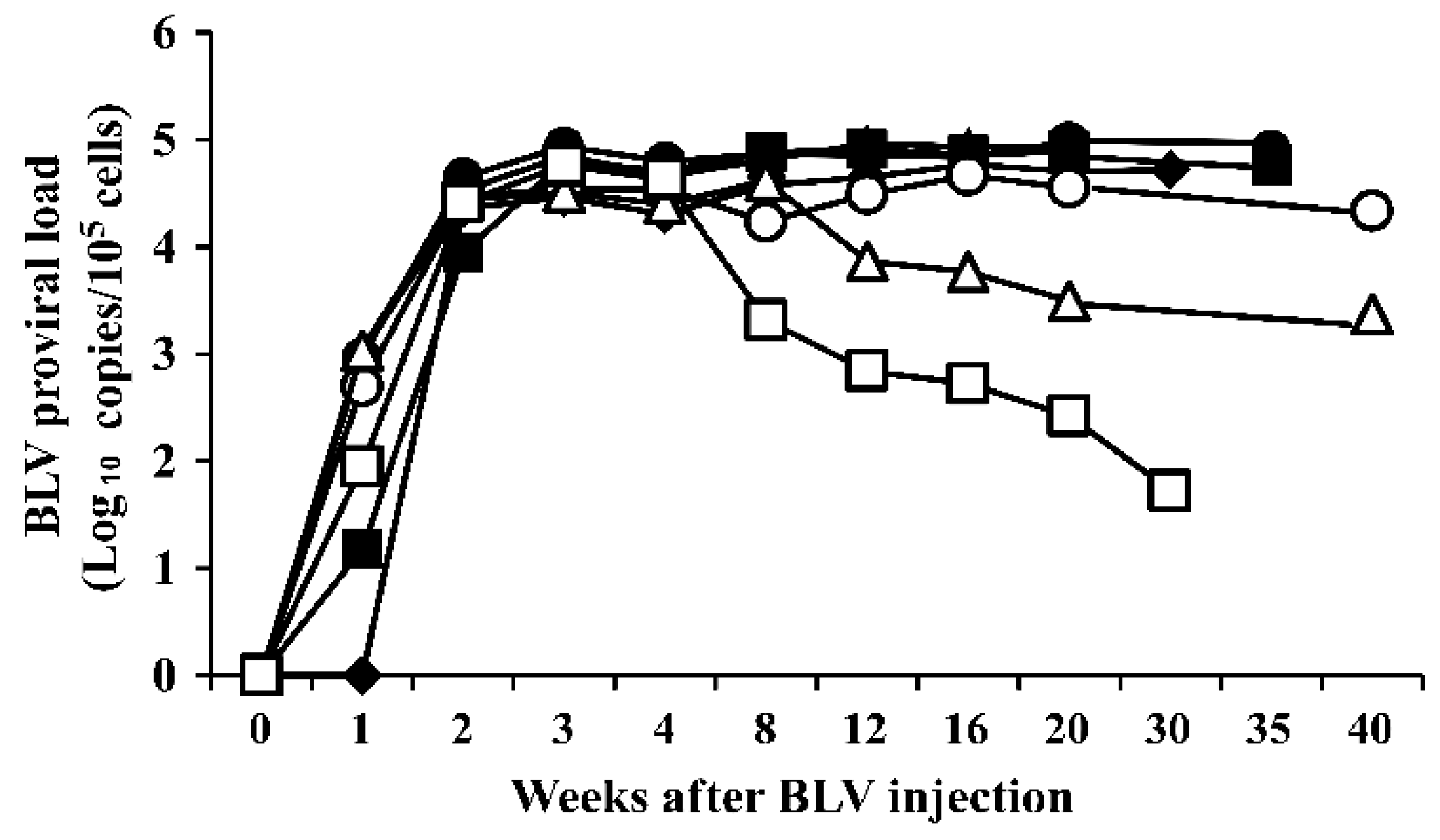
3.1. Bovine Leukemia Virus Proviral Load in the Blood of Bovine Leukemia Virus-Infected Cattle
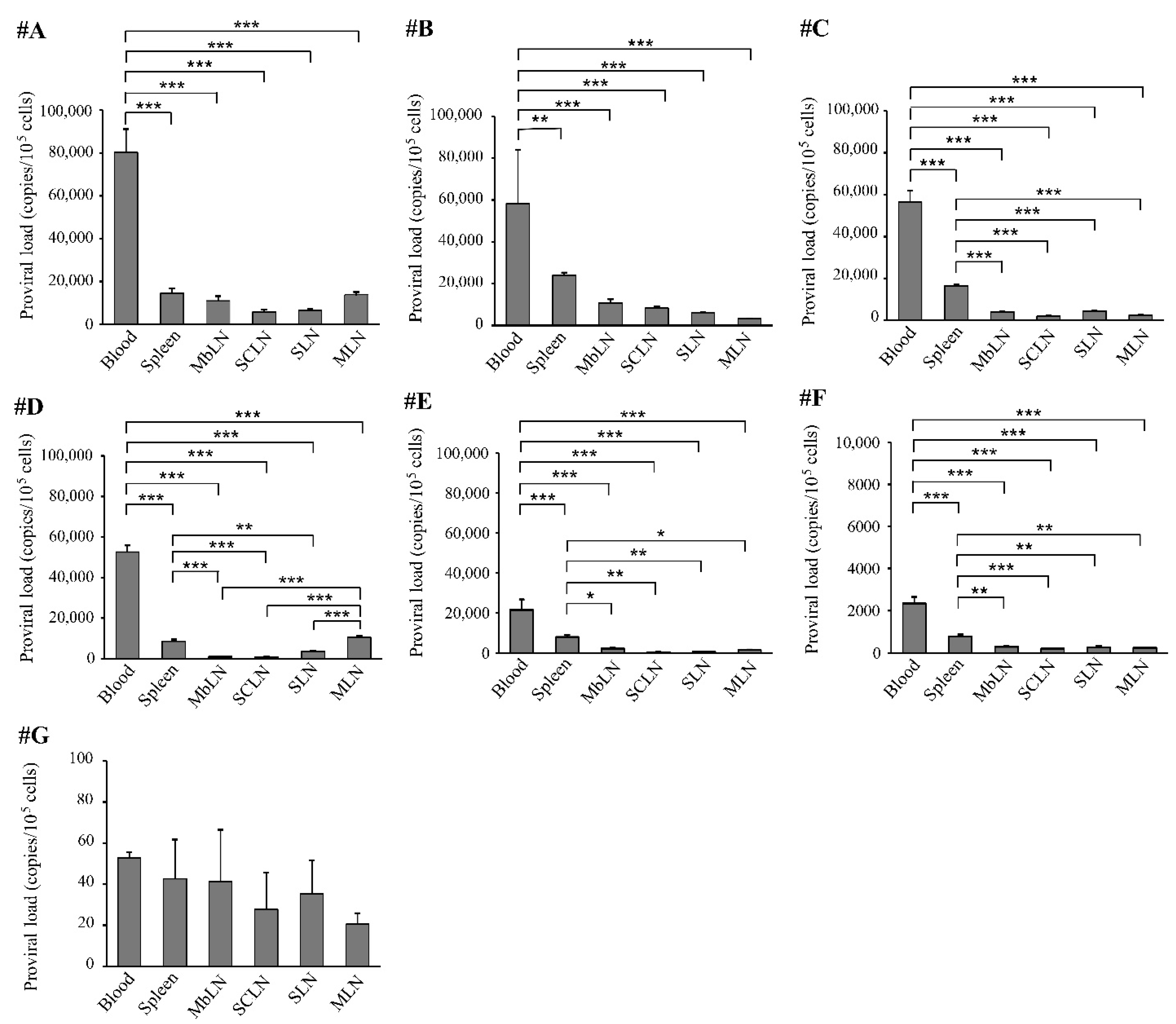
3.2. Distribution of Bovine Leukemia Virus Proviral DNA in the Organs
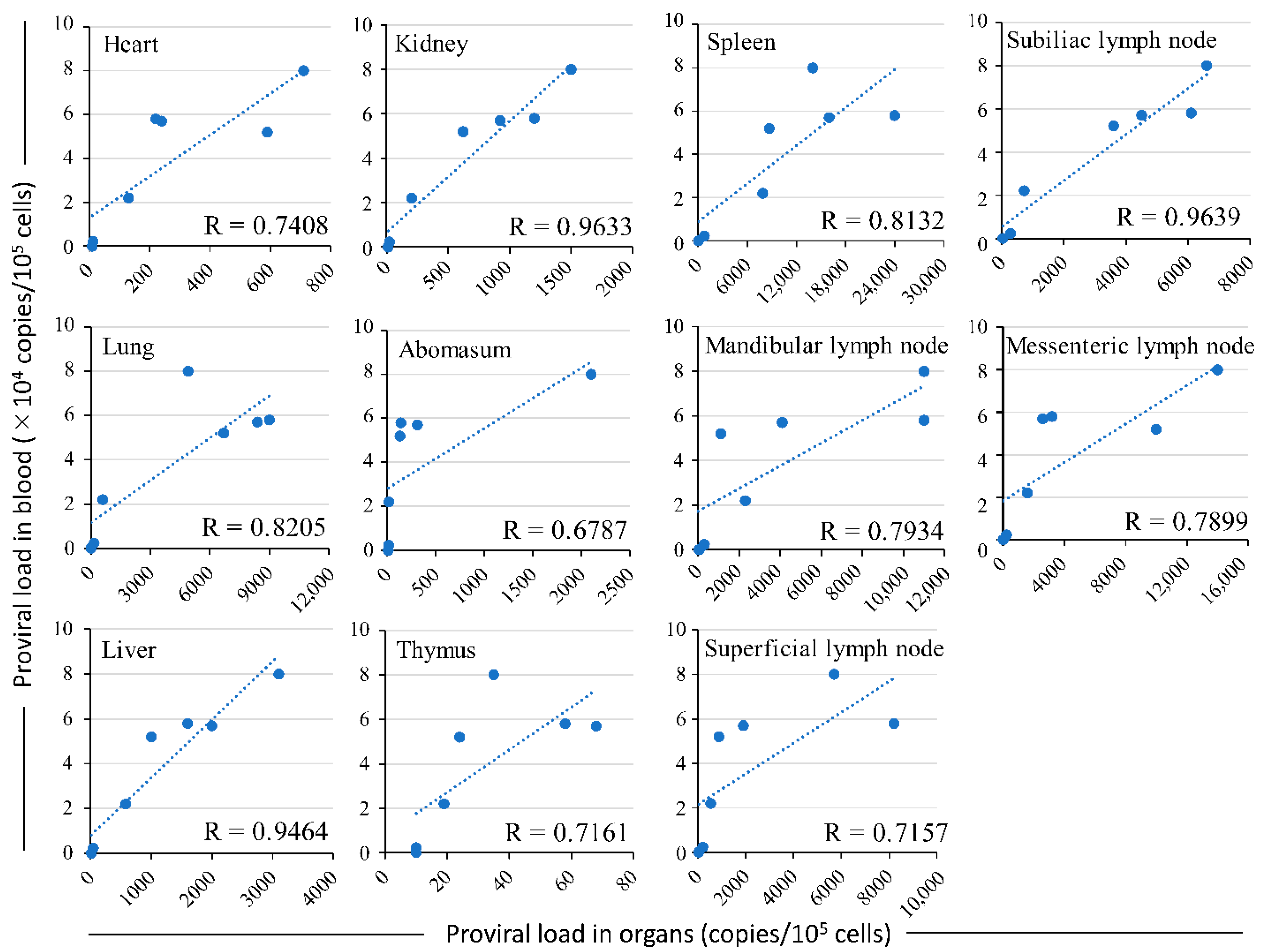
3.3. Comparison of the Distribution of Bovine Leukemia Virus Proviral DNA in the Organs and the Proviral Load in the Blood
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aida, Y.; Murakami, H.; Takahashi, M.; Takeshima, S.N. Mechanisms of pathogenesis induced by bovine leukemia virus as a model for human T-cell leukemia virus. Front. Microbiol. 2013, 4, 328. [Google Scholar] [CrossRef] [Green Version]
- Kettmann, R.; Deschamps, J.; Cleuter, Y.; Couez, D.; Burny, A.; Marbaix, G. Leukemogenesis by bovine leukemia virus: Proviral DNA integration and lack of RNA expression of viral long terminal repeat and 3′ proximate cellular sequences. Proc. Natl. Acad. Sci. USA 1982, 79, 2465–2469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tajima, S.; Ikawa, Y.; Aida, Y. Complete bovine leukemia virus (BLV) provirus is conserved in BLV-infected cattle throughout the course of B-cell lymphosarcoma development. J. Virol. 1998, 72, 7569–7576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillet, N.; Florins, A.; Boxus, M.; Burteau, C.; Nigro, A.; Vandermeers, F.; Balon, H.; Bouzar, A.B.; Defoiche, J.; Burny, A.; et al. Mechanisms of leukemogenesis induced by bovine leukemia virus: Prospects for novel anti-retroviral therapies in human. Retrovirology 2007, 4, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimba, M.; Takeshima, S.N.; Murakami, H.; Kohara, J.; Kobayashi, N.; Matsuhashi, T.; Ohmori, T.; Nunoya, T.; Aida, Y. BLV-CoCoMo-qPCR: A useful tool for evaluating bovine leukemia virus infection status. BMC Vet. Res. 2012, 8, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimba, M.; Takeshima, S.N.; Matoba, K.; Endoh, D.; Aida, Y. BLV-CoCoMo-qPCR: Quantitation of bovine leukemia virus proviral load using the CoCoMo algorithm. Retrovirology 2010, 7, 91. [Google Scholar] [CrossRef] [Green Version]
