
Epidemiology of Ebolaviruses from an Etiological Perspective


Abstract
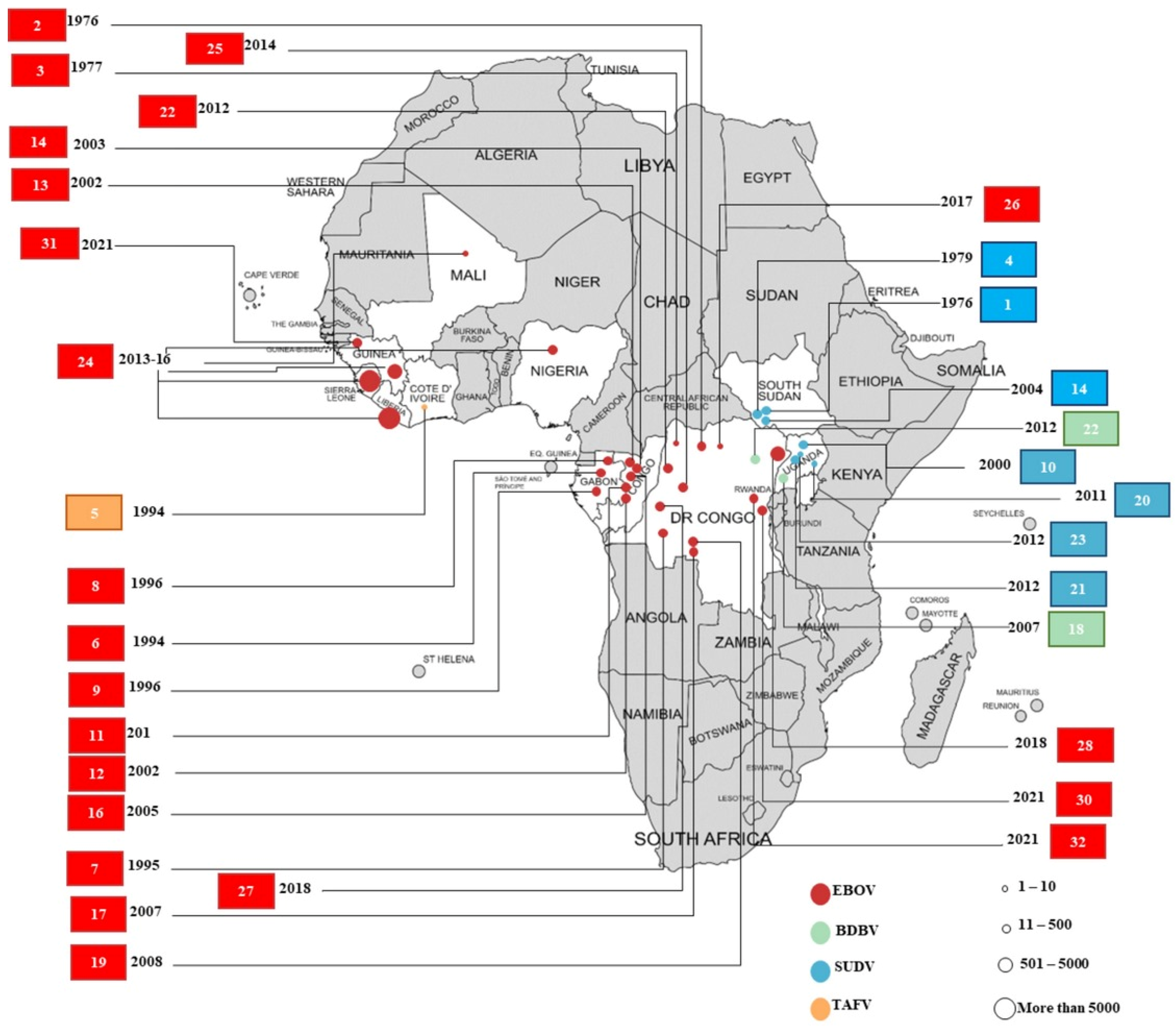
:1. Introduction
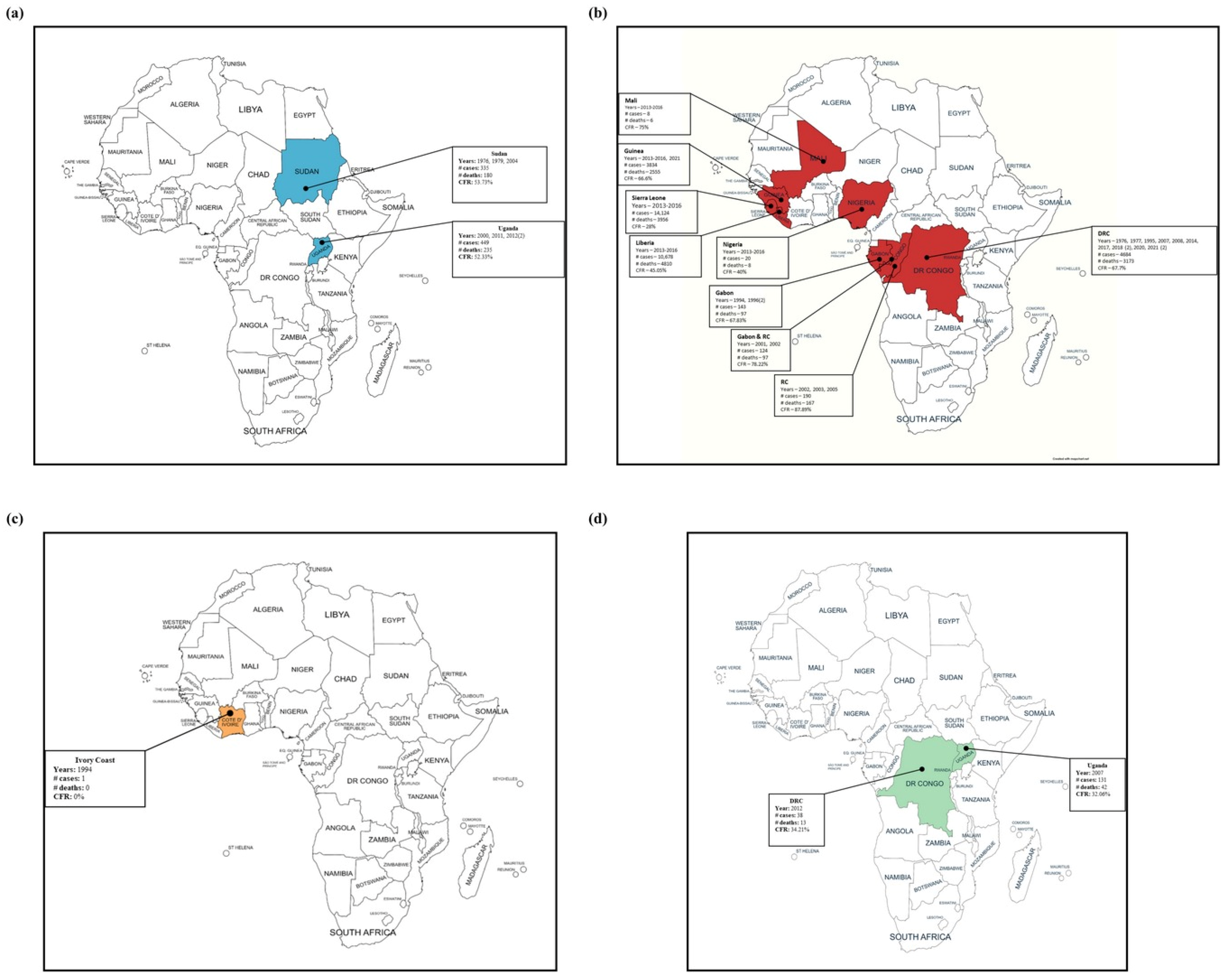
2. Epidemiology
2.1. Democratic Republic of the Congo

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Country | % Mortality (Deaths/Cases) | Reference | Year | Country | % Mortality (Deaths/Cases) | Reference |
|---|---|---|---|---|---|---|---|
| Ebola Virus (before 2014) | Ebola Virus (2014 onwards) | ||||||
| 1976 | DRC * | 88.05 (280/318) | [1,32] | 2013–2016 | Guinea | 66.72 (2543/3811) | [15] |
| 1977 | DRC | 100 (1/1) | [1,33] | Liberia | 45.04 (4810/10,678) | [15] | |
| 1994 | Gabon | 59.62 (31/52) | [1,31] | Sierra Leone | 28 (3956/14,124) | [15] | |
| 1995 | DRC | 79.37 (250/315) | [1,34] | Mali | 75 (6/8) | [36] | |
| 1996 | Gabon | 67.74 (21/31) | [1,31] | Nigeria | 40 (8/20) | [36] | |
| 1996 *** | Gabon | 75 (45/60) | [1,31] | Senegal | 0 (0/1) | [36] | |
| 2001 | Gabon/RC ** | 81.54 (53/65) | [1,37] | 2014 *** | DRC | 71.01 (49/69) | [1,38] |
| 2002 | Gabon/RC | 74.58 (44/59) | [1] | 2017 | DRC | 50 (4/8) | [1,39,40] |
| 2002 *** | RC | 89.51 (128/143) | [1,41] | 2018 *** | DRC | 61.11 (33/54) | [1] |
| 2003 | RC | 82.86 (29/35) | [1,42,43] | 2018 | DRC | 65.91 (2287/3470) | [1,8] |
| 2005 | RC | 83.33 (10/12) | [1] | 2020 | DRC | 42.31 (55/130) | [1] |
| 2007 | DRC | 70.83 (187/264) | [1,26] | 2021 | DRC | 50 (6/12) | [1,44] |
| 2008 | DRC | 46.88 (15/32) | [1] | 2021 | Guinea | 52.17 (12/23) | [1,10,45] |
