

The Mechanisms Involved in the Fluoroquinolone Resistance of Salmonella enterica Strains Isolated from Humans in Poland, 2018–2019: The Prediction of Antimicrobial Genes by In Silico Whole-Genome Sequencing
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Salmonella enterica Isolates and Fluoroquinolone Susceptibility Testing
2.2. Whole-Genome Sequencing Analysis
3. Results
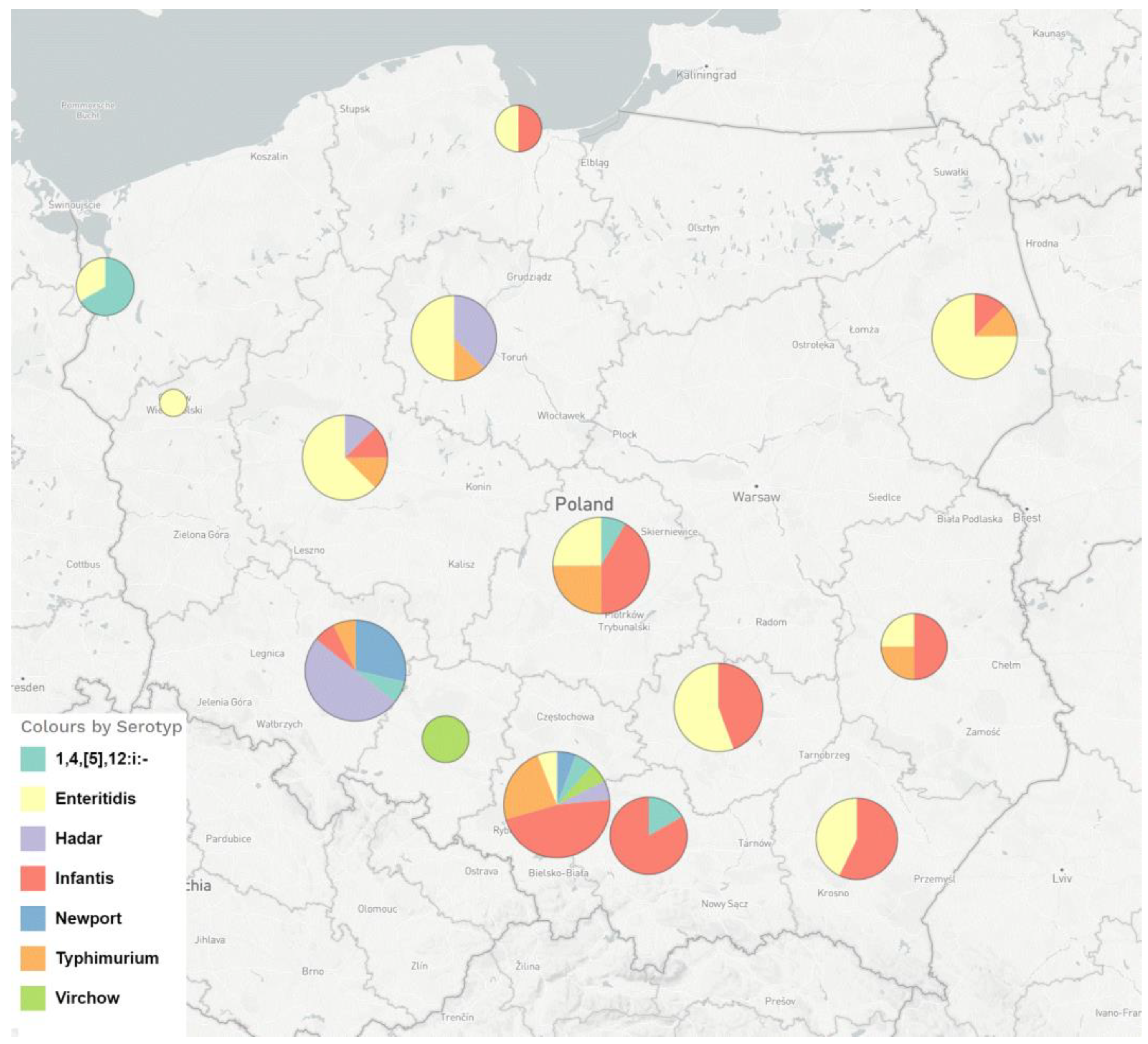
3.1. Salmonella Strains and Phenotypic Resistance to Fluoroquinolones
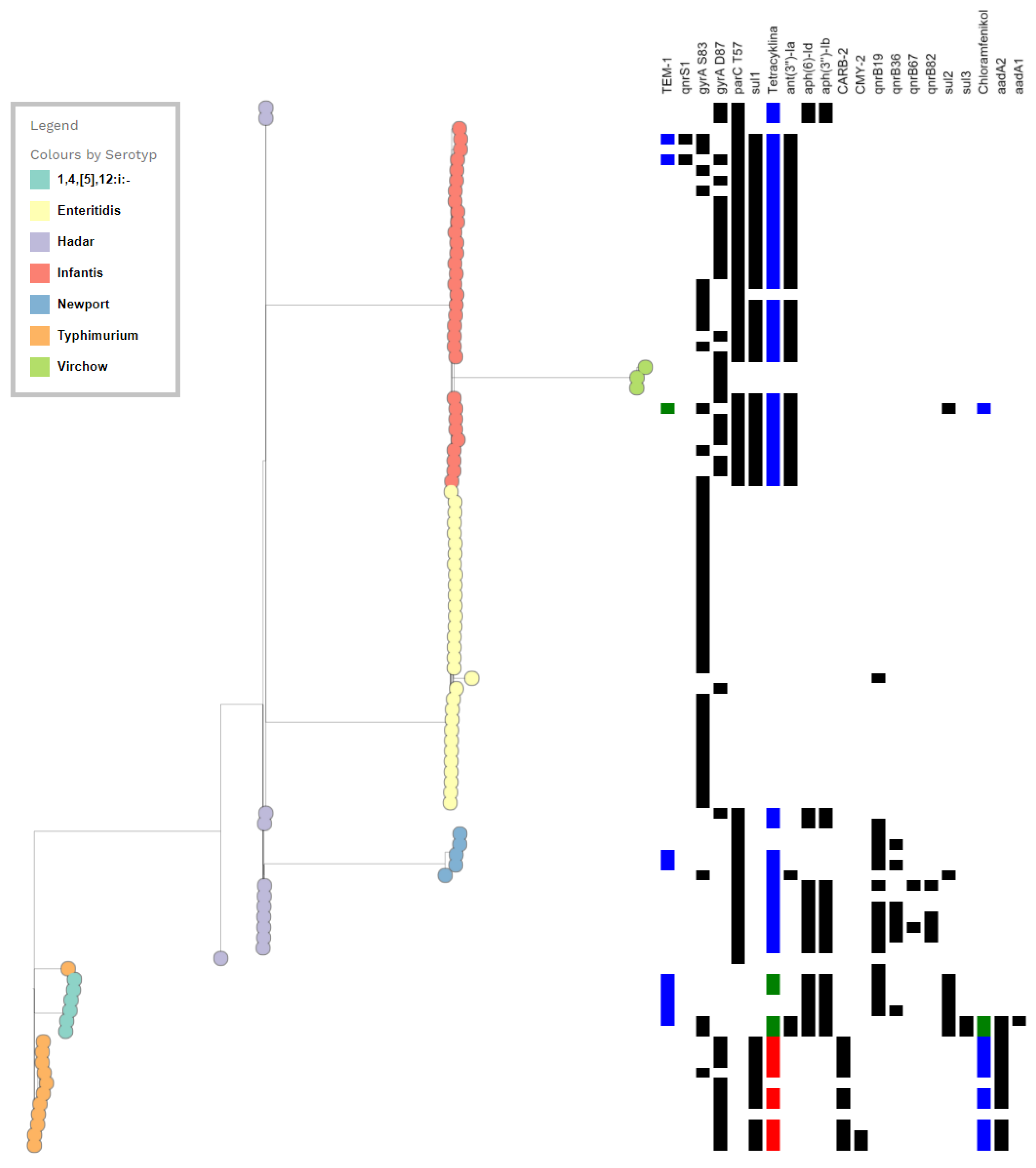
3.2. Genotypic Antimicrobial Analysis In Silico
3.2.1. Resistance to Fluoroquinolones–QRDR Mutations and PMQR Genes
3.2.2. In Silico Antimicrobial Resistance Genes Detected among FQ-Resistant Salmonella Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grimont, P.A.D.; Weill, F.X. Antigenic Formulae of the Salmonella Serovars, 9th ed.; WHO Collaborating Centre for Reference and Research on Salmonella, Pasteur Institute: Paris, France, 2007. [Google Scholar]
- Authority, E.F.S. European Centre for Disease Prevention and Control the European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2018/2019. EFSA J. 2021, 19, e06490. [Google Scholar] [CrossRef]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.-A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne Illness Acquired in the United States—Major Pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.; Hopkins, K.L.; Meunier, D.; de Pinna, E.; Fitzgerald-Hughes, D.; Humphreys, H.; Woodford, N. Resistance to Third-Generation Cephalosporins in Human Non-Typhoidal Salmonella enterica Isolates from England and Wales, 2010–12. J. Antimicrob. Chemother. 2014, 69, 977–981. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Critically Important Antimicrobials for Human Medicine, 6th ed.; World Health Organization: Geneva, Switzerland, 2019; ISBN 978-92-4-151552-8. [Google Scholar]
- Medicine, C.; For, V. Extralabel Use and Antimicrobials; FDA: Silver Spring, MD, USA, 2022.