- Somura, Y.; Sugiyama, E.; Fujikawa, H.; Murakami, K. Comparison of the copy numbers of bovine leukemia virus in the lymph nodes of cattle with enzootic bovine leukosis and cattle with latent infection. Arch. Virol. 2014, 159, 2693–2697. [Google Scholar] [CrossRef]
- Kobayashi, T.; Inagaki, Y.; Ohnuki, N.; Sato, R.; Murakami, S.; Imakawa, K. Increasing Bovine leukemia virus (BLV) proviral load is a risk factor for progression of Enzootic bovine leucosis: A prospective study in Japan. Prev. Vet. Med. 2020, 178, 104680. [Google Scholar] [CrossRef]
- Sato, H.; Watanuki, S.; Bai, L.; Borjigin, L.; Ishizaki, H.; Matsumoto, Y.; Hachiya, Y.; Sentsui, H.; Aida, Y. A sensitive luminescence syncytium induction assay (LuSIA) based on a reporter plasmid containing a mutation in the glucocorticoid response element in the long terminal repeat U3 region of bovine leukemia virus. Virol. J. 2019, 16, 66. [Google Scholar] [CrossRef] [Green Version]
- Sato, H.; Watanuki, S.; Murakami, H.; Sato, R.; Ishizaki, H.; Aida, Y. Development of a luminescence syncytium induction assay (LuSIA) for easily detecting and quantitatively measuring bovine leukemia virus infection. Arch. Virol. 2018, 163, 1519–1530. [Google Scholar] [CrossRef]
- Bai, L.; Borjigin, L.; Sato, H.; Takeshima, S.N.; Asaji, S.; Ishizaki, H.; Kawashima, K.; Obuchi, Y.; Sunaga, S.; Ando, A.; et al. Kinetic Study of BLV Infectivity in BLV Susceptible and Resistant Cattle in Japan from 2017 to 2019. Pathogens 2021, 10, 1281. [Google Scholar] [CrossRef] [PubMed]
- Ohno, A.; Takeshima, S.N.; Matsumoto, Y.; Aida, Y. Risk factors associated with increased bovine leukemia virus proviral load in infected cattle in Japan from 2012 to 2014. Virus. Res. 2015, 210, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Panei, C.J.; Takeshima, S.N.; Omori, T.; Nunoya, T.; Davis, W.C.; Ishizaki, H.; Matoba, K.; Aida, Y. Estimation of bovine leukemia virus (BLV) proviral load harbored by lymphocyte subpopulations in BLV-infected cattle at the subclinical stage of enzootic bovine leucosis using BLV-CoCoMo-qPCR. BMC Vet. Res. 2013, 9, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Kitamura-Muramatsu, Y.; Saito, S.; Ishizaki, H.; Nakano, M.; Haga, S.; Matoba, K.; Ohno, A.; Murakami, H.; Takeshima, S.N.; et al. Detection of the BLV provirus from nasal secretion and saliva samples using BLV-CoCoMo-qPCR-2: Comparison with blood samples from the same cattle. Virus Res. 2015, 210, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Watanuki, S.; Takeshima, S.N.; Borjigin, L.; Sato, H.; Bai, L.; Murakami, H.; Sato, R.; Ishizaki, H.; Matsumoto, Y.; Aida, Y. Visualizing bovine leukemia virus (BLV)-infected cells and measuring BLV proviral loads in the milk of BLV seropositive dams. Vet. Res. 2019, 50, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakatsuchi, A.; Watanuki, S.; Borjigin, L.; Sato, H.; Bai, L.; Matsuura, R.; Kuroda, M.; Murakami, H.; Sato, R.; Asaji, S.; et al. BoLA-DRB3 Polymorphism Controls Proviral Load and Infectivity of Bovine Leukemia Virus (BLV) in Milk. Pathogens 2022, 11, 210. [Google Scholar] [CrossRef]
- Takeshima, S.N.; Ohno, A.; Aida, Y. Bovine leukemia virus proviral load is more strongly associated with bovine major histocompatibility complex class II DRB3 polymorphism than with DQA1 polymorphism in Holstein cow in Japan. Retrovirology 2019, 16, 14. [Google Scholar] [CrossRef]
- Lo, C.W.; Borjigin, L.; Saito, S.; Fukunaga, K.; Saitou, E.; Okazaki, K.; Mizutani, T.; Wada, S.; Takeshima, S.N.; Aida, Y. BoLA-DRB3 Polymorphism is Associated with Differential Susceptibility to Bovine Leukemia Virus-Induced Lymphoma and Proviral Load. Viruses 2020, 12, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juliarena, M.A.; Barrios, C.N.; Ceriani, M.C.; Esteban, E.N. Hot topic: Bovine leukemia virus (BLV)-infected cows with low proviral load are not a source of infection for BLV-free cattle. J. Dairy Sci. 2016, 99, 4586–4589. [Google Scholar] [CrossRef] [Green Version]
- Lagarias, D.M.; Radke, K. Transient increases of blood mononuclear cells that could express bovine leukemia virus early after experimental infection of sheep. Microb. Pathog. 1990, 9, 147–158. [Google Scholar] [CrossRef]
