
Marketed Quinoa (Chenopodium quinoa Willd.) Seeds: A Mycotoxin-Free Matrix Contaminated by Mycotoxigenic Fungi

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Marketed Quinoa Seed Samples
2.2. Isolation and Molecular Identification of the Fungal Microorganisms Associated with the Marketed Quinoa Seed Samples
- for those isolates morphologically ascribed to the genus Cladosporium, the sequences of the ITS region combined with those of the actin (ACT) gene [33] were used;
2.3. Fungal Secondary Metabolites Analysis in the Marketed Quinoa Seed Samples and in Fungal Cultures
2.4. Statistical Analysis
3. Results
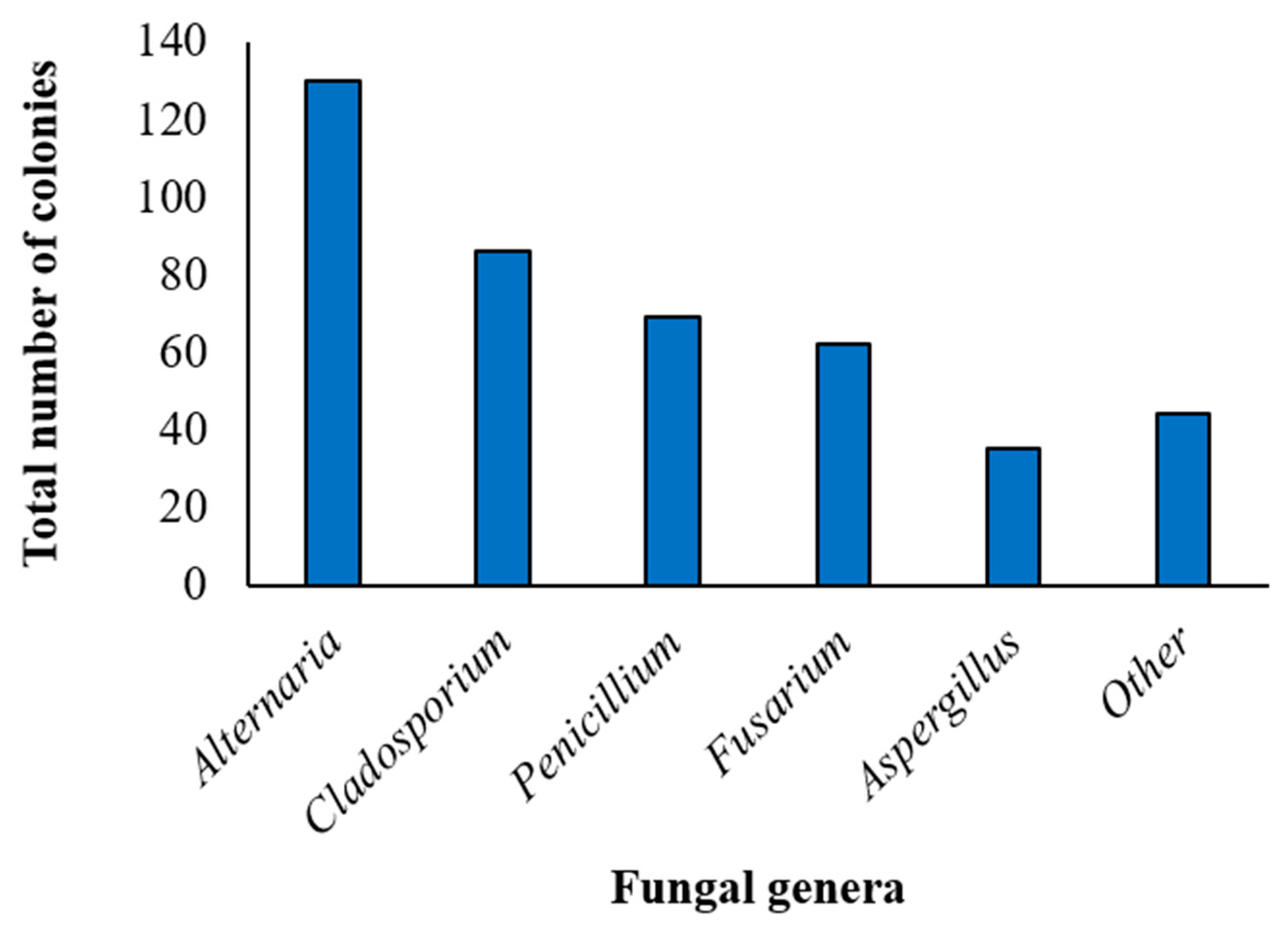
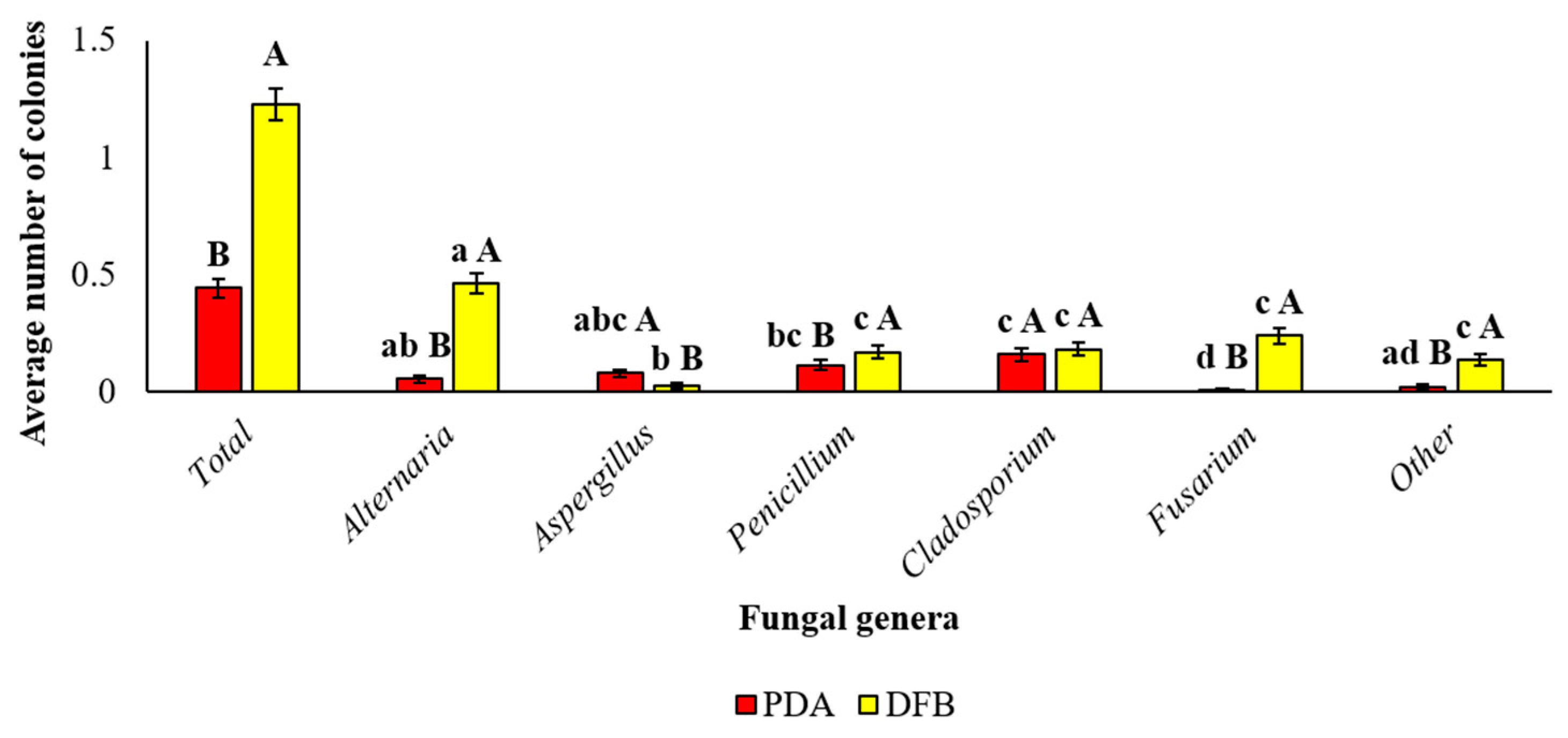
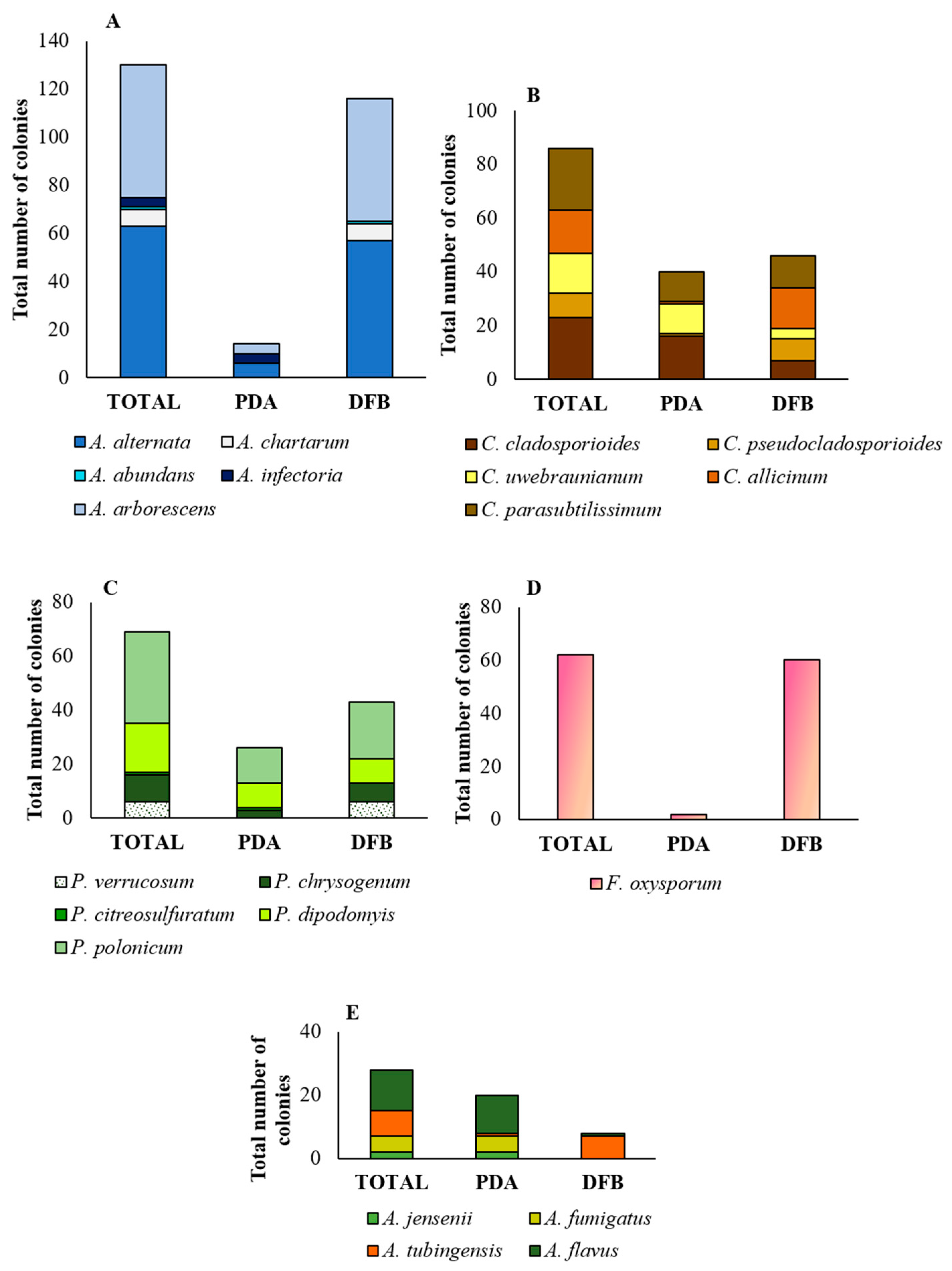
3.1. Mycobiota Associated with Marketed Quinoa Seed Samples
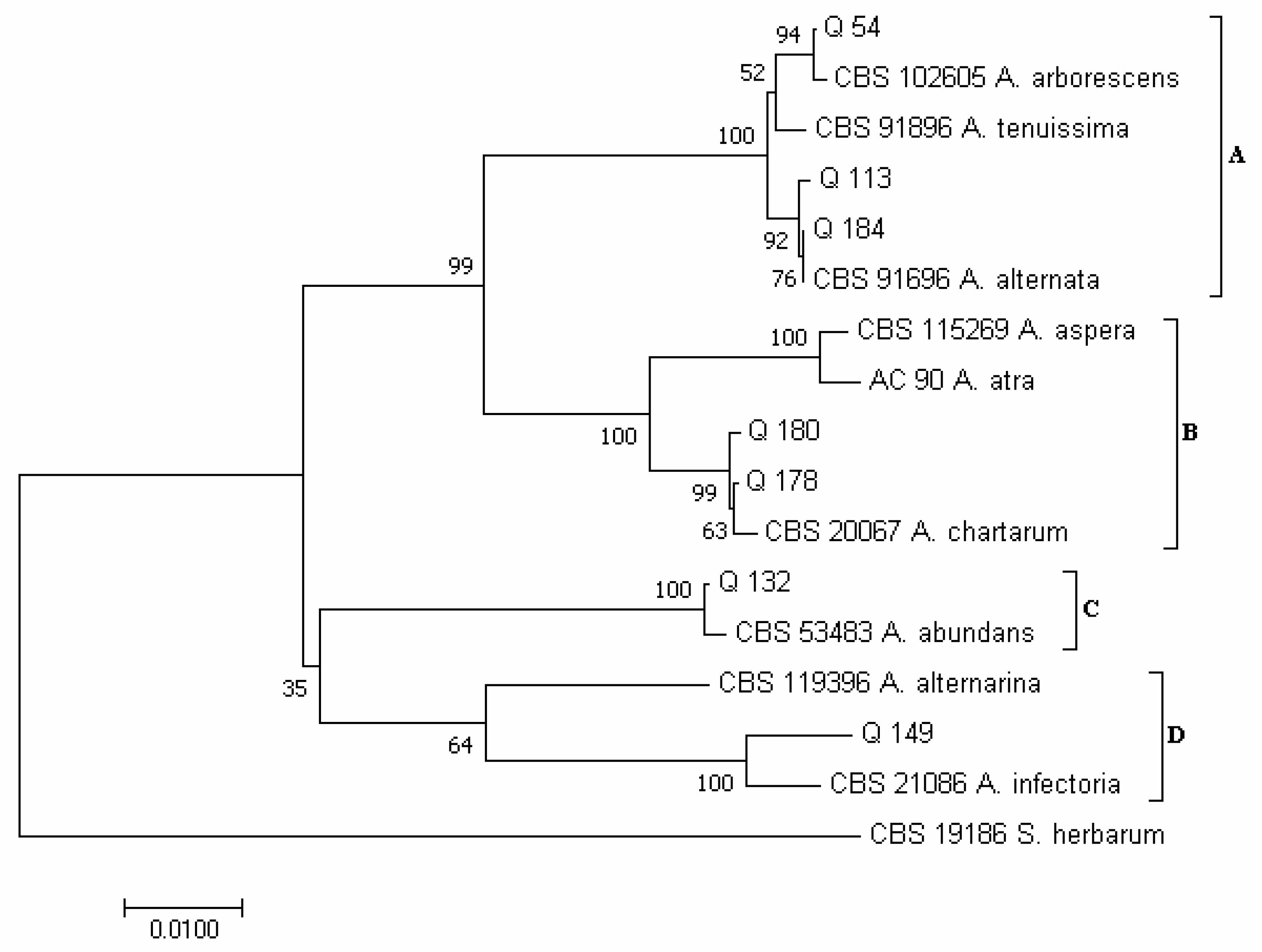
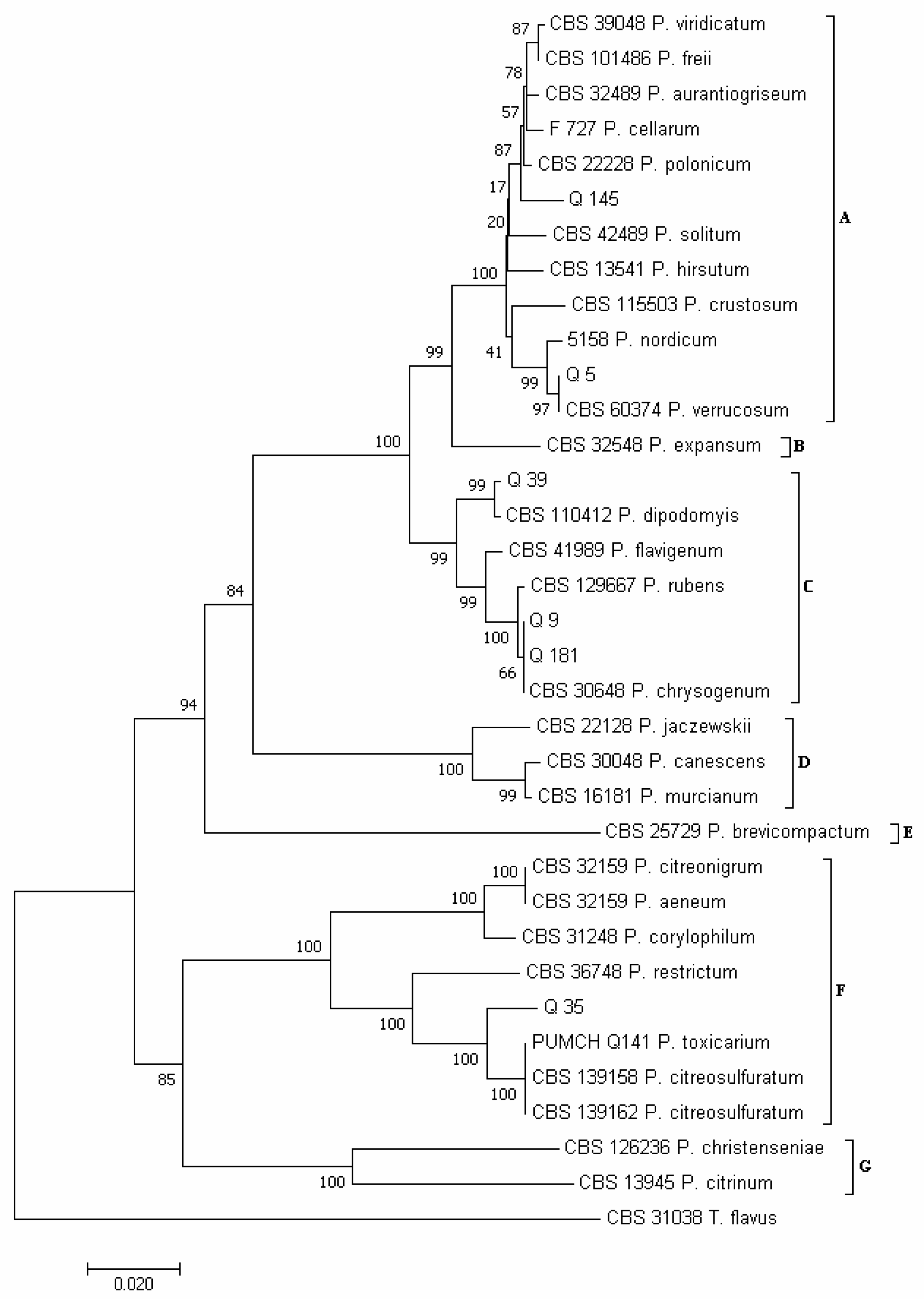
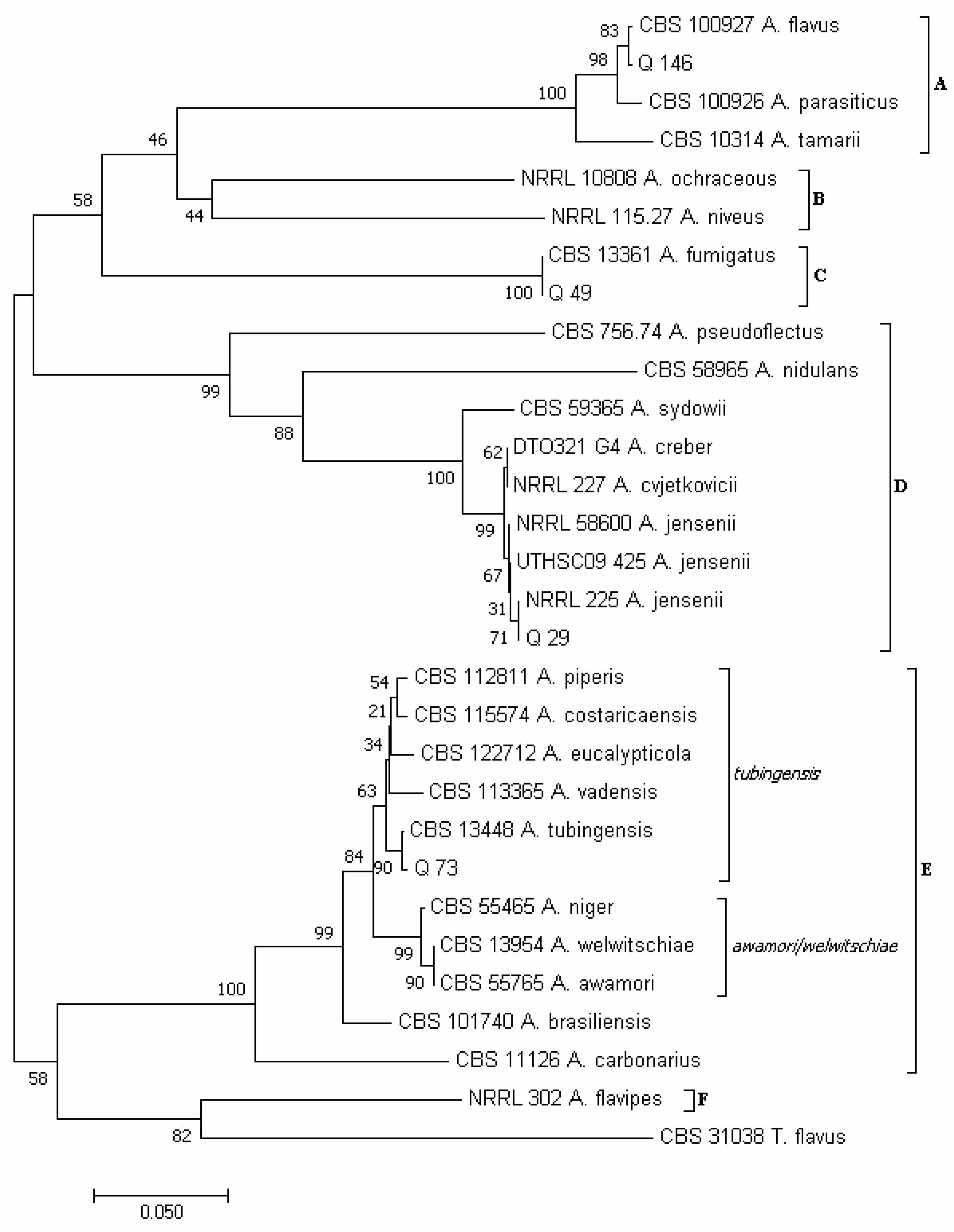
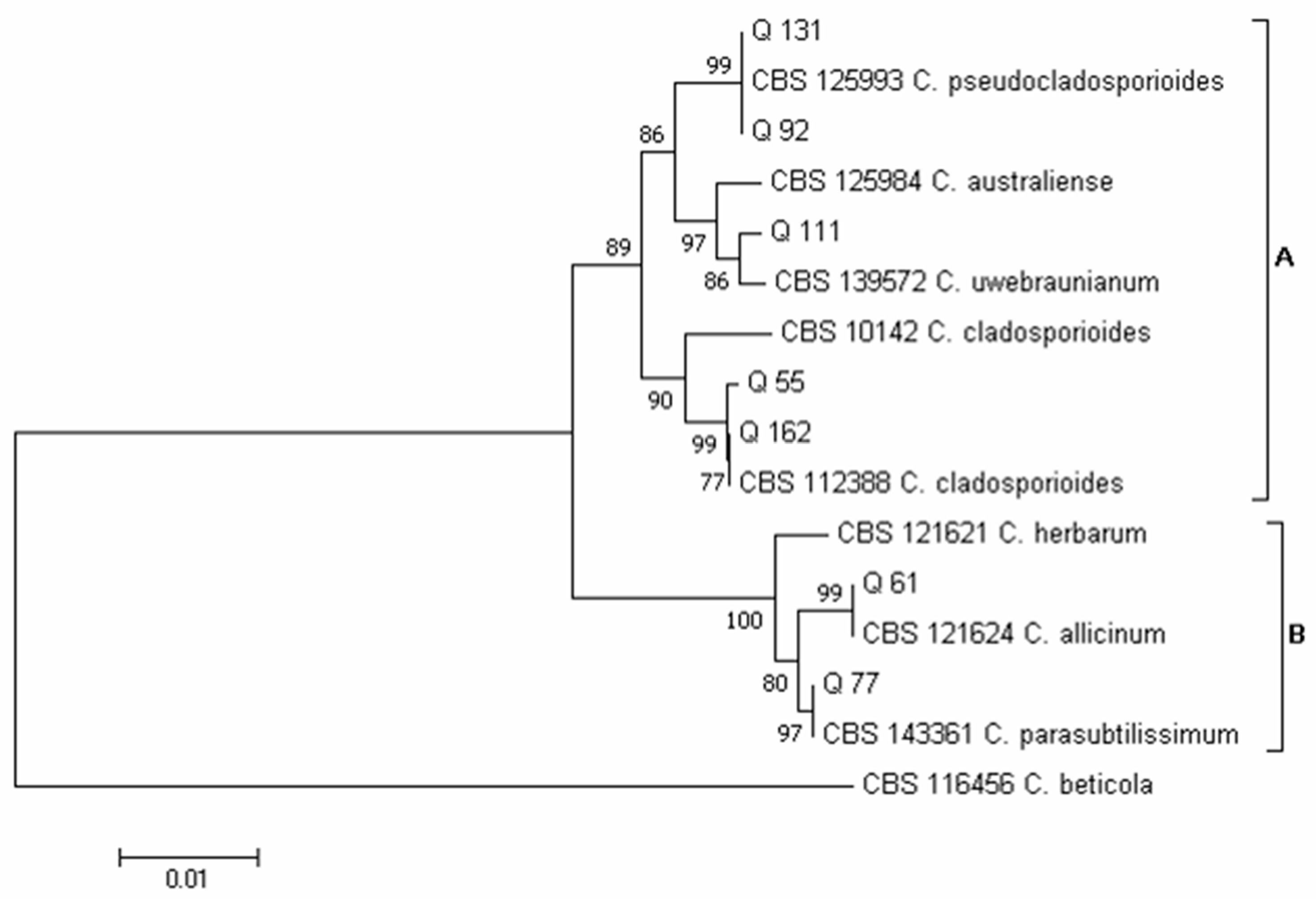
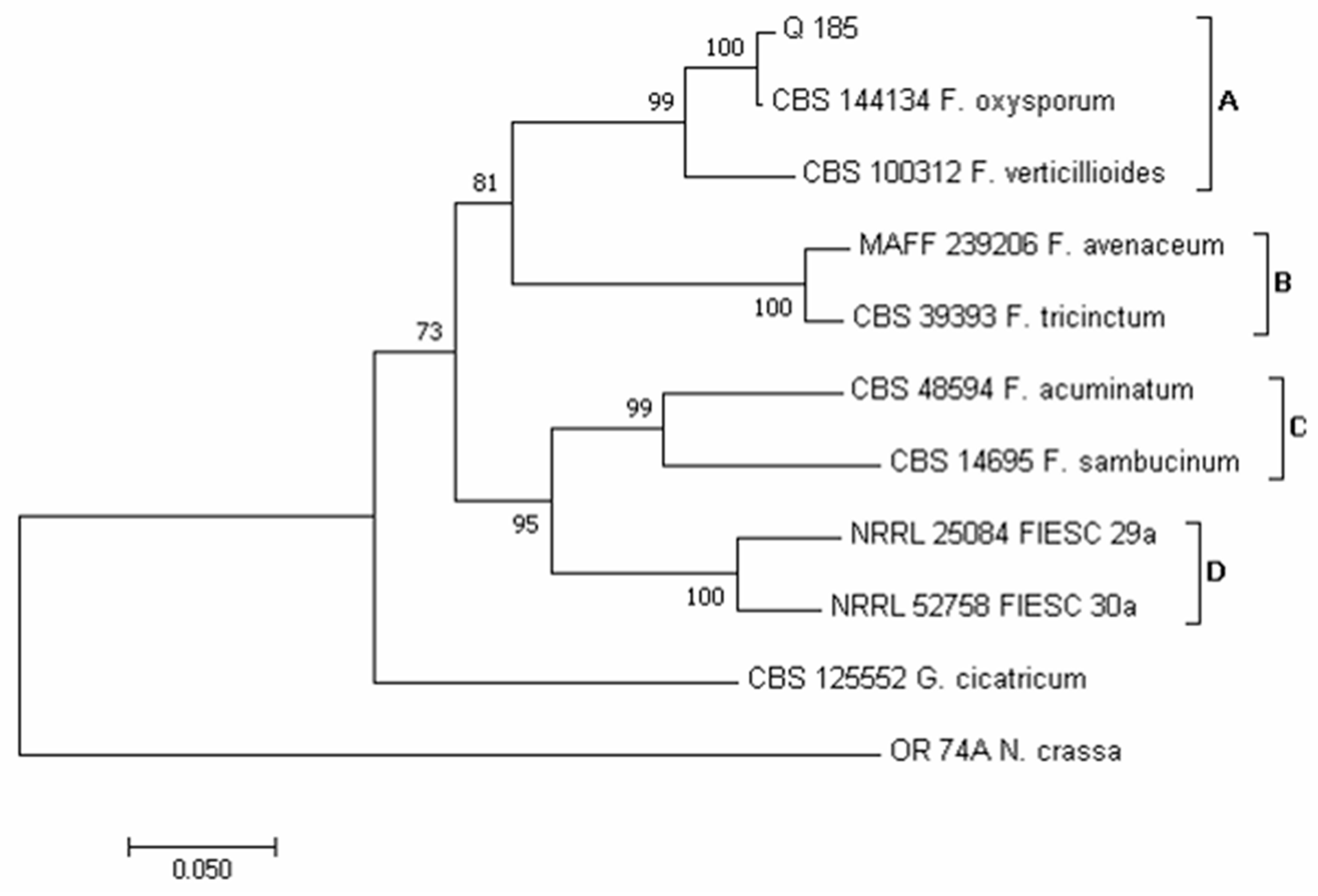
3.2. Phylogenetic Analysis of Isolates Developed from Marketed Quinoa Seeds
3.3. Secondary Metabolites in the Marketed Quinoa Seed Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, J.; Zhou, X.; Huang, H.; Li, G. Diseases characteristic and control measurements for Chenopodium quinoa Willd. Adv. Eng. Res. 2017, 143, 305–308. [Google Scholar]
- Hussain, M.I.; Farog, M.; Syed, Q.A.; Ishaq, A.; Al-Ghamdi, A.A.; Hatamleh, A.A. Botany, nutritional value, phytochemical composition and biological activities of quinoa. Plants 2021, 10, 2258. [Google Scholar] [CrossRef]
- Wilson, H.; Heiser, C.B. The origin and evolutionary relationship of huauzonthe (Chenopodium nuttalliae) domesticated chenopod of Mexico. Am. J. Bot. 1979, 66, 198–206. [Google Scholar] [CrossRef]