- Van, T.T.H.; Nguyen, H.N.K.; Smooker, P.M.; Coloe, P.J. The Antibiotic Resistance Characteristics of Non-Typhoidal Salmonella Enterica Isolated from Food-Producing Animals, Retail Meat and Humans in South East Asia. Int. J. Food Microbiol. 2012, 154, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Yanat, B.; Rodríguez-Martínez, J.-M.; Touati, A. Plasmid-Mediated Quinolone Resistance in Enterobacteriaceae: A Systematic Review with a Focus on Mediterranean Countries. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2017, 36, 421–435. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Z.; Xu, X.; Gu, Z.; Meng, J.; Wufuer, X.; Wang, M.; Huang, M.; Chen, J.; Jing, C.; Xiong, Z.; et al. Molecular Epidemiology and Antimicrobial Resistance of Invasive Non-Typhoidal Salmonella in China, 2007–2016. Infect. Drug Resist. 2019, 12, 2885–2897. [Google Scholar] [CrossRef] [Green Version]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Hooper, D.C. Mechanisms of Fluoroquinolone Resistance. Drug Resist. Updat. Rev. Comment. Antimicrob. Anticancer Chemother. 1999, 2, 38–55. [Google Scholar] [CrossRef]
- Piekarska, K.; Wołkowicz, T.; Zacharczuk, K.; Rzeczkowska, M.; Chróst, A.; Bareja, E.; Olak, M.; Gierczyński, R. Co-Existence of Plasmid-Mediated Quinolone Resistance Determinants and Mutations in GyrA and ParC among Fluoroquinolone-Resistant Clinical Enterobacteriaceae Isolated in a Tertiary Hospital in Warsaw, Poland. Int. J. Antimicrob. Agents 2015, 45, 238–243. [Google Scholar] [CrossRef]
- Jacoby, G.A.; Strahilevitz, J.; Hooper, D.C. Plasmid-Mediated Quinolone Resistance. Microbiol. Spectr. 2014, 2, 475–503. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.-B.; Lauderdale, T.-L.Y.; Huang, C.-H.; Chang, P.-R.; Wang, Y.-H.; Shigemura, K.; Lin, Y.-H.; Chang, W.-C.; Wang, K.-C.; Huang, T.-W.; et al. Genotypic Diversity of Ciprofloxacin Nonsusceptibility and Its Relationship with Minimum Inhibitory Concentrations in Nontyphoidal Salmonella Clinical Isolates in Taiwan. Antibiotics 2021, 10, 1383. [Google Scholar] [CrossRef] [PubMed]
- Salmonellosis-Annual Epidemiological Report for 2017. Available online: https://www.ecdc.europa.eu/en/publications-data/salmonellosis-annual-epidemiological-report-2017 (accessed on 24 October 2022).