| 2021 *** | DRC | 54.55 (6/11) | [1,44] | ||||
| Year | Country | % Mortality (Deaths/Cases) | Reference |
|---|---|---|---|
| SUDV | |||
| 1976 | South Sudan | 53.17 (151/284) | [4] |
| 1979 | South Sudan | 64.71 (22/34) | [54] |
| 2000 | Uganda | 52.71 (224/425) | [55] |
| 2004 | South Sudan | 41.18 (7/17) | [1] |
| 2011 | Uganda | 100 (1/1) | [56] |
| 2012 | Uganda | 36.36 (4/11) | [57] |
| 2012 | Uganda | 50 (3/6) | [57] |
| TAFV | |||
| 1994 | Ivory Coast | 0 (0/1) | [12] |
| BDBV | |||
| 2007 | Uganda | 32.06 (42/131) | [1,13] |
| 2012 | DRC | 34.21 (13/38) | [48] |
2.2. Gabon and the Republic of the Congo
2.3. Uganda
2.4. South Sudan
2.5. West African Countries
2.6. Other Outbreaks and Incidents
3. Etiology of Ebolavirus Infections
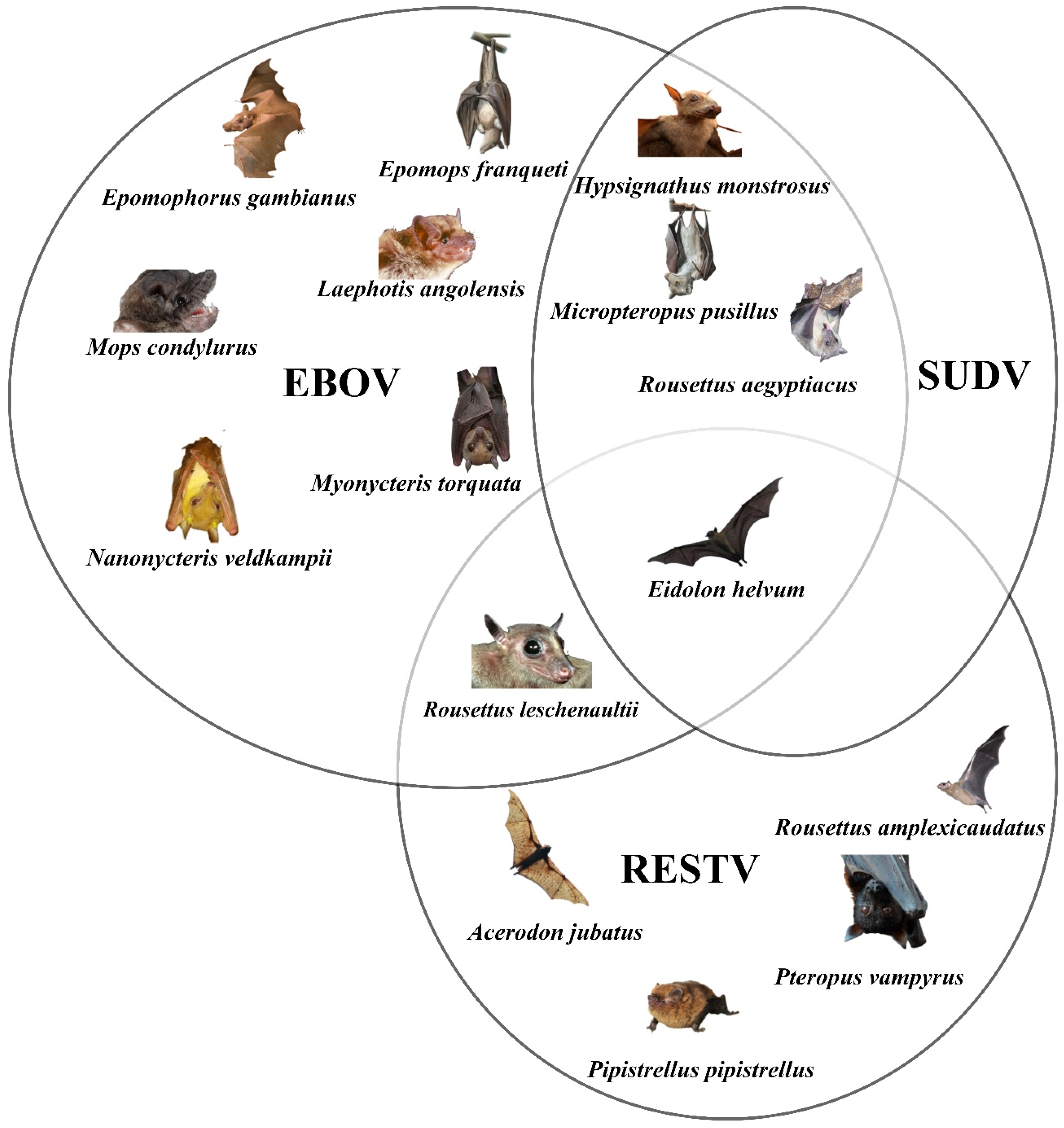
3.1. Bats as the Source of Infection
3.2. Bushmeat and NHPs as the Source of Infection
4. Potential Reservoirs
4.1. Bats
4.2. Other Potential Reservoirs
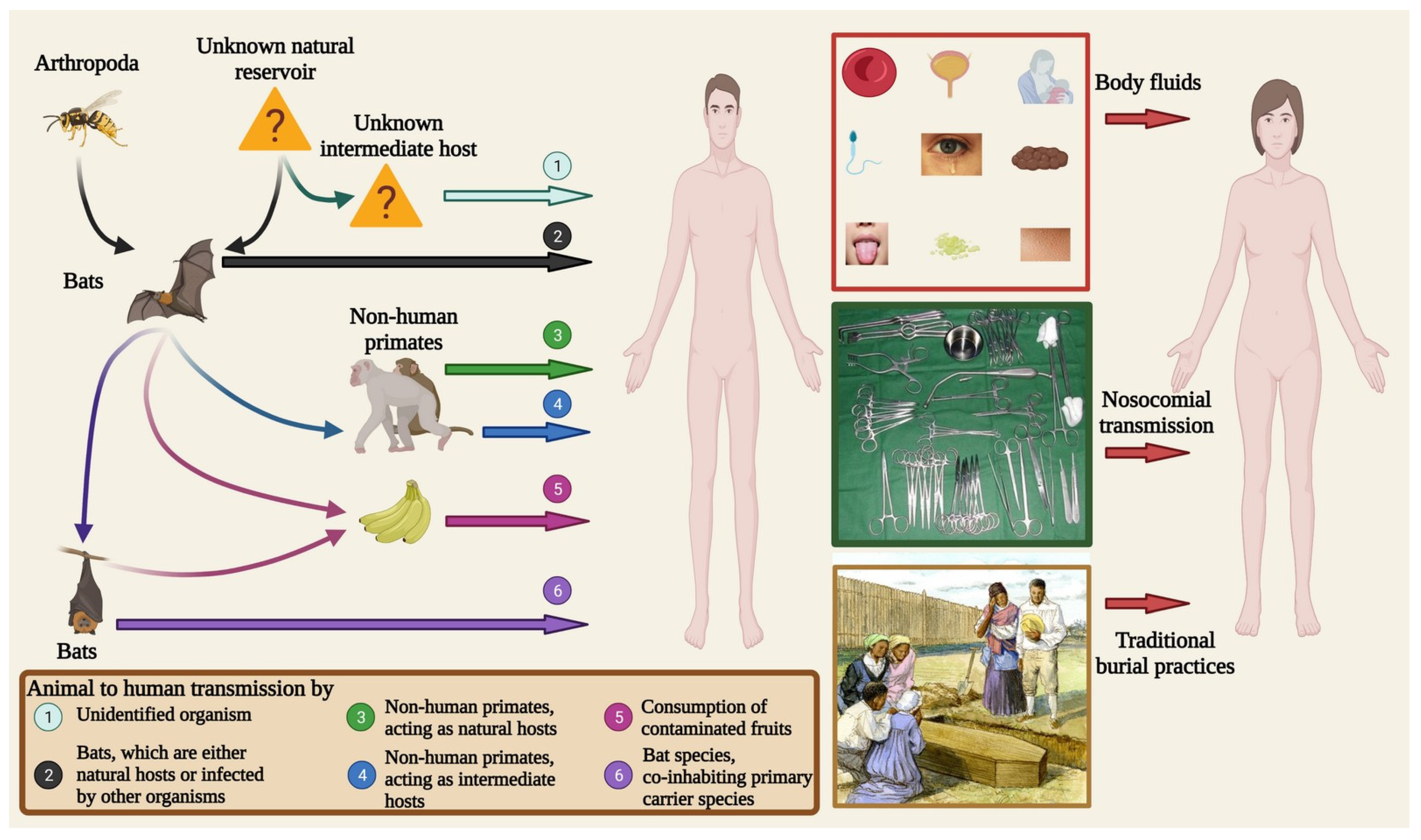
5. Transmission
5.1. Reservoir-to-Animal Transmission
5.2. Reservoir/Secondary Host-to-Human Transmission
5.3. Human-to-Human Transmission
6. Ebolavirus: The African Healthcare Threat
7. Current Status of Vaccines and Drugs
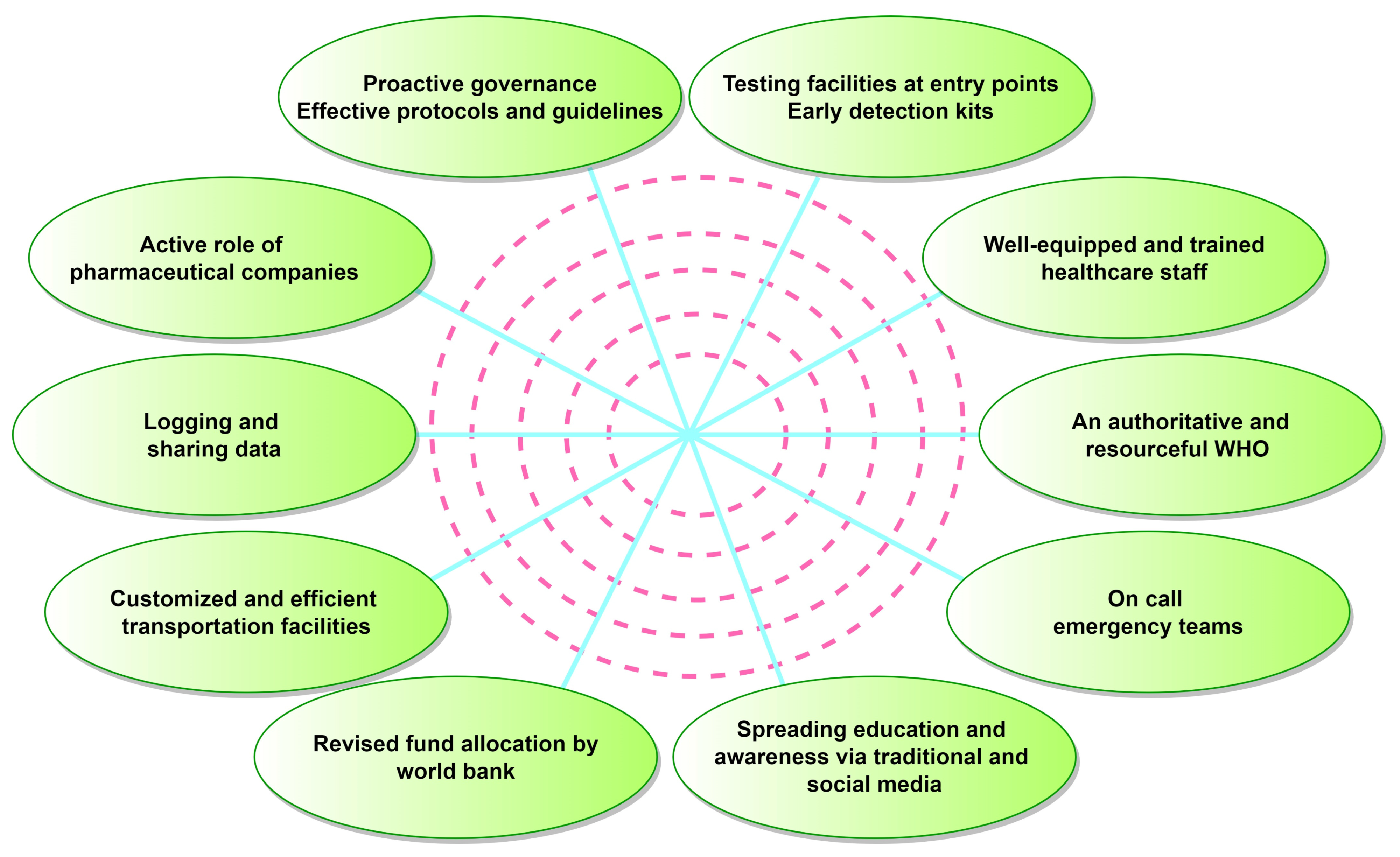
8. Future Challenges

Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CDC. Years of Ebola Virus Disease Outbreaks. Available online: https://www.cdc.gov/vhf/ebola/history/chronology.html (accessed on 14 May 2022).
