
Antimicrobial Potential of Betulinic Acid and Investigation of the Mechanism of Action against Nuclear and Metabolic Enzymes with Molecular Modeling

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
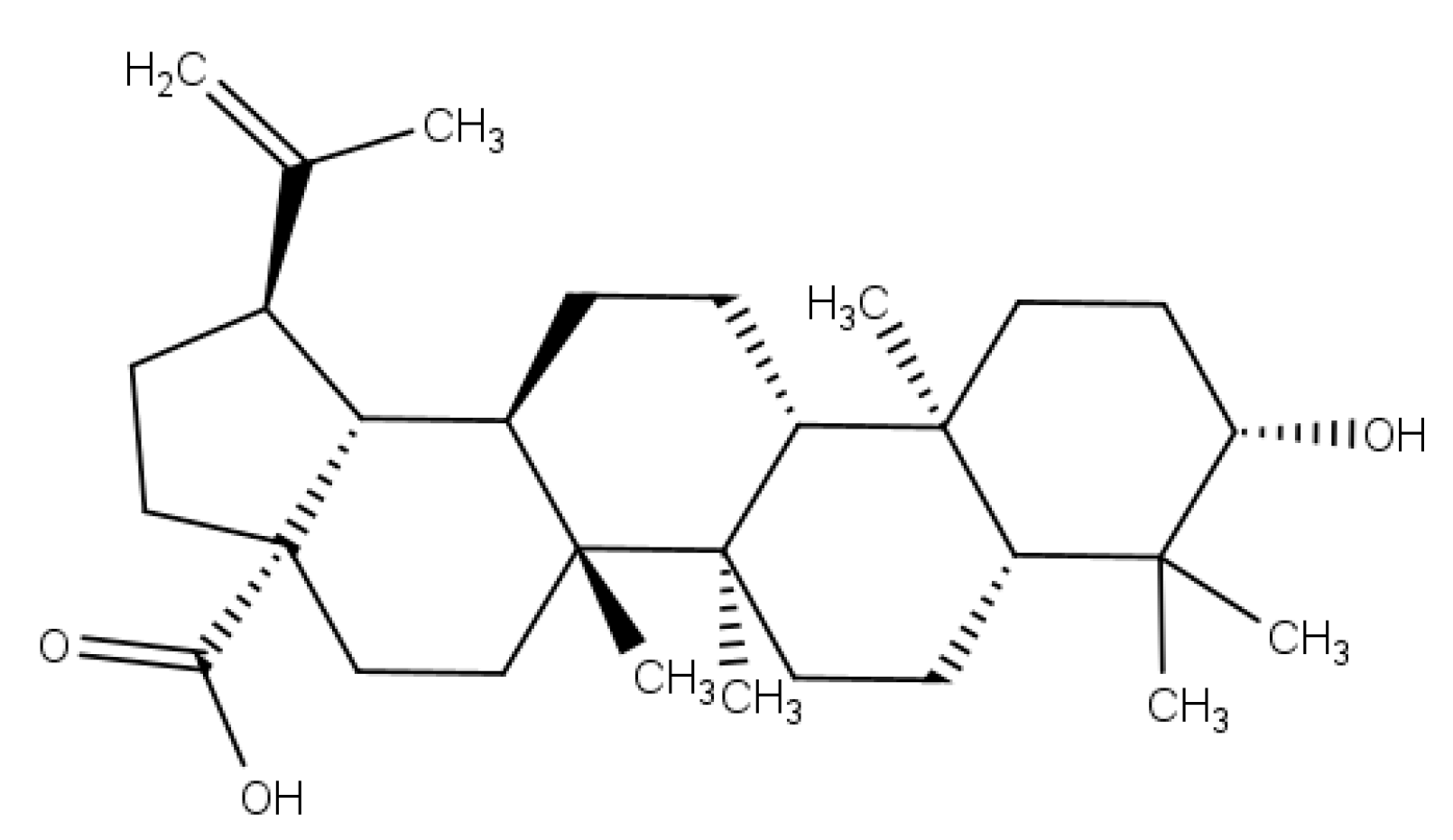
2.1. Extraction of Betulinic Acid
2.2. Quantitative Structure–Activity Relationship Modeling (QSAR)
2.3. Antimicrobial Activity Tests
2.3.1. Preparation of Compounds
2.3.2. Microorganisms
2.3.3. Minimum Inhibitory Concentration (MIC)
2.3.4. Minimum Bactericidal Concentration (MBC) and Minimum Fungicidal Concentration (MFC)
2.4. Molecular Modeling
2.4.1. Alignment of Protein Sequences
2.4.2. Modeling by Homology
2.4.3. Molecular Docking
3. Results
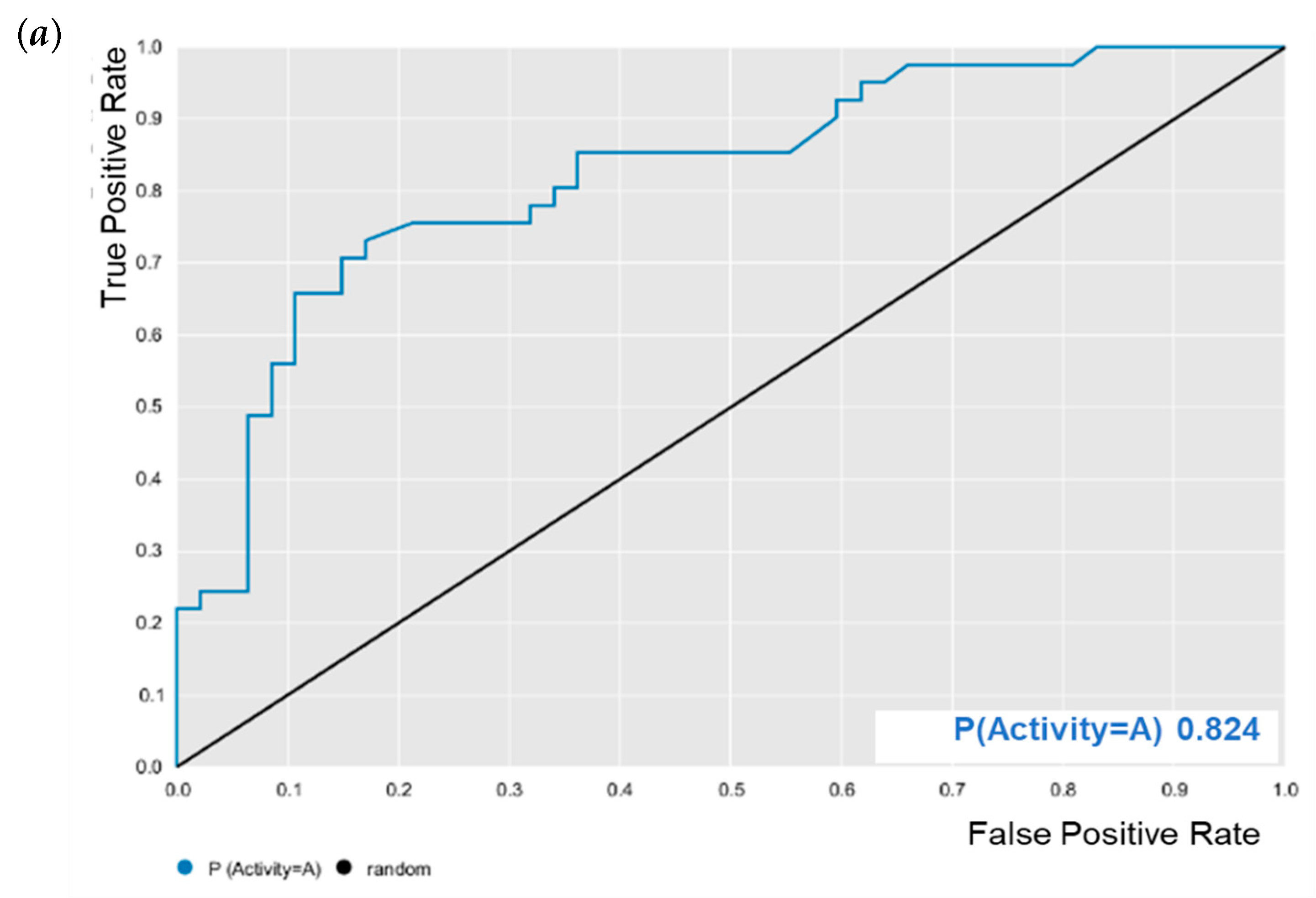
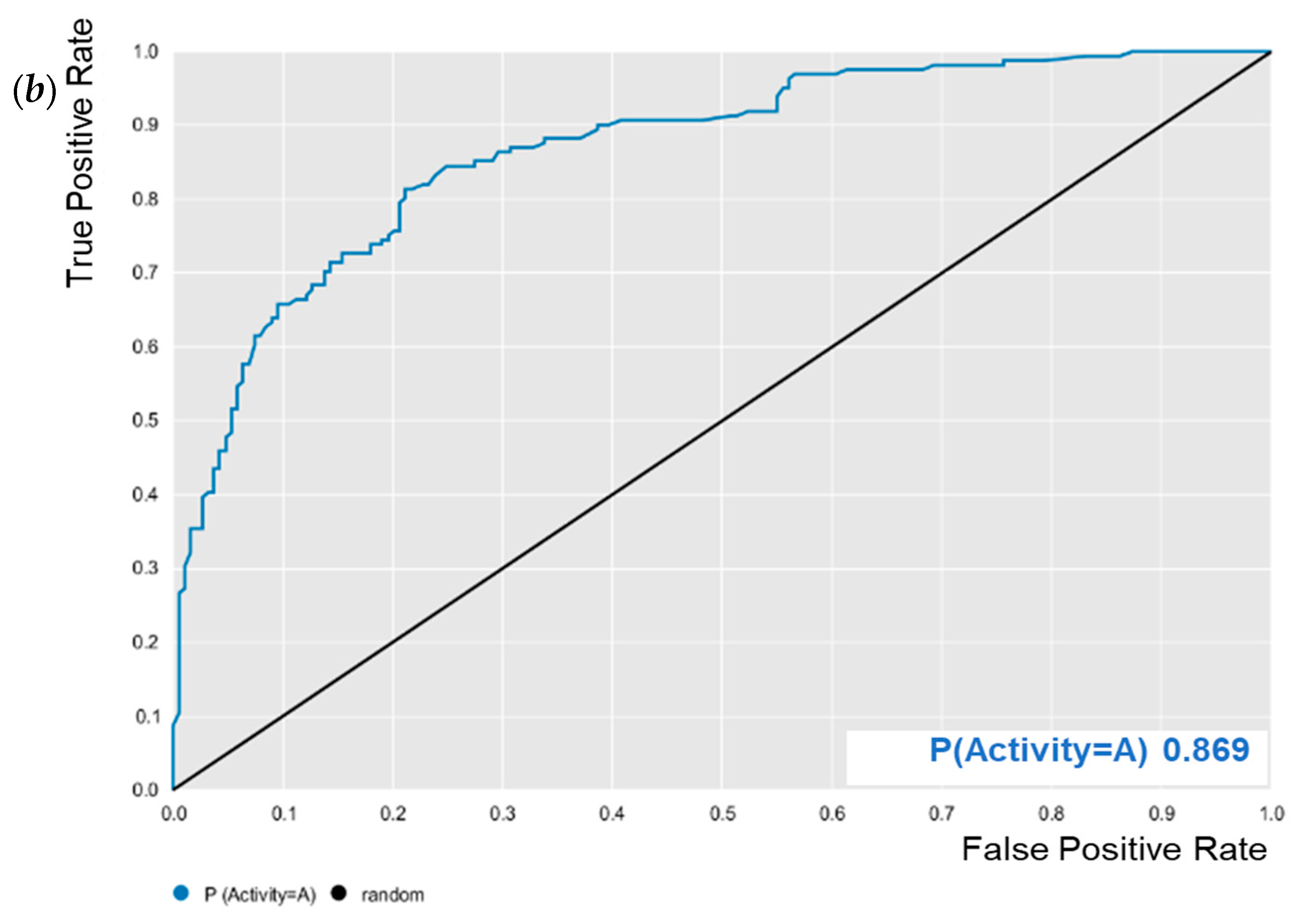
3.1. QSAR Modeling
3.2. Antimicrobial Activity Tests
3.3. Alignment of Protein Sequences
3.4. Modeling by Homology
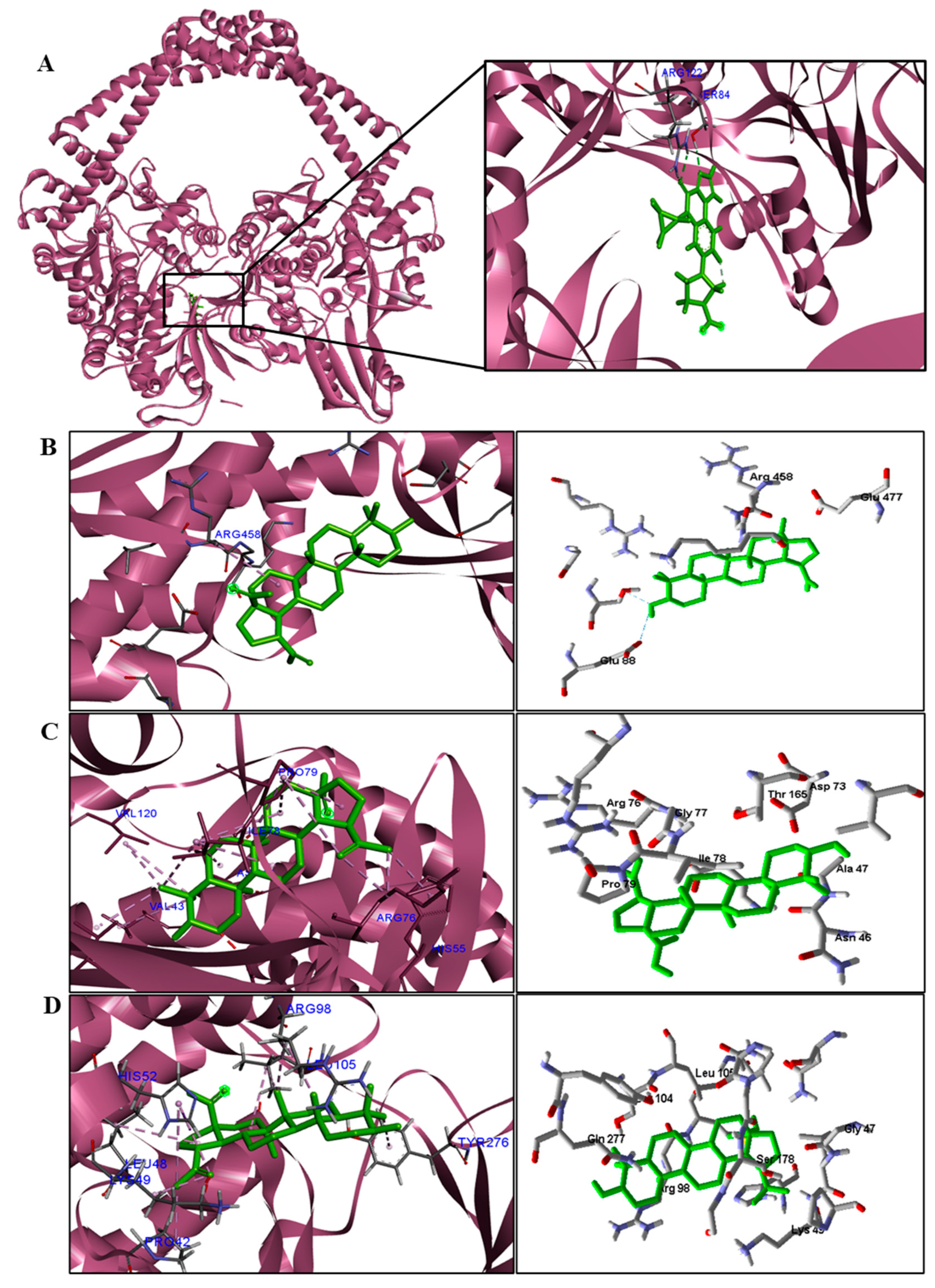
3.5. Molecular Docking
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, M.; Sarma, D.K.; Shubham, S.; Kumawat, M.; Verma, V.; Nina, P.B.; Jp, D.; Kumar, S.; Singh, B.; Tiwari, R.R. Futuristic Non−antibiotic Therapies to Combat Antibiotic Resistance: A Review. Front. Microbiol. 2021, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Abraham, E.P. The Antibiotics. Compr. Biochem. 1963, 11, 181–224. [Google Scholar]
- da Silva, G.N.S.; Primon−Barros, M.; Macedo, A.J.; Gnoatto, S.C.B. Triterpene derivatives as relevant scaffold for new antibiofilm drugs. Biomolecules 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Silver, L.L. Aropriate targets for antibacterial drugs. Cold Spring Harb. Perspect. Med. 2016, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Cárdenas, P.D.; Almeida, A.; Bak, S. Evolution of Structural Diversity of Triterpenoids. Front Plant Sci. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Jäger, S.; Trojan, H.; Ko, T.; Laszczyk, M.N.; Scheffler, A. Pentacyclic triterpene distribution in various plants—Rich sources for a new group of multi−potent plant extracts. Molecules 2009, 14, 2016–2031. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Kim, H.H.; Park, S.U.; Science, C. Recent Studies on Betulinic Acid and its Biological and Pharmacological Activity. EXCLI J. 2015, 14, 199–203. [Google Scholar] [PubMed]
- Sharon, N.; Lis, H. Biological activities. In Lectins; Published online; Springer: Berlin/Heidelberg, Germany, 1989; pp. 26–36. [Google Scholar] [CrossRef]
- Zhao, J.; Li, R.; Pawlak, A.; Henklewska, M.; Sysak, A.; Wen, L.; Yi, J.-E.; Obmińska−Mrukowicz, B. Antitumor activity of betulinic acid and betulin in canine cancer cell lines. In Vivo 2018, 32, 1081–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, J.F.O.; Barbosa−Filho, J.M.; de Azevedo Maia, G.L.; Guimarães, E.T.; Meira, C.S.; Ribeiro−dos−Santos, R.; de Carvalho, L.C.P.; Soares, M.B.P. Potent anti-inflammatory activity of betulinic acid treatment in a model of lethal endotoxemia. Int. Immunopharmacol. 2014, 23, 469–474. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Holmes, S.S.; Baker, G.A.; Challa, S.; Bose, H.S.; Song, Z. Ionic derivatives of betulinic acid as novel HIV−1 protease inhibitors. J. Enzyme Inhib. Med. Chem. 2012, 27, 715–721. [Google Scholar] [CrossRef] [Green Version]
- Song, T.-J.; Park, C.-H.; In, K.-R.; Kim, J.-B.; Kim, J.H.; Kim, M.; Chang, H.J. Antidiabetic effects of betulinic acid mediated by the activation of the AMP−activated protein kinase pathway. PLoS ONE 2021, 16, e0249109. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.A.; Elgindi, M.R.; Ibrahim, R.R.; El−Hosari, D.G. Antibacterial activities of triterpenoidal compounds isolated from Calothamnus quadrifidus leaves. BMC Complement. Altern. Med. 2019, 19, 19–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalcante, B.R.R.; Aragão−França, L.S.; Sampaio, G.L.A.; Nonaka, C.K.V.; Oliveira, M.S.; Campos, G.S.; Sardi, S.I.; Dias, B.R.S.; Menezes, J.P.B.; Rocha, V.P.C.; et al. Betulinic Acid Exerts Cytoprotective Activity on Zika Virus−Infected Neural Progenitor Cells. Front. Cell Infect. Microbiol. 2020, 10, 558324. [Google Scholar] [CrossRef]
- De Sá, M.S.; Costa, J.F.O.; Krettli, A.U.; Zalis, M.G.; Maia, G.L.D.A.; Sette, I.M.F.; Câmara, C.D.A.; Filho, J.M.B.; Giulietti−Harley, A.M.; Ribeiro dos Santos, R.; et al. Antimalarial activity of betulinic acid and derivatives in vitro against Plasmodium falciparum and in vivo in P. berghei−infected mice. Parasitol. Res. 2009, 105, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Yogeeswari, P.; Sriram, D. Betulinic Acid and Its Derivatives: A Review on their Biological Properties. Curr. Med. Chem. 2010, 12, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Stan, D.; Enciu, A.-M.; Mateescu, A.L.; Ion, A.C.; Brezeanu, A.C.; Tanase, C. Natural Compounds with Antimicrobial and Antiviral Effect and Nanocarriers Used for Their Transportation. Front. Pharmacol. 2021, 12, 1–25. [Google Scholar] [CrossRef]
- Aldholmi, M.; Marchand, P.; Ourliac−Garnier, I.; Le Pape, P.; Ganesan, A. A decade of antifungal leads from natural products: 2010–2019. Pharmaceuticals 2019, 12, 2010–2019. [Google Scholar] [CrossRef] [Green Version]
- Khameneh, B.; Iranshahy, M.; Soheili, V.; Sedigheh, B.; Bazzaz, F. Review on plant antimicrobials: A mechanistic viewpoint. Antimicrob. Resist. Infect. Control. 2019, 8, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Scorzoni, L.; de Paula, E.S.A.C.; Marcos, C.M.; Assato, P.A.; de Melo, W.C.; de Oliveira, H.C.; Costa−Orlandi, C.B.; Mendes−Giannini, M.J.; Fusco−Almeida, A.M. Antifungal therapy: New advances in the understanding and treatment of mycosis. Front. Microbiol. 