- Angeli, V.; Silva, P.M.; Massuela, D.C.; Khan, M.W.; Hamar, A.; Khajehei, F.; Graeff-Hönninger, S.; Piatti, C. Quinoa (Chenopodium quinoa Willd.): An overview of the potentials of the “golden grain” and socio-economic and environmental aspects of its cultivation and marketization. Foods 2020, 9, 216. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, S.E. The Worldwide Potential for quinoa (Chenopodium quinoa Willd.). Food Rev. Int. 2006, 19, 167–177. [Google Scholar] [CrossRef]
- Gordillo-Bastidas, E.; Díaz-Rizzolo, D.A.; Roura, E.; Massanés, T.; Gomis, R. Quinoa (Chenopodium quinoa Willd.), from nutritional value to a potential health benefits; an integrative review. J. Nutr. Food Sci. 2016, 6, 497. [Google Scholar]
- Bazile, D.; Jacobsen, S.E.; Verniau, A. The global expansion of Quinoa: Trends and limits. Front. Plant Sci. 2016, 7, 622. [Google Scholar] [CrossRef] [Green Version]
- Lavini, A.; Pulvento, C.; d’Andria, R.; Riccardi, M.; Choukr-Allah, R.; Balhabib, O.; Yazar, A.; Incekaya, C.; Metin Sezen, S.; Qadir, M.; et al. Quinoa’s potential in the mediterranean region. J. Agron. Crop Sci. 2014, 5, 344–360. [Google Scholar] [CrossRef]
- Pulvento, C.; Riccardi, M.; Biondi, S.; Orsini, F.; Jacobsen, S.E.; Ragab, R.; d’Andria, R.; Lavini, A. Quinoa in Italy: Research and perspectives. In State of the Art Report on Quinoa in the World in 2013, 1st ed.; Bazile, D., Bertero, D., Nieto, C., Eds.; Food and Agriculture Organisation (FAO)—UN: Rome, Italy, 2015; pp. 454–465. [Google Scholar]
- Bilalis, D.J.; Roussis, I.; Kakabouki, I.; Folina, A. Quinoa (Chenopodium quinoa Willd.) crop under mediterranean conditions: A review. Int. J. Agric. Nat. Resour. 2019, 46, 51–68. [Google Scholar] [CrossRef]
- Damiani, F.; Pannacci, E.; Brunetti, M. Quinoa cultivation in Italy. In Quinoa Cultivation, Nutritional Properties and Effects on Health, 1st ed.; Gai, F., Peiretti, P.G., Eds.; Nova Science Publisher Inc.: Hauppauge, NY, USA, 2019. [Google Scholar]
- Food and Agriculture Organisation (FAO). Catalogue of Commercial Varieties of Quinoa in Peru; Food and Agriculture Organisation (FAO): Lima, Peru, 2015; Available online: http://www.fao.org/documents/card/en/c/f351e07b-bd68-4724-a849-778dd44e6 (accessed on 15 May 2021).