- Jain, S.; Martynova, E.; Rizvanov, A.; Khaiboullina, S.; Baranwal, M. Structural and Functional Aspects of Ebola Virus Proteins. Pathogens 2021, 10, 1330. [Google Scholar] [CrossRef] [PubMed]
- Gałaś, A. The evolution of Ebola virus disease outbreaks. Folia Med. Crac. 2014, 54, 27–32. [Google Scholar]
- Shears, P.; O’Dempsey, T.J. Ebola virus disease in Africa: Epidemiology and nosocomial transmission. J. Hosp. Infect. 2015, 90, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jahrling, P.; Geisbert, T.; Johnson, E.; Peters, C.; Dalgard, D.; Hall, W. Preliminary report: Isolation of Ebola virus from monkeys imported to USA. Lancet 1990, 335, 502–505. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Jahrling, P.B.; Hanes, M.A.; Zack, P.M. Association of Ebola-related Reston virus particles and antigen with tissue lesions of monkeys imported to the United States. J. Comp. Pathol. 1992, 106, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, H.; Wahl-Jensen, V.; Jones, S.M.; Stroher, U. Ebola virus ecology: A continuing mystery. Trends Microbiol. 2004, 12, 433–437. [Google Scholar] [CrossRef]
- Shears, P.; Garavan, C. The 2018/19 Ebola epidemic the Democratic Republic of the Congo (DRC): Epidemiology, outbreak control, and conflict. Infect. Prev. Pract. 2020, 2, 100038. [Google Scholar] [CrossRef] [PubMed]
- Kerper, M.; Puckett, Y. Filovirus. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2020. [Google Scholar]
- Ohimain, E.I. Ecology of Ebolaviruses. Curr. Opin. Pharmacol. 2021, 60, 66–71. [Google Scholar] [CrossRef]
- Barrette, R.W.; Metwally, S.A.; Rowland, J.M.; Xu, L.; Zaki, S.R.; Nichol, S.T.; Rollin, P.E.; Towner, J.S.; Shieh, W.J.; Batten, B.; et al. Discovery of swine as a host for the Reston ebolavirus. Science 2009, 325, 204–206. [Google Scholar] [CrossRef]
- Le Guenno, B.; Formenty, P.; Wyers, M.; Gounon, P.; Walker, F.; Boesch, C. Isolation and partial characterisation of a new strain of Ebola virus. Lancet 1995, 345, 1271–1274. [Google Scholar] [CrossRef]
- Towner, J.S.; Sealy, T.K.; Khristova, M.L.; Albariño, C.G.; Conlan, S.; Reeder, S.A.; Quan, P.L.; Lipkin, W.I.; Downing, R.; Tappero, J.W.; et al. Newly discovered ebola virus associated with hemorrhagic fever outbreak in Uganda. PLoS Pathog. 2008, 4, e1000212. [Google Scholar] [CrossRef] [Green Version]
- Forbes, K.M.; Webala, P.W.; Jääskeläinen, A.J.; Abdurahman, S.; Ogola, J.; Masika, M.M.; Kivistö, I.; Alburkat, H.; Plyusnin, I.; Levanov, L.; et al. Bombali Virus in Mops condylurus Bat, Kenya. Emerg. Infect. Dis. 2019, 25, 955–957. [Google Scholar] [CrossRef]
- Shultz, J.M.; Espinel, Z.; Espinola, M.; Rechkemmer, A. Distinguishing epidemiological features of the 2013–2016 West Africa Ebola virus disease outbreak. Disaster Health 2016, 3, 78–88. [Google Scholar] [CrossRef]
- Gostin, L.O.; Friedman, E.A. A retrospective and prospective analysis of the west African Ebola virus disease epidemic: Robust national health systems at the foundation and an empowered WHO at the apex. Lancet 2015, 385, 1902–1909. [Google Scholar] [CrossRef]
- Tariq, A.; Roosa, K.; Mizumoto, K.; Chowell, G. Assessing reporting delays and the effective reproduction number: The Ebola epidemic in DRC, May 2018-January 2019. Epidemics 2019, 26, 128–133. [Google Scholar] [CrossRef]
- Kock, R.; Begovoeva, M.; Ansumana, R.; Suluku, R. Searching for the source of Ebola: The elusive factors driving its spillover into humans during the West African outbreak of 2013–2016. OIE Sci. Tech. Rev. 2019, 38, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Ohimain, E.I. Ecology of Ebolavirus: A Review of Current Knowledge, Speculations and Future Research Directions. Virology 2016, 1, 555564. [Google Scholar] [CrossRef]
- Reed, P.E.; Mulangu, S.; Cameron, K.N.; Ondzie, A.U.; Joly, D.; Bermejo, M.; Rouquet, P.; Fabozzi, G.; Bailey, M.; Shen, Z. A new approach for monitoring ebolavirus in wild great apes. PLoS Negl. Trop. Dis. 2014, 8, e3143. [Google Scholar] [CrossRef] [PubMed]
- Pourrut, X.; Kumulungui, B.; Wittmann, T.; Moussavou, G.; Delicat, A.; Yaba, P.; Nkoghe, D.; Gonzalez, J.P.; Leroy, E.M. The natural history of Ebola virus in Africa. Microbes Infect. 2005, 7, 1005–1014. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.P. Outbreaks of Ebola virus disease in Africa: The beginnings of a tragic saga. J. Venom. Anim. Toxins Incl. Trop. Dis. 2014, 20, 44. [Google Scholar] [CrossRef] [PubMed]
- Groseth, A.; Feldmann, H.; Strong, J.E. The ecology of Ebola virus. Trends Microbiol. 2007, 15, 408–416. [Google Scholar] [CrossRef]
- Lawrence, P.; Danet, N.; Reynard, O.; Volchkova, V.; Volchkov, V. Human transmission of Ebola virus. Curr. Opin. Virol. 2017, 22, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.; Qiu, X.; Richardson, J.S.; Cutts, T.; Collignon, B.; Gren, J.; Aviles, J.; Embury-Hyatt, C.; Kobinger, G.P. Ebola Virus Transmission in Guinea Pigs. J. Virol. 2015, 89, 1314–1323. [Google Scholar] [CrossRef]
- Leroy, E.M.; Epelboin, A.; Mondonge, V.; Pourrut, X.; Gonzalez, J.P.; Muyembe-Tamfum, J.J.; Formenty, P. Human Ebola outbreak resulting from direct exposure to fruit bats in Luebo, Democratic Republic of Congo, 2007. Vector Borne Zoonotic Dis. 2009, 9, 723–728. [Google Scholar] [CrossRef]
- Wong, S.S.; Wong, S.C. Ebola virus disease in nonendemic countries. J. Formos. Med. Assoc. Taiwan Yi Zhi 2015, 114, 384–398. [Google Scholar] [CrossRef] [PubMed]
- Vetter, P.; Fischer, W.A., 2nd; Schibler, M.; Jacobs, M.; Bausch, D.G.; Kaiser, L. Ebola Virus Shedding and Transmission: Review of Current Evidence. J. Infect. Dis. 2016, 214, S177–S184. [Google Scholar] [CrossRef] [PubMed]
- Jemimah, C.; Sheeba, S.L. Analysis of bodily fluids and fomites in transmission of ebola virus using bigdata. Procedia Comput. Sci. 2016, 92, 56–62. [Google Scholar] [CrossRef]
- Thorson, A.; Formenty, P.; Lofthouse, C.; Broutet, N. Systematic review of the literature on viral persistence and sexual transmission from recovered Ebola survivors: Evidence and recommendations. BMJ Open 2016, 6, e008859. [Google Scholar] [CrossRef]
- Georges, A.J.; Leroy, E.M.; Renaut, A.A.; Benissan, C.T.; Nabias, R.J.; Ngoc, M.T.; Obiang, P.I.; Lepage, J.P.; Bertherat, E.J.; Bénoni, D.D.; et al. Ebola hemorrhagic fever outbreaks in Gabon, 1994-1997: Epidemiologic and health control issues. J. Infect. Dis. 1999, 179 (Suppl. S1), S65–S75. [Google Scholar] [CrossRef]
- International Commission. Ebola haemorrhagic fever in Zaire, 1976. Bull. World Health Organ. 1978, 56, 271–293. [Google Scholar]
- Heymann, D.L.; Weisfeld, J.S.; Webb, P.A.; Johnson, K.M.; Cairns, T.; Berquist, H. Ebola hemorrhagic fever: Tandala, Zaire, 1977–1978. J. Infect. Dis. 1980, 142, 372–376. [Google Scholar] [CrossRef]
- Khan, A.S.; Tshioko, F.K.; Heymann, D.L.; Le Guenno, B.; Nabeth, P.; Kerstiëns, B.; Fleerackers, Y.; Kilmarx, P.H.; Rodier, G.R.; Nkuku, O.; et al. The reemergence of Ebola hemorrhagic fever, Democratic Republic of the Congo, 1995. Commission de Lutte contre les Epidémies à Kikwit. J. Infect. Dis. 1999, 179 (Suppl. S1), S76–S86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Epidemic and Pandemic Alert and Response (EPR). Available online: https://web.archive.org/web/20090219160037/http://www.who.int/csr/don/2009_02_17/en/index.html (accessed on 28 May 2022).