2017, 8, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Hargrove, T.Y.; Friggeri, L.; Wawrzak, Z.; Qi, A.; Hoekstra, W.J.; Schotzinger, R.J.; York, J.D.; Guengerich, F.P.; Lepesheva, G.I. Structural analyses of Candida albicans sterol 14α−demethylase complexed with azole drugs address the molecular basis of azole−mediated inhibition of fungal sterol biosynthesis. J. Biol. Chem. 2017, 292, 6728–6743. [Google Scholar] [CrossRef] [Green Version]
- Guglielmi, P.; Pontecorvi, V.; Rotondi, G. Natural compounds and extracts as novel antimicrobial agents. Expert. Opin. Ther. Pat. 2020, 30, 949–962. [Google Scholar] [CrossRef] [PubMed]
- Matamoros−Recio, A.; Franco−Gonzalez, J.F.; Forgione, R.E.; Torres−Mozas, A.; Silipo, A.; Martín−Santamaría, S. Understanding the Antibacterial Resistance: Computational Explorations in Bacterial Membranes. ACS Omega 2021, 6, 6041–6054. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, J.M.; Cheriyan, U.O.; Pessoa, J. Constituents of the stem−bark of Zizyphus joazeiro. J. Nat. Prod. 1985, 48, 152–153. [Google Scholar]
- Aiken, C.; Chen, C.H. Betulinic acid derivatives as HIV-1 antivirals. Trends Mol. Med. 2005, 11, 31–36. [Google Scholar] [CrossRef]
- Cavalcanti, A.B.; de Figueiredo, P.T.; Veloso, C.A.; Rodrigues, G.C.; Maia, M.D.S.; Monteiro, A.F.M.; Junior, V.S.R.; Castelo−Branco, A.P.; Agra, M.D.F.; Raimundo Filho, B.; et al. A new labdane diterpene from the aerial segments of Leptohyptis macrostachys (L’Hérit.) Harley & J.F.B. Pastore. Phytochem Lett. 2021, 43, 117–122. [Google Scholar]
- Mauri, A.; Consonni, V.; Pavan, M.; Todeschini, R. DRAGON software: An easy aroach to molecular descriptor calculations. MATCH Commun. Math. Comput. Chem. 2006, 56, 237–248. [Google Scholar]