- Wołkowicz, T.; Zacharczuk, K.; Gierczyński, R.; Nowakowska, M.; Piekarska, K. Antimicrobial Resistance and Whole-Genome Characterisation of High-Level Ciprofloxacin-Resistant Salmonella Enterica Serovar Kentucky ST 198 Strains Isolated from Human in Poland. Int. J. Mol. Sci. 2021, 22, 9381. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yin, Y.; Jones, M.B.; Zhang, Z.; Deatherage Kaiser, B.L.; Dinsmore, B.A.; Fitzgerald, C.; Fields, P.I.; Deng, X. Salmonella Serotype Determination Utilizing High-Throughput Genome Sequencing Data. J. Clin. Microbiol. 2015, 53, 1685–1692. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Official Journal of the European Union. Commission Implementing Decision (EU) 2018/945 of 22 June 2018 on the Communicable Diseases and Related Special Health Issues to Be Covered by Epidemiological Surveillance as Well as Relevant Case Definitions (Text with EEA Relevance); Official Journal of the European Union: Luxembourg, 2018; Volume 170. [Google Scholar]
- Apreja, M.; Sharma, A.; Balda, S.; Kataria, K.; Capalash, N.; Sharma, P. Antibiotic Residues in Environment: Antimicrobial Resistance Development, Ecological Risks, and Bioremediation. Environ. Sci. Pollut. Res. 2022, 29, 3355–3371. [Google Scholar] [CrossRef]
- Pikkemaat, M.G.; Yassin, H.; Fels-Klerx, H.J.; Berendsen, B.J.A. RIKILT-BU Toxicology Bioassays & Novel Foods, VLAG, RIKILT-Business unit Dierbehandelingsmiddelen. Antibiotic Residues and Resistance in the Environment; Pikkemaat, M.G., Yassin, H., Fels-Klerx, H.J., Berendsen, B.J.A., Eds.; RIKILT Wageningen UR: Wageningen, The Netherlands, 2016. [Google Scholar]
- Campos, M.J.; Palomo, G.; Hormeño, L.; Herrera-León, S.; Domínguez, L.; Vadillo, S.; Píriz, S.; Quesada, A. Prevalence of Quinolone Resistance Determinants in Non-Typhoidal Salmonella Isolates from Human Origin in Extremadura, Spain. Diagn. Microbiol. Infect. Dis. 2014, 79, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Gunell, M.; Webber, M.A.; Kotilainen, P.; Lilly, A.J.; Caddick, J.M.; Jalava, J.; Huovinen, P.; Siitonen, A.; Hakanen, A.J.; Piddock, L.J.V. Mechanisms of Resistance in Nontyphoidal Salmonella Enterica Strains Exhibiting a Nonclassical Quinolone Resistance Phenotype. Antimicrob. Agents Chemother. 2009, 53, 3832–3836. [Google Scholar] [CrossRef] [Green Version]
- Chang, M.-X.; Zhang, J.-F.; Sun, Y.-H.; Li, R.-S.; Lin, X.-L.; Yang, L.; Webber, M.A.; Jiang, H.-X. Contribution of Different Mechanisms to Ciprofloxacin Resistance in Salmonella Spp. Front. Microbiol. 2021, 12, 663731. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, J.M.; Machuca, J.; Cano, M.E.; Calvo, J.; Martínez-Martínez, L.; Pascual, A. Plasmid-Mediated Quinolone Resistance: Two Decades On. Drug Resist. Updat. 2016, 29, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Acheampong, G.; Owusu, M.; Owusu-Ofori, A.; Osei, I.; Sarpong, N.; Sylverken, A.; Kung, H.-J.; Cho, S.-T.; Kuo, C.-H.; Park, S.E.; et al. Chromosomal and Plasmid-Mediated Fluoroquinolone Resistance in Human Salmonella Enterica Infection in Ghana. BMC Infect. Dis. 2019, 19, 898. [Google Scholar] [CrossRef] [Green Version]