- Bell, B.P.; Damon, I.K.; Jernigan, D.B.; Kenyon, T.A.; Nichol, S.T.; O’Connor, J.P.; Tappero, J.W. Overview, Control Strategies, and Lessons Learned in the CDC Response to the 2014–2016 Ebola Epidemic. MMWR Suppl. 2016, 65, 4–11. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Outbreak (s) of Ebola haemorrhagic fever, Congo and Gabon, October 2001–July 2002. Wkly. Epidemiol. Rec. Relev. Épidémiologique Hebd. 2003, 78, 223–228. [Google Scholar]
- Maganga, G.D.; Kapetshi, J.; Berthet, N.; Kebela Ilunga, B.; Kabange, F.; Mbala Kingebeni, P.; Mondonge, V.; Muyembe, J.J.; Bertherat, E.; Briand, S.; et al. Ebola virus disease in the Democratic Republic of Congo. N. Engl. J. Med. 2014, 371, 2083–2091. [Google Scholar] [CrossRef]
- Gryseels, S.; Mbala-Kingebeni, P.; Akonda, I.; Angoyo, R.; Ayouba, A.; Baelo, P.; Mukadi, D.B.; Bugentho, E.; Bushmaker, T.; Butel, C. Role of wildlife in emergence of Ebola virus in Kaigbono (Likati), Democratic Republic of the Congo, 2017. Emerg. Infect. Dis. 2020, 26, 2205. [Google Scholar] [CrossRef]
- Nsio, J.; Kapetshi, J.; Makiala, S.; Raymond, F.; Tshapenda, G.; Boucher, N.; Corbeil, J.; Okitandjate, A.; Mbuyi, G.; Kiyele, M.; et al. 2017 Outbreak of Ebola Virus Disease in Northern Democratic Republic of Congo. J. Infect. Dis. 2020, 221, 701–706. [Google Scholar] [CrossRef]
- Formenty, P.; Libama, F.; Epelboin, A.; Allarangar, Y.; Leroy, E.; Moudzeo, H.; Tarangonia, P.; Molamou, A.; Lenzi, M.; Ait-Ikhlef, K.; et al. Outbreak of Ebola hemorrhagic fever in the Republic of the Congo, 2003: A new strategy? Med. Trop. Rev. Corps Sante Colonial 2003, 63, 291–295. [Google Scholar]
- Boumandouki, P.; Formenty, P.; Epelboin, A.; Campbell, P.; Atsangandoko, C.; Allarangar, Y.; Leroy, E.; Kone, M.; Molamou, A.; Dinga-Longa, O. Prise en charge des malades et des défunts lors de lépidémie de fievre hémorragique due au virus Ebola d’octobre adécembre 2003 au Congo. Bull. Soc. Pathol. Exot. 2005, 98, 218–223. [Google Scholar]
- Kuhn, J. Filoviruses: A Compendium of 40 Years of Epidemiological, Clinical, and Laboratory Studies; Springer: New York, NY, USA, 2008. [Google Scholar]
- Lacroix, A.; Mbala Kingebeni, P.; Ndimbo Kumugo, S.P.; Lempu, G.; Butel, C.; Serrano, L.; Vidal, N.; Thaurignac, G.; Esteban, A.; Mukadi Bamuleka, D.; et al. Investigating the Circulation of Ebola Viruses in Bats during the Ebola Virus Disease Outbreaks in the Equateur and North Kivu Provinces of the Democratic Republic of Congo from 2018. Pathogens 2021, 10, 557. [Google Scholar] [CrossRef]
- Adepoju, P. Ebola returns to Guinea and DR Congo. Lancet 2021, 397, 781. [Google Scholar] [CrossRef]
- Hulseberg, C.E.; Kumar, R.; Di Paola, N.; Larson, P.; Nagle, E.R.; Richardson, J.; Hanson, J.; Wauquier, N.; Fair, J.N.; Makuwa, M. Molecular analysis of the 2012 Bundibugyo virus disease outbreak. Cell Rep. Med. 2021, 2, 100351. [Google Scholar] [CrossRef] [PubMed]
- Epelboin, A. Rapport de mission anthropologique sur l’épidémie d’Ebola: Isiro, RD Congo, 4 au 30 Septembre 2012. OMS 2012. [Google Scholar]
- Kratz, T.; Roddy, P.; Tshomba Oloma, A.; Jeffs, B.; Pou Ciruelo, D.; de la Rosa, O.; Borchert, M. Ebola virus disease outbreak in Isiro, Democratic Republic of the Congo, 2012: Signs and symptoms, management and outcomes. PLoS ONE 2015, 10, e0129333. [Google Scholar] [CrossRef] [PubMed]
- EBOLA VIRUS DISEASE. Available online: https://apps.who.int/iris/bitstream/handle/10665/273640/SITREP_EVD_DRC_20180807-eng.pdf?ua%C2%BC1 (accessed on 25 May 2022).
- Nsubuga, P.; Masiira, B.; Kihembo, C.; Byakika-Tusiime, J.; Ryan, C.; Nanyunja, M.; Kamadjeu, R.; Talisuna, A. Evaluation of the Ebola Virus Disease (EVD) preparedness and readiness program in Uganda: 2018 to 2019. Pan Afr. Med. J. 2021, 38, 130. [Google Scholar] [CrossRef] [PubMed]
- Ebola Outbreak in the Democratic Republic of the Congo Declared a Public Health Emergency of International Concern. Available online: https://www.who.int/news-room/detail/17-07-2019-ebola-outbreak-in-the-democratic-republic-of-the-congo-declared-a-public-health-emergency-of-international-concern (accessed on 14 May 2020).
- Keita, M.; Lucaccioni, H.; Ilumbulumbu, M.K.; Polonsky, J.; Nsio-Mbeta, J.; Panda, G.T.; Adikey, P.C.; Ngwama, J.K.; Tosalisana, M.K.; Diallo, B. Evaluation of Early Warning, Alert and Response System for Ebola Virus Disease, Democratic Republic of the Congo, 2018–2020. Emerg. Infect. Dis. 2021, 27, 2988. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J. Hoping to head off an epidemic, Congo turns to experimental Ebola vaccine. Sci. (News) 2018, 15. [Google Scholar] [CrossRef]
- Baron, R.C.; McCormick, J.B.; Zubeir, O.A. Ebola virus disease in southern Sudan: Hospital dissemination and intrafamilial spread. Bull. World Health Organ. 1983, 61, 997–1003. [Google Scholar]
- Centers for Disease Control and Prevention. Outbreak of Ebola hemorrhagic fever Uganda, August 2000–January 2001. MMWR Morb. Mortal. Wkly. Rep. 2001, 50, 73–77. [Google Scholar]
- Shoemaker, T.; MacNeil, A.; Balinandi, S.; Campbell, S.; Wamala, J.F.; McMullan, L.K.; Downing, R.; Lutwama, J.; Mbidde, E.; Ströher, U.; et al. Reemerging Sudan Ebola virus disease in Uganda, 2011. Emerg. Infect. Dis. 2012, 18, 1480–1483. [Google Scholar] [CrossRef]
- Albarino, C.; Shoemaker, T.; Khristova, M.; Wamala, J.; Muyembe, J.; Balinandi, S.; Tumusiime, A.; Campbell, S.; Cannon, D.; Gibbons, A. Genomic analysis of filoviruses associated with four viral hemorrhagic fever outbreaks in Uganda and the Democratic Republic of the Congo in 2012. Virology 2013, 442, 97–100. [Google Scholar] [CrossRef]
- Kupferschmidt, K. New Ebola outbreak likely sparked by a person infected 5 years ago. Science 2021, 372, 1342–1344. [Google Scholar]
- Nkoghe, D.; Kone, M.L.; Yada, A.; Leroy, E. A limited outbreak of Ebola haemorrhagic fever in Etoumbi, Republic of Congo, 2005. Trans. R. Soc. Trop. Med. Hyg. 2011, 105, 466–472. [Google Scholar] [CrossRef]
- Rouquet, P.; Froment, J.-M.; Bermejo, M.; Kilbourn, A.; Karesh, W.; Reed, P.; Kumulungui, B.; Yaba, P.; Délicat, A.; Rollin, P.E. Wild animal mortality monitoring and human Ebola outbreaks, Gabon and Republic of Congo, 2001–2003. Emerg. Infect. Dis. 2005, 11, 283. [Google Scholar] [CrossRef]
- Leroy, E.M.; Rouquet, P.; Formenty, P.; Souquiere, S.; Kilbourne, A.; Froment, J.-M.; Bermejo, M.; Smit, S.; Karesh, W.; Swanepoel, R. Multiple Ebola virus transmission events and rapid decline of central African wildlife. Science 2004, 303, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Lamunu, M.; Lutwama, J.J.; Kamugisha, J.; Opio, A.; Nambooze, J.; Ndayimirije, N.; Okware, S. Containing a haemorrhagic fever epidemic: The Ebola experience in Uganda (October 2000–January 2001). Int. J. Infect. Dis. 2004, 8, 27–37. [Google Scholar] [CrossRef]
- Okware, S.I.; Omaswa, F.G.; Zaramba, S.; Opio, A.; Lutwama, J.J.; Kamugisha, J.; Rwaguma, E.B.; Kagwa, P.; Lamunu, M. An outbreak of Ebola in Uganda. Trop. Med. Int. Health TM IH 2002, 7, 1068–1075. [Google Scholar] [CrossRef]
- Wamala, J.F.; Lukwago, L.; Malimbo, M.; Nguku, P.; Yoti, Z.; Musenero, M.; Amone, J.; Mbabazi, W.; Nanyunja, M.; Zaramba, S. Ebola hemorrhagic fever associated with novel virus strain, Uganda, 2007–2008. Emerg. Infect. Dis. 2010, 16, 1087. [Google Scholar] [CrossRef]
- Dixon, M.G.; Schafer, I.J. Ebola viral disease outbreak—West Africa, 2014. MMWR Morb. Mortal. Wkly. Rep. 2014, 63, 548–551. [Google Scholar] [PubMed]
- Baize, S.; Pannetier, D.; Oestereich, L.; Rieger, T.; Koivogui, L.; Magassouba, N.F.; Soropogui, B.; Sow, M.S.; Keïta, S.; De Clerck, H. Emergence of Zaire Ebola virus disease in Guinea. N. Engl. J. Med. 2014, 371, 1418–1425. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO Statement on the Meeting of the International Health Regulations Emergency Committee Regarding the 2014 Ebola Outbreak in West Africa. Available online: https://web.archive.org/web/20140808155521/http://www.who.int/mediacentre/news/statements/2014/ebola-20140808/en/ (accessed on 28 May 2022).