- Radke, K.; Grossman, D.; Kidd, L.C. Humoral immune response of experimentally infected sheep defines two early periods of bovine leukemia virus replication. Microb. Pathog. 1990, 9, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Heeney, J.L.; de Vries, P.; Dubbes, R.; Koornstra, W.; Niphuis, H.; ten Haaft, P.; Boes, J.; Dings, M.E.; Morein, B.; Osterhaus, A.D. Comparison of protection from homologous cell-free vs cell-associated SIV challenge afforded by inactivated whole SIV vaccines. J. Med. Primatol. 1992, 21, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Domenech, A.; Goyache, J.; Llames, L.; Jesus Paya, M.; Suarez, G.; Gomez-Lucia, E. In vitro infection of cells of the monocytic/macrophage lineage with bovine leukaemia virus. J. Gen. Virol. 2000, 81, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Forletti, A.; Lutzelschwab, C.M.; Cepeda, R.; Esteban, E.N.; Gutierrez, S.E. Early events following bovine leukaemia virus infection in calves with different alleles of the major histocompatibility complex DRB3 gene. Vet. Res. 2020, 51, 4. [Google Scholar] [CrossRef] [Green Version]
- Klintevall, K.; Ballagi-Pordany, A.; Naslund, K.; Belak, S. Bovine leukaemia virus: Rapid detection of proviral DNA by nested PCR in blood and organs of experimentally infected calves. Vet. Microbiol. 1994, 42, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Takeshima, S.N.; Kitamura-Muramatsu, Y.; Yuan, Y.; Polat, M.; Saito, S.; Aida, Y. BLV-CoCoMo-qPCR-2: Improvements to the BLV-CoCoMo-qPCR assay for bovine leukemia virus by reducing primer degeneracy and constructing an optimal standard curve. Arch. Virol. 2015, 160, 1325–1332. [Google Scholar] [CrossRef]
- Tajima, S.; Takahashi, M.; Takeshima, S.N.; Konnai, S.; Yin, S.A.; Watarai, S.; Tanaka, Y.; Onuma, M.; Okada, K.; Aida, Y. A mutant form of the tax protein of bovine leukemia virus (BLV), with enhanced transactivation activity, increases expression and propagation of BLV in vitro but not in vivo. J. Virol. 2003, 77, 1894–1903. [Google Scholar] [CrossRef] [Green Version]