- Maia, S.M.; de Sousa, N.F.; Rodrigues, G.C.S.; Monteiro, A.F.M.; Scotti, M.T.; Scotti, L. Lignans and Neolignans Anti−tuberculosis Identified by QSAR and Molecular Modeling. Comb. Chem. High Throughput Screen. 2020, 23, 504–516. [Google Scholar] [CrossRef]
- Matthews, B.W. Comparison of the predicted and observed secondary structure of T4 phage lysozyme. Biochim. Biophys. Acta (BBA)—Protein Struct. 1975, 405, 442–451. [Google Scholar] [CrossRef]
- Nascimento, P.F.; Nascimento, A.C.; Rodrigues, C.S.; Antoniolli, Â.R.; Santos, P.O.; Barbosa Júnior, A.M.; Trindade, R.C. Atividade antimicrobiana dos óleos essenciais: Uma abordagem multifatorial dos métodos. Rev. Bras. Farmacogn. 2007, 17, 108–113. [Google Scholar] [CrossRef]
- De Oliveira Pereira, F.; Mendes, J.M.; Lima, I.O.; De Lira Mota, K.S.; De Oliveira, W.A.; De Oliveira Lima, E. Antifungal activity of geraniol and citronellol, two monoterpenes alcohols, against Trichophyton rubrum involves inhibition of ergosterol biosynthesis. Pharm. Biol. 2015, 53, 228–234. [Google Scholar] [CrossRef] [Green Version]
- Humphries, R.M.; Ambler, J.; Mitchell, S.L.; Castanheira, M.; Dingle, T.; Hindler, J.A.; Koeth, L.; Sei, K.; on behalf of the CLSI Methods Development and Standardization Working Group of the Subcommittee on Antimicrobial Susceptibility Testing. CLSI methods development and standardization working group best practices for evaluation of antimicrobial susceptibility tests. J. Clin. Microbiol. 2018, 56, e01934-17. [Google Scholar] [CrossRef] [Green Version]
- Hadacek, F.; Greger, H. Testing of antifungal natural products: Methodologies, comparability of results and assay choice. Phytochem. Anal. 2000, 11, 137–147. [Google Scholar] [CrossRef]