- WHO. Situation Report Ebola Virus Disease. Available online: http://apps.who.int/iris/bitstream/handle/10665/208883/ebolasitrep_10Jun2016_eng.pdf;jsessionid=E4865AEE56C29E7B875648B29CDA8D84?sequence=1 (accessed on 28 May 2022).
- Emond, R.T.; Evans, B.; Bowen, E.T.; Lloyd, G. A case of Ebola virus infection. Br. Med. J. 1977, 2, 541–544. [Google Scholar] [CrossRef] [PubMed]
- WHO. 1996—South Africa. Available online: https://www.who.int/emergencies/disease-outbreak-news/item/1996_11_22-en (accessed on 28 May 2022).
- Borisevich, I.V.; Markin, V.A.; Firsova, I.V.; Evseev, A.A.; Khamitov, R.A.; Maksimov, V.A. Hemorrhagic (Marburg, Ebola, Lassa, and Bolivian) fevers: Epidemiology, clinical pictures, and treatment. Vopr. Virusol. 2006, 51, 8–16. [Google Scholar] [PubMed]
- Akinfeyeva, L.; Aksyonova, O.; Vasilyevich, I.; Ginko, Z.; Zarkov, K.; Zubavichene, N.; Katkova, L.; Kuzovlev, O.; Kuzubov, V.; Lokteva, L. A case of Ebola hemorrhagic fever. Infektsionnye Bolezn. 2005, 3, 85–88. [Google Scholar]
- RTVE. Muere el Religioso Español Miguel Pajares a Causa del Ébola. Available online: https://www.rtve.es/noticias/20140812/muere-miguel-pajares-ebola/991420.shtml (accessed on 28 May 2022).
- Peterson, A.T.; Bauer, J.T.; Mills, J.N. Ecologic and geographic distribution of filovirus disease. Emerg. Infect. Dis. 2004, 10, 40–47. [Google Scholar] [CrossRef] [PubMed]
- White, L.J. Patterns of fruit-fall phenology in the Lopé Reserve, Gabon. J. Trop. Ecol. 1994, 10, 289–312. [Google Scholar] [CrossRef]
- Leroy, E.M.; Kumulungui, B.; Pourrut, X.; Rouquet, P.; Hassanin, A.; Yaba, P.; Délicat, A.; Paweska, J.T.; Gonzalez, J.-P.; Swanepoel, R. Fruit bats as reservoirs of Ebola virus. Nature 2005, 438, 575–576. [Google Scholar] [CrossRef]
- Marí Saéz, A.; Weiss, S.; Nowak, K.; Lapeyre, V.; Zimmermann, F.; Düx, A.; Kühl, H.S.; Kaba, M.; Regnaut, S.; Merkel, K.; et al. Investigating the zoonotic origin of the West African Ebola epidemic. EMBO Mol. Med. 2015, 7, 17–23. [Google Scholar] [CrossRef]
- Amman, B.R.; Swanepoel, R.; Nichol, S.T.; Towner, J.S. Ecology of Filoviruses. In Marburg-and Ebolaviruses: From Ecosystems to Molecules; Mühlberger, E., Hensley, L.L., Towner, J.S., Eds.; Springer International Publishing: Cham, Swizerland, 2017; pp. 23–61. [Google Scholar]
- Walsh, P.D.; Abernethy, K.A.; Bermejo, M.; Beyers, R.; De Wachter, P.; Akou, M.E.; Huijbregts, B.; Mambounga, D.I.; Toham, A.K.; Kilbourn, A.M. Catastrophic ape decline in western equatorial Africa. Nature 2003, 422, 611–614. [Google Scholar] [CrossRef]
- Irving, A.T.; Ahn, M.; Goh, G.; Anderson, D.E.; Wang, L.-F. Lessons from the host defences of bats, a unique viral reservoir. Nature 2021, 589, 363–370. [Google Scholar] [CrossRef]
- Pourrut, X.; Delicat, A.; Rollin, P.; Ksiazek, T.; Gonzalez, J.-P.; Leroy, E. Spatial and temporal patterns of Zaire ebolavirus antibody prevalence in the possible reservoir bat species. J. Infect. Dis. 2007, 196, S176–S183. [Google Scholar] [CrossRef] [Green Version]
- Olival, K.J.; Hayman, D.T. Filoviruses in bats: Current knowledge and future directions. Viruses 2014, 6, 1759–1788. [Google Scholar] [CrossRef] [PubMed]
- Pourrut, X.; Souris, M.; Towner, J.S.; Rollin, P.E.; Nichol, S.T.; Gonzalez, J.-P.; Leroy, E. Large serological survey showing cocirculation of Ebola and Marburg viruses in Gabonese bat populations, and a high seroprevalence of both viruses in Rousettus aegyptiacus. BMC Infect. Dis. 2009, 9, 159. [Google Scholar] [CrossRef]
- Hayman, D.T.; Emmerich, P.; Yu, M.; Wang, L.-F.; Suu-Ire, R.; Fooks, A.R.; Cunningham, A.A.; Wood, J.L. Long-term survival of an urban fruit bat seropositive for Ebola and Lagos bat viruses. PLoS ONE 2010, 5, e11978. [Google Scholar] [CrossRef]
- Hayman, D.T.; Yu, M.; Crameri, G.; Wang, L.-F.; Suu-Ire, R.; Wood, J.L.; Cunningham, A.A. Ebola virus antibodies in fruit bats, Ghana, West Africa. Emerg. Infect. Dis. 2012, 18, 1207. [Google Scholar] [CrossRef]
- Olival, K.J.; Islam, A.; Yu, M.; Anthony, S.J.; Epstein, J.H.; Khan, S.A.; Khan, S.U.; Crameri, G.; Wang, L.-F.; Lipkin, W.I. Ebola virus antibodies in fruit bats, Bangladesh. Emerg. Infect. Dis. 2013, 19, 270. [Google Scholar] [CrossRef] [PubMed]
- De Nys, H.M.; Kingebeni, P.M.; Keita, A.K.; Butel, C.; Thaurignac, G.; Villabona-Arenas, C.-J.; Lemarcis, T.; Geraerts, M.; Vidal, N.; Esteban, A. Survey of Ebola viruses in frugivorous and insectivorous bats in Guinea, Cameroon, and the Democratic Republic of the Congo, 2015–2017. Emerg. Infect. Dis. 2018, 24, 2228. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Zhang, Y.; Li, J.; Zhang, Y.; Wang, L.-F.; Shi, Z. Serological evidence of ebolavirus infection in bats, China. Virol. J. 2012, 9, 236. [Google Scholar] [CrossRef]
- Taniguchi, S.; Watanabe, S.; Masangkay, J.S.; Omatsu, T.; Ikegami, T.; Alviola, P.; Ueda, N.; Iha, K.; Fujii, H.; Ishii, Y. Reston Ebolavirus antibodies in bats, the Philippines. Emerg. Infect. Dis. 2011, 17, 1559. [Google Scholar] [CrossRef]
- Jayme, S.I.; Field, H.E.; de Jong, C.; Olival, K.J.; Marsh, G.; Tagtag, A.M.; Hughes, T.; Bucad, A.C.; Barr, J.; Azul, R.R.; et al. Molecular evidence of Ebola Reston virus infection in Philippine bats. Virol. J. 2015, 12, 107. [Google Scholar] [CrossRef]
- Ogawa, H.; Miyamoto, H.; Nakayama, E.; Yoshida, R.; Nakamura, I.; Sawa, H.; Ishii, A.; Thomas, Y.; Nakagawa, E.; Matsuno, K. Seroepidemiological prevalence of multiple species of filoviruses in fruit bats (Eidolon helvum) migrating in Africa. J. Infect. Dis. 2015, 212, S101–S108. [Google Scholar] [CrossRef]
- Karan, L.S.; Makenov, M.T.; Korneev, M.G.; Sacko, N.; Boumbaly, S.; Yakovlev, S.A.; Kourouma, K.; Bayandin, R.B.; Gladysheva, A.V.; Shipovalov, A.V. Bombali virus in Mops condylurus bats, Guinea. Emerg. Infect. Dis. 2019, 25, 1774. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, T.; Anthony, S.J.; Gbakima, A.; Bird, B.H.; Bangura, J.; Tremeau-Bravard, A.; Belaganahalli, M.N.; Wells, H.L.; Dhanota, J.K.; Liang, E. The discovery of Bombali virus adds further support for bats as hosts of ebolaviruses. Nat. Microbiol. 2018, 3, 1084–1089. [Google Scholar] [CrossRef]
- Moratelli, R.; Calisher, C.H. Bats and zoonotic viruses: Can we confidently link bats with emerging deadly viruses? Mem. Inst. Oswaldo Cruz 2015, 110, 1–22. [Google Scholar] [CrossRef] [PubMed]
- O’shea, T.J.; Cryan, P.M.; Cunningham, A.A.; Fooks, A.R.; Hayman, D.T.; Luis, A.D.; Peel, A.J.; Plowright, R.K.; Wood, J.L. Bat flight and zoonotic viruses. Emerg. Infect. Dis. 2014, 20, 741. [Google Scholar] [CrossRef] [PubMed]
- Pavlovich, S.S.; Lovett, S.P.; Koroleva, G.; Guito, J.C.; Arnold, C.E.; Nagle, E.R.; Kulcsar, K.; Lee, A.; Thibaud-Nissen, F.; Hume, A.J. The Egyptian rousette genome reveals unexpected features of bat antiviral immunity. Cell 2018, 173, 1098–1110.e18. [Google Scholar] [CrossRef]
- Feldmann, H.; Geisbert, T.W. Ebola haemorrhagic fever. Lancet 2011, 377, 849–862. [Google Scholar] [CrossRef]
- Leendertz, S.A.J.; Gogarten, J.F.; Düx, A.; Calvignac-Spencer, S.; Leendertz, F.H. Assessing the evidence supporting fruit bats as the primary reservoirs for Ebola viruses. EcoHealth 2016, 13, 18–25. [Google Scholar] [CrossRef]
- Paweska, J.T.; Storm, N.; Grobbelaar, A.A.; Markotter, W.; Kemp, A.; Jansen van Vuren, P. Experimental inoculation of Egyptian fruit bats (Rousettus aegyptiacus) with Ebola virus. Viruses 2016, 8, 29. [Google Scholar] [CrossRef]
- Jones, M.E.; Schuh, A.J.; Amman, B.R.; Sealy, T.K.; Zaki, S.R.; Nichol, S.T.; Towner, J.S. Experimental inoculation of Egyptian rousette bats (Rousettus aegyptiacus) with viruses of the Ebolavirus and Marburgvirus genera. Viruses 2015, 7, 3420–3442. [Google Scholar] [CrossRef]
- Morvan, J.M.; Deubel, V.; Gounon, P.; Nakouné, E.; Barrière, P.; Murri, S.; Perpète, O.; Selekon, B.; Coudrier, D.; Gautier-Hion, A. Identification of Ebola virus sequences present as RNA or DNA in organs of terrestrial small mammals of the Central African Republic. Microbes Infect. 1999, 1, 1193–1201. [Google Scholar] [CrossRef]