- van der Kamp, J.W.; Poutanen, K.; Seal, C.J.; Richardson, D.P. The HEALTGRAIN definition of ‘whole grain’. Food Nutr. Res. 2014, 4, 58. [Google Scholar]
- Ross, A.B.; van der Kamp, J.W.; King, R.; Lê, K.A.; Mejborn, H.; Seal, C.J.; Thielecke, F.; on behalf of the Healthgrain Forum. Perspective: A definition for whole-grain food products-recommendations from the healthgrain forum. Adv. Nutr. 2017, 8, 525–531. [Google Scholar] [CrossRef] [Green Version]
- Ruggiero, E.; Bonaccio, M.; Di Castelnuovo, A.; Bonanni, A.; Costanzo, S.; Persichillo, M.; Bracone, F.; Cerletti, C.; Donati, M.B.; de Gaetano, G.; et al. Consumption of whole grain food and its determinants in a general Italian population: Results from the INHES study. Nutr. Metab. Cardiovasc. Dis. 2019, 6, 611–620. [Google Scholar] [CrossRef]
- Milićević, D.R.; Škrinjar, M.; Baltić, T. Real and perceived risks for mycotoxin contamination in foods and feeds: Challenges for food safety control. Toxins 2010, 2, 572–592. [Google Scholar] [CrossRef] [Green Version]
- Abrunhosa, L.; Morales, H.; Soares, C.; Calado, T.; Vila-Chã, A.S.; Pereira, M.; Venâncio, A. A review of mycotoxins in food and feed products in Portugal and estimation of probable daily intakes. Crit. Rev. Food Sci. Nutr. 2016, 56, 249–265. [Google Scholar] [CrossRef] [Green Version]
- Pappier, U.; Fernández, V.; Larumbe, G.; Vaamonde, G. Effect of processing for saponin removal of fungal contamination of quinoa seeds (Chenopodium quinoa Willd.). Int. J. Food Microbiol. 2008, 125, 153–157. [Google Scholar] [CrossRef]
- Guarniz-Benites, J.; Valdez-Arana, J.D.C. Morphological identification of mycotoxigenic fungi in accessions of quinoa (Chenopodium quinoa Wild.) of the Peruvian coast and sierra. Sci. Agropecu. 2019, 10, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Herrera, M.; Bervis, N.; Carramiñana, J.J.; Juan, T.; Herrera, A.; Ariño, A.; Lorán, S. Occurrence and exposure assessment of aflatoxins and deoxynivalenol in cereal-based baby food for infants. Toxins 2019, 11, 150. [Google Scholar] [CrossRef] [Green Version]