- Karp, B.E.; Campbell, D.; Chen, J.C.; Folster, J.P.; Friedman, C.R. Plasmid-Mediated Quinolone Resistance in Human Nontyphoidal Salmonella Infections: An Emerging Public Health Problem in the United States. Zoonoses Public Health 2018, 65, 838–849. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Lee, S.-K.; Park, M.-S.; Na, H.-T. Analysis of the Fluoroquinolone Antibiotic Resistance Mechanism of Salmonella Enterica Isolates. J. Microbiol. Biotechnol. 2016, 26, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Park, N.; Yun, S.; Hur, E.; Song, J.; Lee, H.; Kim, Y.; Ryu, S. Presence of Plasmid-Mediated Quinolone Resistance (PMQR) Genes in Non-Typhoidal Salmonella Strains with Reduced Susceptibility to Fluoroquinolones Isolated from Human Salmonellosis in Gyeonggi-Do, South Korea from 2016 to 2019. Gut Pathog. 2021, 13, 35. [Google Scholar] [CrossRef] [PubMed]
- Garneau-Tsodikova, S.; Labby, K.J. Mechanisms of Resistance to Aminoglycoside Antibiotics: Overview and Perspectives. MedChemComm 2016, 7, 11–27. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, G.L.; Panzenhagen, P.; Ferrari, R.G.; dos Santos, A.; Paschoalin, V.M.F.; Conte-Junior, C.A. Frequency of Antimicrobial Resistance Genes in Salmonella from Brazil by in Silico Whole-Genome Sequencing Analysis: An Overview of the Last Four Decades. Front. Microbiol. 2020, 11, 1864. [Google Scholar] [CrossRef] [PubMed]
- Cox, G.W.; Parmley, E.J.; Avery, B.P.; Irwin, R.J.; Reid-Smith, R.J.; Deckert, A.E.; Finley, R.L.; Daignault, D.; Alexander, D.C.; Allen, V.; et al. A One-Health Genomic Investigation of Gentamicin Resistance in Salmonella from Human and Chicken Sources in Canada, 2014 to 2017. Antimicrob. Agents Chemother. 2021, 65, e00966. [Google Scholar] [CrossRef] [PubMed]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global Trends in Antimicrobial Use in Food Animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harb, A.; Habib, I.; Mezal, E.H.; Kareem, H.S.; Laird, T.; O’Dea, M.; Abraham, S. Occurrence, Antimicrobial Resistance and Whole-Genome Sequencing Analysis of Salmonella Isolates from Chicken Carcasses Imported into Iraq from Four Different Countries. Int. J. Food Microbiol. 2018, 284, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Randall, L.P.; Cooles, S.W.; Osborn, M.K.; Piddock, L.J.V.; Woodward, M.J. Antibiotic Resistance Genes, Integrons and Multiple Antibiotic Resistance in Thirty-Five Serotypes of Salmonella Enterica Isolated from Humans and Animals in the UK. J. Antimicrob. Chemother. 2004, 53, 208–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llanes, C.; Kirchgesner, V.; Plesiat, P. Propagation of TEM- and PSE-Type β-Lactamases among Amoxicillin-Resistant Salmonella Spp. Isolated in France. Antimicrob. Agents Chemother. 1999, 43, 2430–2436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, A.C.; Turnidge, J.; Collignon, P.; Looke, D.; Barton, M.; Gottlieb, T. Control of Fluoroquinolone Resistance through Successful Regulation, Australia. Emerg. Infect. Dis. 2012, 18, 1453–1460. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Serotype | No of Isolates Tested | No of Ciprofloxacin Resistant Isolates (%) | Ciprofloxacin MIC Value (mg/L) |
|---|---|---|---|
| 1,4,[5],12:i:- | 37 | 6 (16.2%) | 0.38–0.5 |
| Hadar | 13 | 12 (92.3%) | 0.125–0.25 |
| Infantis | 58 | 32 (55.2%) | 0.19–3 |
| Newport | 7 | 5 (71.4%) | 0.19–0.75 |