- Breman, J.G.; Johnson, K.M.; van der Groen, G.; Robbins, C.B.; Szczeniowski, M.V.; Ruti, K.; Webb, P.A.; Meier, F.; Heymann, D.L. A search for Ebola virus in animals in the Democratic Republic of the Congo and Cameroon: Ecologic, virologic, and serologic surveys, 1979–1980. J. Infect. Dis. 1999, 179, S139–S147. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Zhang, W.; Cui, L.; Hua, X.; Wang, M.; Zeng, Q. Reston virus in domestic pigs in China. Arch. Virol. 2014, 159, 1129–1132. [Google Scholar] [CrossRef]
- Fischer, K.; Jabaty, J.; Suluku, R.; Strecker, T.; Groseth, A.; Fehling, S.K.; Balkema-Buschmann, A.; Koroma, B.; Schmidt, K.M.; Atherstone, C. Serological evidence for the circulation of ebolaviruses in pigs from Sierra Leone. J. Infect. Dis. 2018, 218, S305–S311. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.J.; Bushmaker, T.; Cameron, K.; Ondzie, A.; Niama, F.R.; Parra, H.-J.; Mombouli, J.-V.; Olson, S.H.; Munster, V.J.; Goldberg, T.L. Diverse RNA viruses of arthropod origin in the blood of fruit bats suggest a link between bat and arthropod viromes. Virology 2019, 528, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Dutto, M.; Bertero, M.; Petrosillo, N.; Pombi, M.; Otranto, D. Ebola virus and arthropods: A literature review and entomological consideration on the vector role. Bull. Soc. Pathol. Exot. 2016, 109, 244–247. [Google Scholar] [CrossRef]
- Leendertz, S.A.J. Testing New Hypotheses Regarding Ebolavirus Reservoirs. Viruses 2016, 8, 30. [Google Scholar] [CrossRef]
- Formenty, P.; Boesch, C.; Wyers, M.; Steiner, C.; Donati, F.; Dind, F.; Walker, F.; Le Guenno, B. Ebola virus outbreak among wild chimpanzees living in a rain forest of Cote d’Ivoire. J. Infect. Dis. 1999, 179, S120–S126. [Google Scholar] [CrossRef]
- Kamins, A.O.; Rowcliffe, J.M.; Ntiamoa-Baidu, Y.; Cunningham, A.A.; Wood, J.L.; Restif, O. Characteristics and risk perceptions of Ghanaians potentially exposed to bat-borne zoonoses through bushmeat. EcoHealth 2015, 12, 104–120. [Google Scholar] [CrossRef]
- Leroy, E.; Gonzalez, J.; Pourrut, X. Ebolavirus and other filoviruses. Wildl. Emerg. Zoonotic Dis. Biol. Circumst. Conseq. Cross-Species Transm. 2007, 315, 363–387. [Google Scholar]
- Tapanes, E.; Detwiler, K.M.; Cords, M. Bat predation by Cercopithecus monkeys: Implications for zoonotic disease transmission. EcoHealth 2016, 13, 405–409. [Google Scholar] [CrossRef]
- Bermejo, M.; Rodríguez-Teijeiro, J.D.; Illera, G.; Barroso, A.; Vilà, C.; Walsh, P.D. Ebola outbreak killed 5000 gorillas. Science 2006, 314, 1564. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, J.T.; Sovie, A.R.; Faller, C.R.; Monadjem, A.; Fletcher, R.J.; McCleery, R.A. Ebola spillover correlates with bat diversity. Eur. J. Wildl. Res. 2020, 66, 12. [Google Scholar] [CrossRef]
- Mickleburgh, S.; Waylen, K.; Racey, P. Bats as bushmeat: A global review. Oryx 2009, 43, 217–234. [Google Scholar] [CrossRef]
- Monadjem, A. Survival and roost-site selection in the African bat Nycteris thebaica (Chiroptera: Nycteridae) in Swaziland. Belg. J. Zool 2005, 135, 103–107. [Google Scholar]
- Monadjem, A.; Taylor, P.J.; Cotterill, F.; Schoeman, M. Bats of Central and Southern Africa: A Biogeographic and Taxonomic Synthesis; Wits University Press: Johannesbg, South Africa, 2010; Volume 459, p. 14. [Google Scholar]
- Feldmann, H.; Jones, S.; Klenk, H.D.; Schnittler, H.J. Ebola virus: From discovery to vaccine. Nat. Rev. Immunol. 2003, 3, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Monath, T.P. Ecology of Marburg and Ebola viruses: Speculations and directions for future research. J. Infect. Dis. 1999, 179, S127–S138. [Google Scholar] [CrossRef] [PubMed]
- Bertherat, E.; Renaut, A.; Nabias, R.; Dubreuil, G.; Georges-Courbot, M.-C. Leptospirosis and Ebola virus infection in five gold-panning villages in northeastern Gabon. Am. J. Trop. Med. Hyg. 1999, 60, 610–615. [Google Scholar] [CrossRef]
- Smith, I.; Wang, L.-F. Bats and their virome: An important source of emerging viruses capable of infecting humans. Curr. Opin. Virol. 2013, 3, 84–91. [Google Scholar] [CrossRef]
- Narayanan, R. Ebola-associated genes in the human genome: Implications for novel targets. MOJ Proteom. Bioinform 2014, 1, 00032. [Google Scholar] [CrossRef]
- Lever, R.A.; Whitty, C.J. Ebola virus disease: Emergence, outbreak and future directions. Br. Med. Bull. 2016, 117, 95–106. [Google Scholar] [CrossRef]
- Rojas, M.; Monsalve, D.M.; Pacheco, Y.; Acosta-Ampudia, Y.; Ramírez-Santana, C.; Ansari, A.A.; Gershwin, M.E.; Anaya, J.M. Ebola virus disease: An emerging and re-emerging viral threat. J. Autoimmun. 2020, 106, 102375. [Google Scholar] [CrossRef]
- Kerstiëns, B.; Matthys, F. Interventions to control virus transmission during an outbreak of Ebola hemorrhagic fever: Experience from Kikwit, Democratic Republic of the Congo, 1995. J. Infect. Dis. 1999, 179, S263–S267. [Google Scholar] [CrossRef] [Green Version]
- Leroy, E.M.; Baize, S.; Volchkov, V.E.; Fisher-Hoch, S.P.; Georges-Courbot, M.C.; Lansoud-Soukate, J.; Capron, M.; Debré, P.; McCormick, J.B.; Georges, A.J. Human asymptomatic Ebola infection and strong inflammatory response. Lancet 2000, 355, 2210–2215. [Google Scholar] [CrossRef] [PubMed]
- Timothy, J.W.; Hall, Y.; Akoi-Boré, J.; Diallo, B.; Tipton, T.R.; Bower, H.; Strecker, T.; Glynn, J.R.; Carroll, M.W. Early transmission and case fatality of Ebola virus at the index site of the 2013–2016 west African Ebola outbreak: A cross-sectional seroprevalence survey. Lancet Infect. Dis. 2019, 19, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Golden, C.D.; Fernald, L.C.; Brashares, J.S.; Rasolofoniaina, B.R.; Kremen, C. Benefits of wildlife consumption to child nutrition in a biodiversity hotspot. Proc. Natl. Acad. Sci. USA 2011, 108, 19653–19656. [Google Scholar] [CrossRef]
- Bennett, E.L.; Blencowe, E.; Brandon, K.; Brown, D.; Burn, R.W.; Cowlishaw, G.; Davies, G.; Dublin, H.; Fa, J.E.; Milner-Gulland, E. Hunting for consensus: Reconciling bushmeat harvest, conservation, and development policy in West and Central Africa. Conserv. Biol. 2007, 21, 884–887. [Google Scholar] [CrossRef] [PubMed]
- Baudel, H.; De Nys, H.; Mpoudi Ngole, E.; Peeters, M.; Desclaux, A. Understanding Ebola virus and other zoonotic transmission risks through human–bat contacts: Exploratory study on knowledge, attitudes and practices in Southern Cameroon. Zoonoses Public Health 2019, 66, 288–295. [Google Scholar] [CrossRef]
- Fa, J.E.; Nasi, R.; van Vliet, N. Bushmeat, anthropogenic change, and human health in tropical rainforests: The case of the Ebola virus. St. Publique 2019, S1(HS), 107–114. [Google Scholar] [CrossRef]
- Khan, A.S.; Sesay, S.S.S. Seafood insecurity, bush meat consumption, and public health emergency in West Africa: Did we miss the early warning signs of an Ebola epidemic? Marit. Stud. 2015, 14, 3. [Google Scholar] [CrossRef]
- van Vliet, N.; Mbazza, P. Recognizing the multiple reasons for bushmeat consumption in urban areas: A necessary step toward the sustainable use of wildlife for food in Central Africa. Hum. Dimens. Wildl. 2011, 16, 45–54. [Google Scholar] [CrossRef]
- Sirén, A. Festival hunting by the Kichwa people in the Ecuadorian Amazon. J. Ethnobiol. 2012, 32, 30–50. [Google Scholar] [CrossRef]
- Timen, A.; Sprenger, M.; Edelstein, M.; Martin-Moreno, J.; McKee, M. The Ebola crisis: Perspectives from European Public Health. Eur. J. Public Health 2015, 25, 187–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiffany, A.; Dalziel, B.D.; Kagume Njenge, H.; Johnson, G.; Nugba Ballah, R.; James, D.; Wone, A.; Bedford, J.; McClelland, A. Estimating the number of secondary Ebola cases resulting from an unsafe burial and risk factors for transmission during the West Africa Ebola epidemic. PLoS Negl. Trop. Dis. 2017, 11, e0005491. [Google Scholar] [CrossRef] [PubMed]
- Azuonwu, O.; Chukwu, E.C. Emergence and Re-emergence of 2014 Ebola outbreak in Sub-Sahara Africa: ‘Challenges and lessons learned’ from Nigerian epidemic outbreak. Sch. J. App. Med. Sci. 2015, 3, 2802–2814. [Google Scholar]
- Jones, B.A.; Grace, D.; Kock, R.; Alonso, S.; Rushton, J.; Said, M.Y.; McKeever, D.; Mutua, F.; Young, J.; McDermott, J. Zoonosis emergence linked to agricultural intensification and environmental change. Proc. Natl. Acad. Sci. USA 2013, 110, 8399–8404. [Google Scholar] [CrossRef]
- Emergencies Preparedness, Response. Available online: https://www.who.int/csr/disease/ebola/one-year-report/factors/en/ (accessed on 14 May 2020).