- Sacco, C.; Donato, R.; Zanella, B.; Pini, G.; Pettini, L.; Marino, M.F.; Rookmin, A.D.; Marvasi, M. Mycotoxins and flours: Effect of type crop, organic production, packaging type on the recovery of fungal genus and mycotoxins. Int. J. Food Microbiol. 2020, 334, 108808. [Google Scholar] [CrossRef]
- Ducos, C.; Pinson-Gadais, L.; Chereau, S.; Richard-Forget, F.; Vásquez-Ocmín, P.; Cerapio, J.P.; Casavilca-Zambrano, S.; Ruiz, E.; Pineau, P.; Bertani, S. Natural occurrence of mycotoxin-producing Fusaria market-bought Peruvian cereals: A food safety threat for Andean populations. Toxins 2021, 13, 172. [Google Scholar] [CrossRef]
- Ramos-Diaz, J.M.; Sulyok, M.; Jacobsen, S.E.; Jouppila, K.; Nathanail, A.V. Comparative study of mycotoxin occurrence in Andean and cereal grains cultivated in South America and North Europe. Food Control 2021, 130, 108260. [Google Scholar] [CrossRef]
- Limonard, T.A. Modified blotter test for seed health. Neth. J. Plant Pathol. 1966, 72, 319. [Google Scholar] [CrossRef]
- Covarelli, L.; Beccari, G.; Prodi, A.; Generotti, S.; Etruschi, F.; Juan, C.; Ferrer, E.; Manes, J. Fusarium species, chemotype characterization and trichotecene contamination of durum and soft wheat in an area of central Italy. J. Sci. Food Agric. 2015, 95, 540–551. [Google Scholar] [CrossRef]
- Beccari, G.; Caproni, L.; Tini, F.; Uhlig, S.; Covarelli, L. Presence of Fusarium species and other toxigenic fungi in malting barley and multi-mycotoxin analysis by Liquid Chromatography-High-Resolution mass spectrometry. J. Agric. Food Chem. 2016, 64, 4390–4399. [Google Scholar] [CrossRef]