- Freire, I.C.M.; Pérez, A.L.A.L.; Cardoso, A.M.R.; Mariz, B.A.L.A.; Almeida, L.F.D.; Cavalcanti, Y.W.; Padilha, W.W.N. Atividade antibacteriana de Óleos Essenciais sobre Streptococcus mutans e Staphylococcus aureus. Rev. Bras. Plantas Med. 2014, 16, 372–377. [Google Scholar] [CrossRef] [Green Version]
- CLSI; Dolinsky, A.L.; Ohiro, R.K.; Fan, W.; Xiao, C.; Wu, F. National Committee for Clinical Laboratory Standards. 2000. Performance standard for antimicrobial susceptibility testing. Document M100–S10. J. Int. Med. Res. 2019, 46, 18. [Google Scholar]
- Muradás, T.C.; Abbadi, B.L.; Villela, A.D.; Macchi, F.S.; Bergo, P.F.; de Freitas, T.F.; Sperotto, N.D.M.; Timmers, L.F.S.M.; de Souza, O.N.; Picada, J.N.; et al. Pre−clinical evaluation of quinoxaline−derived chalcones in tuberculosis. PLoS ONE 2018, 13, e0202568. [Google Scholar] [CrossRef] [Green Version]
- Sidrônio, M.G.S.; Branco, A.P.O.C.; Abbadi, B.L.; Macchi, F.; Silveira, M.D.; Lock, G.D.A.; Dalla Costa, T.; de Araújo, D.M.; Cibulski, S.; Bizarro, C.V.; et al. Effects of tafenoquine against active, dormant and resistant Mycobacterium tuberculosis. Tuberculosis 2021, 128, 102089. [Google Scholar] [CrossRef] [PubMed]
- Pankey, G.A.; Sabath, L.D. Clinical relevance of bacteriostatic versus bactericidal mechanisms of action in the treatment of Gram−positive bacterial infections. Clin. Infect. Dis. 2004, 38, 864–870. [Google Scholar] [CrossRef] [Green Version]
- Ncube, N.S.; Afolayan, A.J.; Okoh, A.I. Assessment techniques of antimicrobial properties of natural compounds of plant origin: Current methods and future trends. Afr. J. Biotechnol. 2008, 7, 1797–1806. [Google Scholar] [CrossRef] [Green Version]
- Hafidh, R.R. Inhibition of Growth of Highly Resistant Bacterial and Fungal Pathogens by a Natural Product. Open Microbiol. J. 2011, 5, 96–106. [Google Scholar] [CrossRef]