- Miyasaka, T.; Takeshima, S.N.; Jimba, M.; Matsumoto, Y.; Kobayashi, N.; Matsuhashi, T.; Sentsui, H.; Aida, Y. Identification of bovine leukocyte antigen class II haplotypes associated with variations in bovine leukemia virus proviral load in Japanese Black cattle. Tissue Antigens 2013, 81, 72–82. [Google Scholar] [CrossRef]
- Polat, M.; Ohno, A.; Takeshima, S.N.; Kim, J.; Kikuya, M.; Matsumoto, Y.; Mingala, C.N.; Onuma, M.; Aida, Y. Detection and molecular characterization of bovine leukemia virus in Philippine cattle. Arch. Virol. 2015, 160, 285–296. [Google Scholar] [CrossRef]
- Mammerickx, M.; Otte, J.; Rase, F.; Braibant, E.; Portetelle, D.; Burny, A.; Dekegel, D.; Ghysdael, J. Large scale serological detection in Belgium of enzootic bovine leukosis. Zent. Vet. B 1978, 25, 416–424. [Google Scholar] [CrossRef]
- Mebius, R.E.; Kraal, G. Structure and function of the spleen. Nat. Rev. Immunol. 2005, 5, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Van Der Maaten, M.J.; Miller, J.M. Sites of in vivo replication of bovine leukemia virus in experimentally infected cattle. Ann. Rech. Vet. 1978, 9, 831–835. [Google Scholar] [PubMed]
- Chakrabarti, A.C. Permeability of membranes to amino acids and modified amino acids: Mechanisms involved in translocation. Amino Acids 1994, 6, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Lafeuillade, A.; Poggi, C.; Sayada, C.; Pellegrino, P.; Profizi, N. Focusing on the second phase of plasma HIV-1 RNA clearance. AIDS 1997, 11, 264–266. [Google Scholar] [PubMed]
- Mirsky, M.L.; Olmstead, C.; Da, Y.; Lewin, H.A. Reduced bovine leukaemia virus proviral load in genetically resistant cattle. Anim. Genet. 1998, 29, 245–252. [Google Scholar] [CrossRef]
- Miyasaka, T.; Takeshima, S.N.; Sentsui, H.; Aida, Y. Identification and diversity of bovine major histocompatibility complex class II haplotypes in Japanese Black and Holstein cattle in Japan. J. Dairy Sci. 2012, 95, 420–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juliarena, M.A.; Poli, M.; Sala, L.; Ceriani, C.; Gutierrez, S.; Dolcini, G.; Rodriguez, E.M.; Marino, B.; Rodriguez-Dubra, C.; Esteban, E.N. Association of BLV infection profiles with alleles of the BoLA-DRB3.2 gene. Anim. Genet. 