- Quaglia, M.; Santinelli, M.; Sulyok, M.; Onofri, A.; Covarelli, L.; Beccari, G. Aspergillus, Penicillium and Cladosporium species associated with dried date fruits collected in the Perugia (Umbria, Central Italy) market. Int. J. Food Microbiol. 2020, 322, 108585. [Google Scholar] [CrossRef]
- Castegnaro, M.; Wild, C.P. IARC activities in mycotoxins research. Nat. Toxins 1995, 3, 327–331. [Google Scholar] [CrossRef]
- Perrone, G.; Mulè, G.; Susca, A.; Battilani, P.; Pietri, A.; Logrieco, A. Ochratoxin A production and amplified fragment length polymorphism analysis of Aspergillus carbonarius, Aspergillus tubingensis, and Aspergillus niger strains isolated from grapes in Italy. Appl. Environ. Microbiol. 2006, 72, 680–685. [Google Scholar] [CrossRef] [Green Version]
- Houbraken, J.; Samson, R.A. Phylogeny of Penicillium and segregation of Trichocomaceae into three families. Stud. Mycol. 2011, 70, 1–51. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef] [Green Version]
- Wondenberg, J.H.C.; Seidi, M.F.; Groenewald, J.Z.; de Vries, M.; Stielow, J.B.; Thomma, B.P.H.J.; Crous, P.W. Alternaria section Alternaria: Species, formae speciales or pathotypes? Stud. Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Bensch, K.; Groenewald, J.Z.; Dijksterhuis, J.; Starink-Willemse, M.; Andersen, B.; Summerell, B.A.; Shin, H.D.; Dugan, F.M.; Schroers, H.J.; Braun, U.; et al. Species and ecological diversity within the Cladosporium cladosporioides complex (Davidiellaceae, Capnodiales). Stud. Mycol. 2010, 67, 1–94. [Google Scholar] [CrossRef]
- Varga, J.; Frisvad, J.C.; Kocsubé, S.; Brankovics, B.; Tóth, B.; Samson, R.A. New and revised species in Aspergillus section Nigri. Stud. Mycol. 2011, 69, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frisvad, J.C.; Hubka, V.; Ezekiel, C.N.; Hong, S.B.; Nováková, A.; Chen, A.J.; Larsen, T.O.; Sklenár, F.; Mahakarnchanakul, W.; Samson, R.A.; et al. Taxonomy of Aspergillus section Flavi and their production of aflatoxins, ochratoxins and other mycotoxins. Stud. Mycol. 2019, 93, 1–63. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Ward, T.J.; Abeffa, D.; Kistler, H.C.; Aoki, T.; Orwig, N.; Kimura, M.; Bjørnstad, S.; Klemsdal, S.S. Multilocus genotyping and molecular phylogenetics resolve a novel head blight pathogen within the Fusarium graminearum species complex from Ethiopia. Fungal Genet. Biol. 2008, 45, 1514–1522. [Google Scholar] [CrossRef]
- Geiser, D.M.; Jimenes-Gasco, M.; Kang, S.; Makalowska, I.; Veeraraghavan, N.; Ward, T.J.; Zhang, N.; Khuldau, G.A.; O’Donnell, K. FUSARIUM-ID v.1.0: A DNA sequence database for identifying Fusarium. Eur. J. Plant Pathol. 2004, 110, 473–479. [Google Scholar] [CrossRef]
- Watanabe, M.; Yonezawa, T.; Lee, K.I.; Kumagai, S.; Sugita-Konishi, Y.; Goto, K.; Hara-Kudo, Y. Molecular phylogeny of the higher and lower taxonomy of the Fusarium genus and differences in the evolutionary histories of multiple genes. BMC Evol. Biol. 2011, 11, 322. [Google Scholar] [CrossRef] [Green Version]
- White, T.M.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal DNA for phylogenetics. In PCR Protocols: A Guide to Methods and Applications, 1st ed.; Inns, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–321. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Whelen, S.; Hall, B. Phylogenetic relationship among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.B.; Cho, H.S.; Shin, H.D.; Frisvad, J.C.; Samson, R.A. Novel Neosartorya species isolated from soil in Korea. Int. J. Syst. Evol. Microbiol. 2006, 56, 477–486. [Google Scholar] [CrossRef] [Green Version]
- Varga, J.; Due, M.; Frisvad, J.C.; Samson, R.A. Taxonomic revision of Aspergillus section Clavati based on molecular, morphological and physiological data. Stud. Mycol. 2007, 59, 89–106. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for bigger dataset. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Centre for Biotechnology Information (NCBI). Basic Local Alignment Search Tool (BLAST). Available online: http://blast.ncbi.nlm.nih.gov (accessed on 1 November 2021).