- Benson, D.A.; Karsch−Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2010, 38, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, K.; Kiefer, F.; Kopp, J.; Battey, J.N.D.; Podvinec, M.; Westbrook, J.D.; Berman, H.M.; Bordoli, L.; Schwede, T. The Protein Model Portal. J. Struct. Funct. Genomics 2009, 10, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Eisenberg, D.; Lüthy, R.; Bowie, J.U. VERIFY3D: Assessment of protein models with three−dimensional profiles. Methods Enzymol. 1997, 277, 396–404. [Google Scholar] [PubMed]
- Burley, S.K.; Bhikadiya, C.; Bi, C.; Bittrich, S.; Chen, L.; Crichlow, G.V.; Christie, C.H.; Dalenberg, K.; Di Costanzo, L.; Duarte, J.M.; et al. RCSB Protein Data Bank: Powerful new tools for exploring 3D structures of biological macromolecules for basic and alied research and education in fundamental biology, biomedicine, biotechnology, bioengineering and energy sciences. Nucleic Acids Res. 2021, 49, 437–451. [Google Scholar] [CrossRef]
- Thomsen, R.; Christensen, M.H. MolDock: A new technique for high−accuracy molecular docking. J. Med. Chem. 2006, 49, 3315–3321. [Google Scholar] [CrossRef]
- Bitencourt-Ferreira, G.; Azevedo, W. Molegro Virtual Docker for Docking. In Molegro Virtual Docker for Docking; Springer: Berlin/Heidelberg, Germany, 2019; pp. 149–167. [Google Scholar]
- Vanden Broeck, A.; Lotz, C.; Ortiz, J.; Lamour, V. Cryo−EM structure of the complete E. coli DNA gyrase nucleoprotein complex. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Cutfield, S.M.; Dodson, E.J.; Anderson, B.F.; Moody, P.C.; Marshall, C.J.; Sullivan, P.A.; Cutfield, J.F.; Moody, P.; Moody, P.; Marshall, C. The crystal structure of a major secreted aspartic proteinase from Candida albicans in complexes with two inhibitors. Structure 1995, 3, 1261–1271. [Google Scholar] [CrossRef] [Green Version]
- Catteau, L.; Zhu, L.; Van Bambeke, F.; Quetin−Leclercq, J. Natural and hemi−synthetic pentacyclic triterpenes as antimicrobials and resistance modifying agents against Staphylococcus aureus: A review. Phytochem. Rev. 2018, 17, 1129–1163. [Google Scholar] [CrossRef]