2008, 39, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.W.; Takeshima, S.N.; Okada, K.; Saitou, E.; Fujita, T.; Matsumoto, Y.; Wada, S.; Inoko, H.; Aida, Y. Association of Bovine Leukemia Virus-Induced Lymphoma with BoLA-DRB3 Polymorphisms at DNA, Amino Acid, and Binding Pocket Property Levels. Pathogens 2021, 10, 437. [Google Scholar] [CrossRef]
- Takeshima, S.N.; Sasaki, S.; Meripet, P.; Sugimoto, Y.; Aida, Y. Single nucleotide polymorphisms in the bovine MHC region of Japanese Black cattle are associated with bovine leukemia virus proviral load. Retrovirology 2017, 14, 24. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cattle a | Breed | Age b (Month) Start End | Sex | BLV Proviral Load in Blood (Copies/105 Cells) c | Number of PBL (/μL) d | Antibodies to BLV e |
|---|---|---|---|---|---|---|
| ||||||
| #A | Japanese Black | 4–12 | ♂ | 8.0 × 104 | 9.2 × 103 | + |
| #B | Japanese Black | 5–14 | ♂ | 5.8 × 104 | 8.8 × 103 | + |
| #C | Japanese Black | 5–13 | ♂ | 5.7 × 104 | 8.7 × 103 | + |
| #D | Japanese Black | 5–12 | ♀ | 5.2 × 104 | 8.6 × 103 | + |
| #E | Holstein-Friesian | 12–22 | ♂ | 2.2 × 104 | 7.1 × 103 | + |
| ||||||
| #F | Holstein-Friesian | 13–23 | ♂ | 2.3 × 103 | 5.2 × 103 | + |
| ||||||
| #G | Japanese Black | 5–12 | ♀ | 5.3 × 10 | 4.9 × 103 | + |
| Organs and Peripheral Blood | Experimental Infected Cattle | |||||||
|---|---|---|---|---|---|---|---|---|
| #A | #B | #C | #D | #E | #F | #G | Average | |
| Right auricle of the heart | 7.1 × 102 | 2.2 × 102 | 2.4 × 102 | 5.9 × 102 | 1.3 × 102 | 1.3 × 10 | <10 | 2.7 × 102 |
| Lung | 4.9 × 103 | 9.0 × 103 | 8.4 × 103 | 6.7 × 103 | 6.0 × 102 | 1.6 × 102 | <10 | 4.2 × 103 |
| Liver | 3.1 × 103 | 1.6 × 103 | 2.0 × 103 | 1.0 × 103 | 5.8 × 102 | 4.44 × 10 | <10 | 1.2 × 103 |
| Kidney | 1.5 × 103 | 1.2 × 103 | 9.2 × 102 | 6.2 × 102 | 2.0 × 102 | 2.0 × 10 | <10 | 6.3 × 102 |
| Abomasum | 2.1 × 103 | 1.4 × 102 | 3.1 × 102 | 1.3 × 102 | 1.7 × 10 | 1.4 × 10 | <10 | 3.8 × 102 |
| Thymus | 3.5 × 10 | 5.8 × 10 | 6.8 × 10 | 2.4 × 10 | 1.9 × 10 | <10 | <10 | 3.0 × 10 |