| Typhimurium | 84 | 12 (14.3%) | 0.125–0.38 |
| Virchow | 4 | 3 (75%) | 0.125–0.25 |
| Enteritidis | 222 | 98 (44.1%) | 0.125–0.75 |
| Antimicrobial Class | AMR Gene | Isolates Positive for Each Gene (n) | ||
|---|---|---|---|---|
| Fluoroquinolones | QRDR mutations | GyrA | Ser83→Tyr | 45 |
| Asp87→Asn Asp87→Tyr | 14 21 | |||
| ParC | Thr57→Ser | 47 | ||
| PMQRs | qnrB19 | 17 | ||
| qnrB36 | 7 | |||
| qnrB67 | 2 | |||
| qnrB82 | 4 | |||
| qnrS1 | 2 | |||
| β-lactams | blaTEM-1A | 2 | ||
| blaTEM-1B | 9 | |||
| blaTEM-1C | 1 | |||
| blaCARB-2 | 8 | |||
| blaCMY-2 | 2 | |||
| Tetracyclines | tet(A) | 43 | ||
| tet(B) | 4 | |||
| tet(G) | 9 | |||
| Sulphonamides | sul1 | 39 | ||
| sul2 | 8 | |||
| sul3 | 2 | |||
| Aminoglycosides | aac(6′)-Iaa | 68 | ||
| ant(3″)-Ia | 29 | |||
| aph(3″)-Ib | 19 | |||
| aph(6)-Id | 18 | |||
| aadA1 | 1 | |||
| aadA2 | 12 | |||
| Phenicol | floR | 10 | ||
| cmlA1 | 2 | |||
| Serotype (Isolates taken into the WGS) | GyrA | No Isolates | ParC | No Isolates | PMQR | |||
|---|---|---|---|---|---|---|---|---|
| qnrB | No Isolates | qnrS | No Isolates | |||||
| Typhimurium (n = 12) | Ser83→Tyr | 1 | - | - | qnrB 19 | 1 | - | - |
| Asp87→Asn | 10 | |||||||
| 1,4,[5],12:i:- (n = 6) | Ser83→Tyr | 2 | - | - | qnrB 19 qnrB 36 | 4 1 | - | - |
| Hadar (n = 12) | Asp87→Asn | 2 | Thr57→Ser | 12 | qnrB 19 qnrB 36 qnrB 67 qnrB 82 | 7 4 2 4 | - | - |
| Asp87→Tyr | 1 | |||||||
| Infantis (n = 30) | Ser83→Tyr | 12 | Thr57→Ser | 30 | - | - | qnrS 1 | 2 |
| Asp87→Tyr | 18 | |||||||
| Newport (n = 5) | Ser83→Tyr | 1 | Thr57→Ser | 5 | qnrB 19 qnrB 36 | 4 2 | - | - |
| Virchow (n = 3) | Asp87→Asn | 2 | - | - | - | - | - | - |
| Asp87→Tyr | 1 | |||||||
| Enteritidis (n = 32) | Ser83→Tyr | 29 | - | - | qnrB 19 | 1 | - | - |
| Asp87→Tyr | 1 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piekarska, K.; Wołkowicz, T.; Zacharczuk, K.; Stepuch, A.; Gierczyński, R. The Mechanisms Involved in the Fluoroquinolone Resistance of Salmonella enterica Strains Isolated from Humans in Poland, 2018–2019: The Prediction of Antimicrobial Genes by In Silico Whole-Genome Sequencing. Pathogens 2023, 12, 193. https://doi.org/10.3390/pathogens12020193
Piekarska K, Wołkowicz T, Zacharczuk K, Stepuch A, Gierczyński R. The Mechanisms Involved in the Fluoroquinolone Resistance of Salmonella enterica Strains Isolated from Humans in Poland, 2018–2019: The Prediction of Antimicrobial Genes by In Silico Whole-Genome Sequencing. Pathogens. 2023; 12(2):193. https://doi.org/10.3390/pathogens12020193
Chicago/Turabian StylePiekarska, Katarzyna, Tomasz Wołkowicz, Katarzyna Zacharczuk, Agata Stepuch, and Rafał Gierczyński. 2023. "The Mechanisms Involved in the Fluoroquinolone Resistance of Salmonella enterica Strains Isolated from Humans in Poland, 2018–2019: The Prediction of Antimicrobial Genes by In Silico Whole-Genome Sequencing" Pathogens 12, no. 2: 193. https://doi.org/10.3390/pathogens12020193
APA StylePiekarska, K., Wołkowicz, T., Zacharczuk, K., Stepuch, A., & Gierczyński, R. (2023). The Mechanisms Involved in the Fluoroquinolone Resistance of Salmonella enterica Strains Isolated from Humans in Poland, 2018–2019: The Prediction of Antimicrobial Genes by In Silico Whole-Genome Sequencing. Pathogens, 12(2), 193. https://doi.org/10.3390/pathogens12020193