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef]
- Allen, T.; Murray, K.A.; Zambrana-Torrelio, C.; Morse, S.S.; Rondinini, C.; Di Marco, M.; Breit, N.; Olival, K.J.; Daszak, P. Global hotspots and correlates of emerging zoonotic diseases. Nat. Commun. 2017, 8, 1124. [Google Scholar] [CrossRef]
- Van Vliet, N.; Moreno Calderón, J.L.; Gomez, J.; Zhou, W.; Fa, J.E.; Golden, C.; Nobrega Alves, R.R.; Nasi, R. Bushmeat and human health: Assessing the evidence in tropical and sub-tropical forests. Etnobiol. Conserv. 2017, 6, 1–45. [Google Scholar] [CrossRef]
- Munoz-Fontela, C.; McElroy, A.K. Ebola Virus Disease in Humans: Pathophysiology and Immunity. Curr. Top. Microbiol. Immunol. 2017, 411, 141–169. [Google Scholar] [CrossRef]
- Gignoux, E.; Azman, A.S.; de Smet, M.; Azuma, P.; Massaquoi, M.; Job, D.; Tiffany, A.; Petrucci, R.; Sterk, E.; Potet, J.; et al. Effect of Artesunate-Amodiaquine on Mortality Related to Ebola Virus Disease. N. Engl. J. Med. 2016, 374, 23–32. [Google Scholar] [CrossRef]
- McMahon, S.A.; Ho, L.S.; Brown, H.; Miller, L.; Ansumana, R.; Kennedy, C.E. Healthcare providers on the frontlines: A qualitative investigation of the social and emotional impact of delivering health services during Sierra Leone’s Ebola epidemic. Health Policy Plan. 2016, 31, 1232–1239. [Google Scholar] [CrossRef]
- Jegede, A.S. What led to the Nigerian boycott of the polio vaccination campaign? PLoS Med. 2007, 4, e73. [Google Scholar] [CrossRef]
- Lenzer, J. Pfizer settles with victims of Nigerian drug trial. BMJ 2011, 343, d5268. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, Y.; Caposio, P.; Parkins, C.J.; Botto, S.; Messaoudi, I.; Cicin-Sain, L.; Feldmann, H.; Jarvis, M.A. A replicating cytomegalovirus-based vaccine encoding a single Ebola virus nucleoprotein CTL epitope confers protection against Ebola virus. PLoS Negl. Trop. Dis. 2011, 5, e1275. [Google Scholar] [CrossRef]
- Wilson, J.A.; Hart, M.K. Protection from Ebola virus mediated by cytotoxic T lymphocytes specific for the viral nucleoprotein. J. Virol. 2001, 75, 2660–2664. [Google Scholar] [CrossRef] [PubMed]
- Enterlein, S.; Warfield, K.L.; Swenson, D.L.; Stein, D.A.; Smith, J.L.; Gamble, C.S.; Kroeker, A.D.; Iversen, P.L.; Bavari, S.; Mühlberger, E. VP35 knockdown inhibits Ebola virus amplification and protects against lethal infection in mice. Antimicrob. Agents Chemother. 2006, 50, 984–993. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, W.M.; Carter, W.A. The quest for effective Ebola treatment: Ebola VP35 is an evidence-based target for dsRNA drugs. Emerg. Microbes Infect. 2014, 3, e77. [Google Scholar] [CrossRef] [PubMed]
- Stahelin, R.V. Could the Ebola virus matrix protein VP40 be a drug target? Expert Opin. Ther. Targets 2014, 18, 115–120. [Google Scholar] [CrossRef]
- Madara, J.J.; Han, Z.; Ruthel, G.; Freedman, B.D.; Harty, R.N. The multifunctional Ebola virus VP40 matrix protein is a promising therapeutic target. Future Virol. 2015, 10, 537–546. [Google Scholar] [CrossRef]
- Herbert, A.S.; Kuehne, A.I.; Barth, J.F.; Ortiz, R.A.; Nichols, D.K.; Zak, S.E.; Stonier, S.W.; Muhammad, M.A.; Bakken, R.R.; Prugar, L.I.; et al. Venezuelan equine encephalitis virus replicon particle vaccine protects nonhuman primates from intramuscular and aerosol challenge with ebolavirus. J. Virol. 2013, 87, 4952–4964. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Bailey, M.; Hensley, L.; Asiedu, C.; Geisbert, J.; Stanley, D.; Honko, A.; Johnson, J.; Mulangu, S.; Pau, M.G.; et al. Recombinant adenovirus serotype 26 (Ad26) and Ad35 vaccine vectors bypass immunity to Ad5 and protect nonhuman primates against ebolavirus challenge. J. Virol. 2011, 85, 4222–4233. [Google Scholar] [CrossRef] [PubMed]
- Hartlieb, B.; Modrof, J.; Mühlberger, E.; Klenk, H.D.; Becker, S. Oligomerization of Ebola virus VP30 is essential for viral transcription and can be inhibited by a synthetic peptide. J. Biol. Chem. 2003, 278, 41830–41836. [Google Scholar] [CrossRef] [PubMed]
- Warren, T.K.; Whitehouse, C.A.; Wells, J.; Welch, L.; Heald, A.E.; Charleston, J.S.; Sazani, P.; Iversen, P.L.; Bavari, S. A single phosphorodiamidate morpholino oligomer targeting VP24 protects rhesus monkeys against lethal Ebola virus infection. mBio 2015, 6, e02344-14. [Google Scholar] [CrossRef] [Green Version]
- Trunschke, M.; Conrad, D.; Enterlein, S.; Olejnik, J.; Brauburger, K.; Mühlberger, E. The L–VP35 and L–L interaction domains reside in the amino terminus of the Ebola virus L protein and are potential targets for antivirals. Virology 2013, 441, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Pushko, P.; Bray, M.; Ludwig, G.V.; Parker, M.; Schmaljohn, A.; Sanchez, A.; Jahrling, P.B.; Smith, J.F. Recombinant RNA replicons derived from attenuated Venezuelan equine encephalitis virus protect guinea pigs and mice from Ebola hemorrhagic fever virus. Vaccine 2000, 19, 142–153. [Google Scholar] [CrossRef]
- Warfield, K.L.; Swenson, D.L.; Olinger, G.G.; Kalina, W.V.; Aman, M.J.; Bavari, S. Ebola virus-like particle-based vaccine protects nonhuman primates against lethal Ebola virus challenge. J. Infect. Dis. 2007, 196 (Suppl. S2), S430–S437. [Google Scholar] [CrossRef]
- Marzi, A.; Ebihara, H.; Callison, J.; Groseth, A.; Williams, K.J.; Geisbert, T.W.; Feldmann, H. Vesicular stomatitis virus–based Ebola vaccines with improved cross-protective efficacy. J. Infect. Dis. 2011, 204, S1066–S1074. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.A.; Bray, M.; Bakken, R.; Hart, M.K. Vaccine potential of Ebola virus VP24, VP30, VP35, and VP40 proteins. Virology 2001, 286, 384–390. [Google Scholar] [CrossRef]
- Jain, S.; Baranwal, M. Conserved peptide vaccine candidates containing multiple Ebola nucleoprotein epitopes display interactions with diverse HLA molecules. Med. Microbiol. Immunol. 2019, 208, 227–238. [Google Scholar] [CrossRef]
- Jain, S.; Baranwal, M. Computational analysis in designing T cell epitopes enriched peptides of Ebola glycoprotein exhibiting strong binding interaction with HLA molecules. J. Theor. Biol. 2019, 465, 34–44. [Google Scholar] [CrossRef]
- Jain, S.; Baranwal, M. Conserved immunogenic peptides of Ebola glycoprotein elicit immune response in human peripheral blood mononuclear cells. Microbiol. Immunol. 2021, 65, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-X.; Hou, L.-H.; Meng, F.-Y.; Wu, S.-P.; Hu, Y.-M.; Liang, Q.; Chu, K.; Zhang, Z.; Xu, J.-J.; Tang, R. Immunity duration of a recombinant adenovirus type-5 vector-based Ebola vaccine and a homologous prime-boost immunisation in healthy adults in China: Final report of a randomised, double-blind, placebo-controlled, phase 1 trial. Lancet Glob. Health 2017, 5, e324–e334. [Google Scholar] [CrossRef]
- Dolzhikova, I.V.; Zubkova, O.V.; Tukhvatulin, A.I.; Dzharullaeva, A.S.; Tukhvatulina, N.M.; Shcheblyakov, D.V.; Shmarov, M.M.; Tokarskaya, E.A.; Simakova, Y.V.; Egorova, D.A.; et al. Safety and immunogenicity of GamEvac-Combi, a heterologous VSV- and Ad5-vectored Ebola vaccine: An open phase I/II trial in healthy adults in Russia. Hum. Vaccines Immunother. 2017, 13, 613–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.C.; Wurie, A.H.; Hou, L.H.; Liang, Q.; Li, Y.H.; Russell, J.B.; Wu, S.P.; Li, J.X.; Hu, Y.M.; Guo, Q.; et al. Safety and immunogenicity of a recombinant adenovirus type-5 vector-based Ebola vaccine in healthy adults in Sierra Leone: A single-centre, randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2017, 389, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Ollmann Saphire, E. A Vaccine against Ebola Virus. Cell 2020, 181, 6. [Google Scholar] [CrossRef] [PubMed]
- ERVEBO. Available online: https://www.fda.gov/vaccines-blood-biologics/ervebo (accessed on 14 July 2020).