| Spleen | 1.4 × 104 | 2.4 × 104 | 1.6 × 104 | 8.7 × 103 | 7.9 × 103 | 7.7 × 102 | 4.3 × 10 | 1.0 × 104 |
| Mandibular lymph node | 1.1 × 104 | 1.1 × 104 | 4.1 × 103 | 1.1 × 103 | 2.3 × 103 | 3.0 × 102 | 4.1 × 10 | 4.2 × 103 |
| Superficial cervical lymph node | 5.7 × 103 | 8.2 × 103 | 1.9 × 103 | 8.7 × 102 | 5.3 × 102 | 2.0 × 102 | 2.8 × 10 | 2.5 × 103 |
| Subiliac lymph node | 6.6 × 103 | 6.1 × 103 | 4.5 × 103 | 3.6 × 103 | 7.2 × 102 | 2.8 × 102 | 3.6 × 10 | 3.1 × 103 |
| Mesenteric lymph node | 1.4 × 104 | 3.2 × 103 | 2.6 × 103 | 1.0 × 104 | 1.6 × 103 | 2.4 × 102 | 2.0 × 10 | 4.5 × 103 |
| Peripheral Blood | 8.0 × 104 | 5.8 × 104 | 5.7 × 104 | 5.2 × 104 | 2.2 × 104 | 2.3 × 103 | 5.3 × 10 | 3.9 × 104 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kohara, J.; Bai, L.; Takeshima, S.-n.; Matsumoto, Y.; Hirai, T.; Aida, Y. Correlation between the Biodistribution of Bovine Leukemia Virus in the Organs and the Proviral Load in the Peripheral Blood during Early Stages of Experimentally Infected Cattle. Pathogens 2023, 12, 130. https://doi.org/10.3390/pathogens12010130
Kohara J, Bai L, Takeshima S-n, Matsumoto Y, Hirai T, Aida Y. Correlation between the Biodistribution of Bovine Leukemia Virus in the Organs and the Proviral Load in the Peripheral Blood during Early Stages of Experimentally Infected Cattle. Pathogens. 2023; 12(1):130. https://doi.org/10.3390/pathogens12010130
Chicago/Turabian StyleKohara, Junko, Lanlan Bai, Shin-nosuke Takeshima, Yuki Matsumoto, Tsunao Hirai, and Yoko Aida. 2023. "Correlation between the Biodistribution of Bovine Leukemia Virus in the Organs and the Proviral Load in the Peripheral Blood during Early Stages of Experimentally Infected Cattle" Pathogens 12, no. 1: 130. https://doi.org/10.3390/pathogens12010130
APA StyleKohara, J., Bai, L., Takeshima, S. -n., Matsumoto, Y., Hirai, T., & Aida, Y. (2023). Correlation between the Biodistribution of Bovine Leukemia Virus in the Organs and the Proviral Load in the Peripheral Blood during Early Stages of Experimentally Infected Cattle. Pathogens, 12(1), 130. https://doi.org/10.3390/pathogens12010130