- González-Teuber, M.; Vilo, C.; Bascuñán-Godoy, L. Molecular characterization of endophytic fungi associated with the roots of Chenopodium quinoa inhabiting the Atacama Desert, Chile. Genom. Data 2017, 11, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Dřímalková, M.; Veverka, K. Seedlings damping-off of Chenopodium quinoa Willd. Plant Prot. Sci. 2018, 40, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Colque-Little, C.; Amby, D.B.; Andreasen, C. A review of Chenopodium quinoa (Willd.) diseases—An updated perspective. Plants 2021, 10, 1228. [Google Scholar] [CrossRef]
- Beccari, G.; Quaglia, M.; Tini, F.; Pannacci, E.; Covarelli, L. Phytopatological treats associated with Quinoa (Chenopodium quinoa Willd.) cultivation and seed production in an area of Central Italy. Plants 2021, 10, 1993. [Google Scholar] [CrossRef]
- Yin, H.; Zhou, J.B.; Chen, Y.L.; Ren, L.; Qin, N.; Xing, Y.L.; Zhao, X.J. Morphology, phylogeny, and pathogenicity of Trichitecium, Alternaria, and Fusarium species associated with panicle rot on Chenopodium quinoa in Shanxi Province, China. Plant Pathol. 2021, 71, 344–360. [Google Scholar] [CrossRef]
- Fonseca-Guerra, I.R.; Chiquillo-Pompeyo, J.C.; Benavides Rozo, M.E.; Díaz Ovalle, J.F. Fusarium spp. associated with Chenopodium quinoa crops in Colombia. Sci. Rep. 2022, 12, 20841. [Google Scholar] [CrossRef]
- Li, X.; Yu, T.; Jiang, X.; Zhang, C.; Zhao, X.; Wang, J. First report of ear rot of Chenopodium quinoa caused by Cladosporium cladosporioides in Shanxi, China. Plant Dis. 2022, 106, 1532. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Pest categorization of Fusarium brachygibbosum. EFSA J. 2021, 19, 6887. [Google Scholar]
- Bensch, K.; Braun, U.; Groenewald, J.Z.; Crous, P.W. The genus Cladosporium. Stud. Mycol. 2012, 72, 1–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, M.; Yonezawa, T.; Lee, K.I.; Kumagai, S.; Sugita-Konishi, Y.; Goto, K.; Hara-Kudo, Y. Evaluation of genetic markers for identifying isolates of the species of the genus Fusarium. J. Sci. Food Agric. 2011, 91, 2500–2504. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Ryu, D.; Wolf-Hall, C. Yeasts and molds. In Compendium of Methods for the Microbiological Examination of Foods, 5th ed.; Salfinger, Y., Tortorello, M.L., Eds.; American Public Health Association: Washington, DC, USA, 2001. [Google Scholar]
- Sulyok, M.; Stadler, D.; Steiner, D.; Krska, R. Validation of an LC-MS/ MS-based dilute-and-shoot approach for the quantification of >500 mycotoxins and other secondary metabolites in food crops: Challenges and solutions. Anal. Bioanal. Chem. 2020, 412, 2607–2620. [Google Scholar] [CrossRef] [Green Version]
- Weisberg, S. Applied Linear Regression, 3rd ed.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2005. [Google Scholar]
- Bretz, F.; Hothorn, T.; Westfall, P. Multiple Comparisons Using R; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 25 May 2021).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R.V. Emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.7.1-9990006. Available online: https://github.com/rvlenth/emmeans (accessed on 25 May 2022).
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef] [Green Version]
- Kozlovskii, A.G.; Zhelifonova, V.P.; Antipova, T.V. Fungi of the genus Penicillium as producers of physiologically active compounds (Review). Appl. Biochem. Microbiol. 2013, 49, 1–10. [Google Scholar] [CrossRef]
- Bräse, S.; Encinas, A.; Keck, J.; Nising, C.F. Chemistry and biology of mycotoxins and related fungal metabolites. Chem. Rev. 2009, 109, 3903–3990. [Google Scholar] [CrossRef]
- Visagie, C.M.; Seifert, K.A.; Houbraken, J.; Samson, R.A.; Jacobs, K. A phylogenetic revision of Penicillium sect. Exilicaulis, including nine new species from fynbos in South America. IMA Fungus 2016, 7, 75–117. [Google Scholar]
- Houbraken, J.; Kocsubé, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Lombard, L.; Sandoval-Denis, M.; Lamprecht, S.C.; Crous, P.W. Epitypification of Fusarium oxysporum—Clearing the taxonomic chaos. Persoonia 2019, 4, 1–47. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.A.; Schroers, H.J.; Chaverri, P.; Gené, J.; Guarro, J.; Hirooka, Y.; Bensch, K.; et al. Fusarium: More than a node or a foot-shaped basal cell. Stud. Mycol. 2021, 98, 100116. [Google Scholar] [CrossRef]
- Laraba, I.; McCormick, S.P.; Vaughan, M.M.; Geiser, D.M.; O’Donnell, K. Phylogenetic diversity, trichothecene potential, and pathogenicity within Fusarium sambucinum species complex. PLoS ONE 2021, 16, e0245037. [Google Scholar] [CrossRef]
- Castellá, G.; Cabañes, F.J. Phylogenetic diversity of Fusarium incarnatum-equiseti species complex isolated from Spanish wheat. Antoine Van Leeuwenhoek 2014, 106, 309–317. [Google Scholar] [CrossRef]
- Drimalkova, M. Mycoflora of Chenopodium quinoa seeds. Plant Prot. Sci. 2003, 4, 146–150. [Google Scholar]
- Spehar, R.C.; Mendes, M.A.S.; Bhering Nasser, L.C. Analise Micologica de Sementes de Quinoa (Chenopodium quinoa Willd.) Selecionada o Brasil Central; Relatorio Tecnico Annual do Centro de Pesquisa Agropecuaria dos Cerrados Planaltina; EMBRAPA: Brasilia, Brazil, 1997; pp. 213–214. [Google Scholar]
- Al-Sadi, A.M.; Al-Alawi, Z.A.; Patzelt, A. Association of Alternaria alternata and Cladosporium cladosporioides with leaf spot in Cissu quadrangularis and Ficus cycomorus. Plant Pathol. J. 2015, 14, 44–47. [Google Scholar] [CrossRef] [Green Version]
- Commission Regulation (EC). Commission Regulation (EC) No 1126/2007 of 8 September amending Regulation (EC) No 1881/2006 setting maximum levels for certain contaminants in foodstuffs as regards Fusarium toxins in maize and maize products. Off. J. Eur. Union 2007, L225, 14–17. [Google Scholar]
- Escrivá, L.; Oueslati, S.; Font, G.; Manyes, L. Alternaria mycotoxins in food and feed: An overview. J. Food Qual. 2017, 2017, 1569748. [Google Scholar] [CrossRef] [Green Version]
- Hellwig, V.; Grothe, T.; Mayer-Bartschmid, A.; Endermann, R.; Geschke, F.U.; Henkel, T.; Stadler, M. Altersetin, a new antibiotic from cultures of endophytic Alternaria spp. taxonomy, fermentation, isolation, structure elucidation and biological activities. J. Antibiot. 2002, 55, 881–892. [Google Scholar] [CrossRef] [Green Version]
- Arcella, D.; Eskola, M.; Gómez Ruiz, J.A. Dietary exposure assessment to Alternaria toxins in the European population. EFSA J. 2016, 14, 4654. [Google Scholar]