- Markham, A. REGN-EB3: First approval. Drugs 2021, 81, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Lee, A. Ansuvimab: First Approval. Drugs 2021, 81, 595–598. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, H.; Feldmann, F.; Marzi, A. Ebola: Lessons on Vaccine Development. Annu. Rev. Microbiol. 2018, 72, 423–446. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, E.J.; Racine, T.; Kobinger, G.P. The ongoing evolution of antibody-based treatments for Ebola virus infection. Immunotherapy 2017, 9, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Grant-Klein, R.J.; Altamura, L.A.; Schmaljohn, C.S. Progress in recombinant DNA-derived vaccines for Lassa virus and filoviruses. Virus Res. 2011, 162, 148–161. [Google Scholar] [CrossRef] [PubMed]
- Croyle, M.A.; Patel, A.; Tran, K.N.; Gray, M.; Zhang, Y.; Strong, J.E.; Feldmann, H.; Kobinger, G.P. Nasal delivery of an adenovirus-based vaccine bypasses pre-existing immunity to the vaccine carrier and improves the immune response in mice. PLoS ONE 2008, 3, e3548. [Google Scholar] [CrossRef] [PubMed]
- Skwarczynski, M.; Toth, I. Peptide-based synthetic vaccines. Chem. Sci. 2016, 7, 842–854. [Google Scholar] [CrossRef] [PubMed]
- Mora-Rillo, M.; Arsuaga, M.; Ramírez-Olivencia, G.; de la Calle, F.; Borobia, A.M.; Sánchez-Seco, P.; Lago, M.; Figueira, J.C.; Fernández-Puntero, B.; Viejo, A.; et al. Acute respiratory distress syndrome after convalescent plasma use: Treatment of a patient with Ebola virus disease contracted in Madrid, Spain. Lancet Respir. Med. 2015, 3, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Uyeki, T.M.; Mehta, A.K.; Davey, R.T., Jr.; Liddell, A.M.; Wolf, T.; Vetter, P.; Schmiedel, S.; Grunewald, T.; Jacobs, M.; Arribas, J.R.; et al. Clinical Management of Ebola Virus Disease in the United States and Europe. N. Engl. J. Med. 2016, 374, 636–646. [Google Scholar] [CrossRef]
- Jain, S.; Khaiboullina, S.F.; Baranwal, M. Immunological Perspective for Ebola Virus Infection and Various Treatment Measures Taken to Fight the Disease. Pathogens 2020, 9, 850. [Google Scholar] [CrossRef]
- Mobula, L.M.; Samaha, H.; Yao, M.; Gueye, A.S.; Diallo, B.; Umutoni, C.; Anoko, J.; Lokonga, J.-P.; Minikulu, L.; Mossoko, M. Recommendations for the COVID-19 response at the national level based on lessons learned from the Ebola virus disease outbreak in the Democratic Republic of the Congo. Am. J. Trop. Med. Hyg. 2020, 103, 12. [Google Scholar] [CrossRef]
- Outbreak. Available online: http://www.pbs.org/wgbh/frontline/film/outbreak/credits/ (accessed on 14 May 2020).
- Quaglio, G.; Goerens, C.; Putoto, G.; Rübig, P.; Lafaye, P.; Karapiperis, T.; Dario, C.; Delaunois, P.; Zachariah, R. Ebola: Lessons learned and future challenges for Europe. Lancet Infect. Dis. 2016, 16, 259–263. [Google Scholar] [CrossRef]
- Ebola Crisis: AMA Criticises Australia’s Response to Virus Outbreak; West Africa Cases Exceed 10,000. Available online: https://www.abc.net.au/news/2014-10-25/ama-criticises-australias-ebola-response-as-chaotic/5841878?nw=0 (accessed on 14 May 2020).
- Karan, A.; Pogge, T. Ebola and the need for restructuring pharmaceutical incentives. J. Glob. Health 2015, 5, 010303. [Google Scholar] [CrossRef]
- Emrick, P.; Gentry, C.; Morowit, L. Ebola Virus Disease: International perspective on enhanced health surveillance, disposition of the dead, and their effect on isolation and quarantine practices. Disaster Mil. Med. 2016, 2, 13. [Google Scholar] [CrossRef]
- Cameron, A.; Meyer, A.; Faverjon, C.; Mackenzie, C. Quantification of the sensitivity of early detection surveillance. Transbound. Emerg. Dis. 2020, 67, 2532–2543. [Google Scholar] [CrossRef]
- Beam, E.L.; Schwedhelm, M.M.; Boulter, K.C.; Vasa, A.M.; Larson, L.; Cieslak, T.J.; Lowe, J.J.; Herstein, J.J.; Kratochvil, C.J.; Hewlett, A.L. Ebola Virus Disease: Clinical Challenges, Recognition, and Management. Nurs. Clin. N. Am. 2019, 54, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Madariaga, M.G. Ebola Virus Disease: A Perspective for the United States. Am. J. Med. 2015, 128, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Guidance on Air Medical Transport (AMT) for Patients with Ebola Virus Disease (EVD). Available online: https://www.cdc.gov/vhf/ebola/clinicians/emergency-services/air-medical-transport.html (accessed on 14 May 2020).
- Brearley, M.B.; Heaney, M.F.; Norton, I.N. Physiological responses of medical team members to a simulated emergency in tropical field conditions. Prehospital Disaster Med. 2013, 28, 139–144. [Google Scholar] [CrossRef]
- Leligdowicz, A.; Fischer, W.A., 2nd; Uyeki, T.M.; Fletcher, T.E.; Adhikari, N.K.; Portella, G.; Lamontagne, F.; Clement, C.; Jacob, S.T.; Rubinson, L.; et al. Ebola virus disease and critical illness. Crit. Care 2016, 20, 217. [Google Scholar] [CrossRef]
- Guidance on Personal Protective Equipment (PPE) to Be Used by Healthcare Workers during Management of Patients with Confirmed Ebola or Persons under Investigation (PUIs) for Ebola Who Are Clinically Unstable or Have Bleeding, Vomiting, or Diarrhea in U.S. Hospitals, Including Procedures for Donning and Doffing PPE. Available online: https://www.cdc.gov/vhf/ebola/healthcare-us/ppe/guidance.html (accessed on 14 May 2020).
- Lyons, P.; Winters, M.; Zeebari, Z.; Schmidt-Hellerau, K.; Sengeh, P.; Jalloh, M.B.; Jalloh, M.F.; Nordenstedt, H. Engaging religious leaders to promote safe burial practices during the 2014–2016 Ebola virus disease outbreak, Sierra Leone. Bull. World Health Organ. 2021, 99, 271. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, A.; Alonso Aguirre, A.; Jeggo, M.; Woods, R. Effective Coordination and Management of Emerging Infectious Diseases in Wildlife Populations. EcoHealth 2015, 12, 408–411. [Google Scholar] [CrossRef]
- Gortazar, C.; Reperant, L.A.; Kuiken, T.; de la Fuente, J.; Boadella, M.; Martínez-Lopez, B.; Ruiz-Fons, F.; Estrada-Peña, A.; Drosten, C.; Medley, G.; et al. Crossing the interspecies barrier: Opening the door to zoonotic pathogens. PLoS Pathog. 2014, 10, e1004129. [Google Scholar] [CrossRef]
- Morse, S.S.; Mazet, J.A.; Woolhouse, M.; Parrish, C.R.; Carroll, D.; Karesh, W.B.; Zambrana-Torrelio, C.; Lipkin, W.I.; Daszak, P. Prediction and prevention of the next pandemic zoonosis. Lancet 2012, 380, 1956–1965. [Google Scholar] [CrossRef]
- Jacobsen, K.H.; Aguirre, A.A.; Bailey, C.L.; Baranova, A.V.; Crooks, A.T.; Croitoru, A.; Delamater, P.L.; Gupta, J.; Kehn-Hall, K.; Narayanan, A.; et al. Lessons from the Ebola Outbreak: Action Items for Emerging Infectious Disease Preparedness and Response. EcoHealth 2016, 13, 200–212. [Google Scholar] [CrossRef]
- Anyamba, A.; Chretien, J.-P.; Small, J.; Tucker, C.J.; Formenty, P.B.; Richardson, J.H.; Britch, S.C.; Schnabel, D.C.; Erickson, R.L.; Linthicum, K.J. Prediction of a Rift Valley fever outbreak. Proc. Natl. Acad. Sci. USA 2009, 106, 955–959. [Google Scholar] [CrossRef] [Green Version]





| Ebolavirus | DRC | Gabon and RC | Uganda | South Sudan | West Africa | Total Number of Outbreaks | Total Deaths/Cases | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Outbreaks | Deaths/Cases | Outbreaks | Deaths/Cases | Outbreaks | Deaths/Cases | Outbreaks | Deaths/Cases | Outbreaks | Deaths/Cases | |||
| EBOV | 12 | 3173/4684 | 8 | 361/457 | − | − | − | − | 2 | 11,337/28,675 | 22 | 14,871/33,816 |
| SUDV | − | − | − | − | 4 | 235/449 | 3 | 180/335 | − | − | 7 | 415/784 |
| BDBV | 1 | 13/38 | − | − | 1 | 42/131 | − | − | − | − | 2 | 55/169 |
| TAFV | − | − | − | − | − | − | − | − | 1 | 0/1 | 1 | 0/1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jain, S.; Khaiboullina, S.; Martynova, E.; Morzunov, S.; Baranwal, M. Epidemiology of Ebolaviruses from an Etiological Perspective. Pathogens 2023, 12, 248. https://doi.org/10.3390/pathogens12020248
Jain S, Khaiboullina S, Martynova E, Morzunov S, Baranwal M. Epidemiology of Ebolaviruses from an Etiological Perspective. Pathogens. 2023; 12(2):248. https://doi.org/10.3390/pathogens12020248
Chicago/Turabian StyleJain, Sahil, Svetlana Khaiboullina, Ekaterina Martynova, Sergey Morzunov, and Manoj Baranwal. 2023. "Epidemiology of Ebolaviruses from an Etiological Perspective" Pathogens 12, no. 2: 248. https://doi.org/10.3390/pathogens12020248
APA StyleJain, S., Khaiboullina, S., Martynova, E., Morzunov, S., & Baranwal, M. (2023). Epidemiology of Ebolaviruses from an Etiological Perspective. Pathogens, 12(2), 248. https://doi.org/10.3390/pathogens12020248