- Palmieri, A.; Petrini, M. Tryptophol and derivatives; natural occurrence and applications to the synthesis of bioactive compounds. Nat. Prod. Rep. 2019, 36, 490–530. [Google Scholar] [CrossRef]
- Pathak, N.; Zaidi, R.K. Studies on seed-borne fungi of wheat in seed health testing programme. Arch. Phytopathol. 2013, 46, 389–401. [Google Scholar] [CrossRef]
- Beccari, G.; Prodi, A.; Senatore, M.T.; Balmas, V.; Tini, F.; Onofri, A.; Pedini, L.; Sulyok, M.; Brocca, L.; Covarelli, L. Cultivation area affects the presence of fungal communities and secondary metabolites in Italian durum wheat grains. Toxins 2020, 12, 97. [Google Scholar] [CrossRef] [Green Version]
- González, N.; Marquès, M.; Nadal, M.; Domingo, J.L. Occurrence of environmental pollutants in foodstuffs: A review of organic vs. conventional food. Food Chem. Toxicol. 2019, 125, 370–375. [Google Scholar] [CrossRef]
- Malmauret, L.; Parent-Massin, D.; Hardy, J.L.; Verger, P. Contaminants in organic and conventional foodstuffs in France. Food Addit. Contam. 2002, 19, 524–532. [Google Scholar] [CrossRef]
- Piemontese, L.; Solfrizzo, M.; Visconti, A. Occurrence of patulin in conventional and organic fruit products in Italy and subsequent exposure assessment. Food Addit. Contam. 2005, 22, 437–442. [Google Scholar] [CrossRef]
- Opara, U.L.; Caleb, O.J.; Uchechukwu-Agua, A.D. Evaluating the impact of selected packaging materials on the quality attributes of cassava flour (cvs. TME 419 and UMUCASS 36). J. Food Sci. 2016, 81, 324–331. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Origin | Seed Color | Farming | Packaging |
|---|---|---|---|---|
| 1 | Extra EU (Peru) | White | Integrated | Sealed Plastic Box |
| 2 | EU (Italy) | White | Integrated | Sealed Plastic Box |
| 3 | EU (Italy) | White | Integrated | Sealed Plastic Box |
| 4 | Extra EU (Peru) | White | Organic | Sealed Plastic Box |
| 5 | Extra EU (Country not specified) | White | Integrated | Sealed Plastic Box |
| 6 | Extra EU (Peru) | White | Organic | Sealed Plastic Box |
| 7 | EU (Italy) | White | Integrated | Sealed Plastic Box |
| 8 | EU (Italy) | White | Integrated | Sealed Plastic Box |
| 9 | Extra EU (Country not specified) | White | Organic | Sealed Plastic Box |
| 10 | Extra EU (Country not specified) | White | Integrated | Unsealed Cardboard Box |
| 11 | Extra EU (Peru) | White | Integrated | Bulk |
| 12 | Extra EU (Country not specified) | White | Organic | Sealed Plastic Box |
| 13 | Extra EU (Peru) | White | Integrated | Sealed Plastic Box |
| 14 | Extra EU (Peru) | Black | Integrated | Sealed Plastic Box |
| 15 | Extra EU (Peru) | Red | Integrated | Sealed Plastic Box |
| 16 | Extra EU (Peru) | White | Integrated | Sealed Plastic Box |
| 17 | EU (Country not specified) | White | Organic | Sealed Plastic Box |
| 18 | Extra EU (Bolivia) | Black | Organic | Sealed Plastic Box |
| 19 | Extra EU (Bolivia) | White | Organic | Sealed Plastic Box |
| 20 | Extra EU (Peru) | White | Organic | Sealed Plastic Box |
| 21 | Extra EU (Bolivia) | White | Organic | Bulk |
| 22 | Extra EU (Chile) | White | Integrated | Bulk |
| 23 | Extra EU (Ecuador) | White | Integrated | Bulk |
| 24 | Extra EU (Ecuador) | White | Integrated | Bulk |
| 25 | Extra EU (Bolivia) | White | Integrated | Bulk |
| Sample | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | |
| Alternaria | ● * | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ||||||
| Cladosporium | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | |||
| Penicillium | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | |||||||||||
| Aspergillus | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ||||||||||||||
| Fusarium | ● | ● | ● | ● | |||||||||||||||||||||
| Penicillium Secondary Metabolites | μg Kg−1 |
|---|---|
| Anthraquinoids | |
| Endocrocin | 158 |
| Asperphenamates | |
| Asperphenamate | 377 |
| Diketopiperazines | |
| Brevianamide F | 5510 |
| Cyclo (L-Pro_L-Tyr) | 32,900 |
| Cyclo (L-Pro_L-Val) | 20,400 |
| Neoxaline | 1350 |
| Roquefortine C | 4870 |
| Roquefortine D | 146 |
| Puberulin A | 46,400 |
| Rugulosuvin | 17,900 |
| Imidazopyridoindoles | |
| Meleagrin | 1670 |
| Penicillic acids | |
| Penicillic acid | 404,000 |
| Propionic acids | |
| 3-Nitropropionic acid | 1,300,000 |
| Terpenes | |
| Andrastin A | 29,000 |
| Andrastin B | 1850 |
| Andrastin C | 45,900 |
| Mycophenolic acid | 4780 |
| Viridicatols | |
| Cyclopenin | 106,000 |
| Cyclopenol | 335,000 |
| Cyclopeptine | 27,500 |
| Dehydrocyclopeptine | 27,100 |
| O-Methylviridicatin | 36,800 |
| Viridicatin | 17,800 |
| Viridicatol | 428,000 |
| Secondary Metabolites | Seed Samples | ||||
|---|---|---|---|---|---|
| 21 | 22 | 23 | 24 | 25 | |
| Tetramic acid derivatives | |||||
| Altersetin | 30.6 | 1.18 | 13.1 | 4.96 | 8.56 |
| Diketopiperazines | |||||
| Brevianamide F | 3.53 | 0.25 | 1.92 | 3.55 | 2.25 |
| Cyclo (L-Pro L-Tyr) | 12.6 | 2.18 | 15.3 | 12.3 | 10.2 |
| Cyclo (L-Pro_L-Val) | 12.7 | 3.24 | 21.6 | 13.2 | 11.4 |
| Rugulosuvin | 1.04 | <LOD | <LOD | 1.06 | <LOD |
| Indole alcohol | |||||
| Tryptophol | 49.2 | 5.79 | 10.4 | 97.5 | 24.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quaglia, M.; Beccari, G.; Vella, G.F.; Filippucci, R.; Buldini, D.; Onofri, A.; Sulyok, M.; Covarelli, L. Marketed Quinoa (Chenopodium quinoa Willd.) Seeds: A Mycotoxin-Free Matrix Contaminated by Mycotoxigenic Fungi. Pathogens 2023, 12, 418. https://doi.org/10.3390/pathogens12030418
Quaglia M, Beccari G, Vella GF, Filippucci R, Buldini D, Onofri A, Sulyok M, Covarelli L. Marketed Quinoa (Chenopodium quinoa Willd.) Seeds: A Mycotoxin-Free Matrix Contaminated by Mycotoxigenic Fungi. Pathogens. 2023; 12(3):418. https://doi.org/10.3390/pathogens12030418
Chicago/Turabian StyleQuaglia, Mara, Giovanni Beccari, Giovanna Fabiana Vella, Riccardo Filippucci, Dario Buldini, Andrea Onofri, Michael Sulyok, and Lorenzo Covarelli. 2023. "Marketed Quinoa (Chenopodium quinoa Willd.) Seeds: A Mycotoxin-Free Matrix Contaminated by Mycotoxigenic Fungi" Pathogens 12, no. 3: 418. https://doi.org/10.3390/pathogens12030418
APA StyleQuaglia, M., Beccari, G., Vella, G. F., Filippucci, R., Buldini, D., Onofri, A., Sulyok, M., & Covarelli, L. (2023). Marketed Quinoa (Chenopodium quinoa Willd.) Seeds: A Mycotoxin-Free Matrix Contaminated by Mycotoxigenic Fungi. Pathogens, 12(3), 418. https://doi.org/10.3390/pathogens12030418