- Junior, A.R.C.; Martins, A.L.D.B.; Cutrim, B.D.S.; Santos, D.M.; Maia, H.S.; da Silva, M.S.M.; Zagmignan, A.; Silva, M.R.C.; Monteiro, C.D.A.; Guilhon, G.M.S.P.; et al. Betulinic acid prevents the acquisition of ciprofloxacin−mediated mutagenesis in Staphylococcus aureus. Molecules 2019, 24, 1–13. [Google Scholar]
- Chung, P.Y. Novel targets of pentacyclic triterpenoids in Staphylococcus aureus: A systematic review. Phytomedicine 2020, 73, 152933. [Google Scholar] [CrossRef] [PubMed]
- Haque, S.; Nawrot, D.A.; Alakurtti, S.; Ghemtio, L.; Yli−Kauhaluoma, J.; Tammela, P. Screening and characterisation of antimicrobial properties of semisynthetic betulin derivatives. PLoS ONE 2014, 9, e102696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germe, T.; Vörös, J.; Jeannot, F.; Taillier, T.; A Stavenger, R.; Bacqué, E.; Maxwell, A.; Bax, B.D. A new class of antibacterials, the imidazopyrazinones, reveal structural transitions involved in DNA gyrase poisoning and mechanisms of resistance. Nucleic Acids Res. 2018, 46, 4114–4128. [Google Scholar] [CrossRef] [Green Version]
- Pozzi, C.; De Luca, F.; Benvenuti, M.; Poirel, L.; Nordmann, P.; Rossolini, G.M.; Mangani, S.; Docquier, J. −D. Crystal structure of the Pseudomonas aeruginosa BEL−1 extended−spectrum β−lactamase and its complexes with moxalactam and imipenem. Antimicrob. Agents Chemother. 2016, 60, 7189–7199. [Google Scholar] [CrossRef] [Green Version]
- Innocente, A.; Casanova, B.B.; Klein, F.; Lana, A.D.; Pereira, D.; Muniz, M.N.; Sonnet, P.; Gosmann, G.; Fuentefria, A.M.; Gnoatto, S.C.B. Synthesis of isosteric triterpenoid derivatives and antifungal activity. Chem. Biol. Drug Des. 2014, 83, 344–349. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
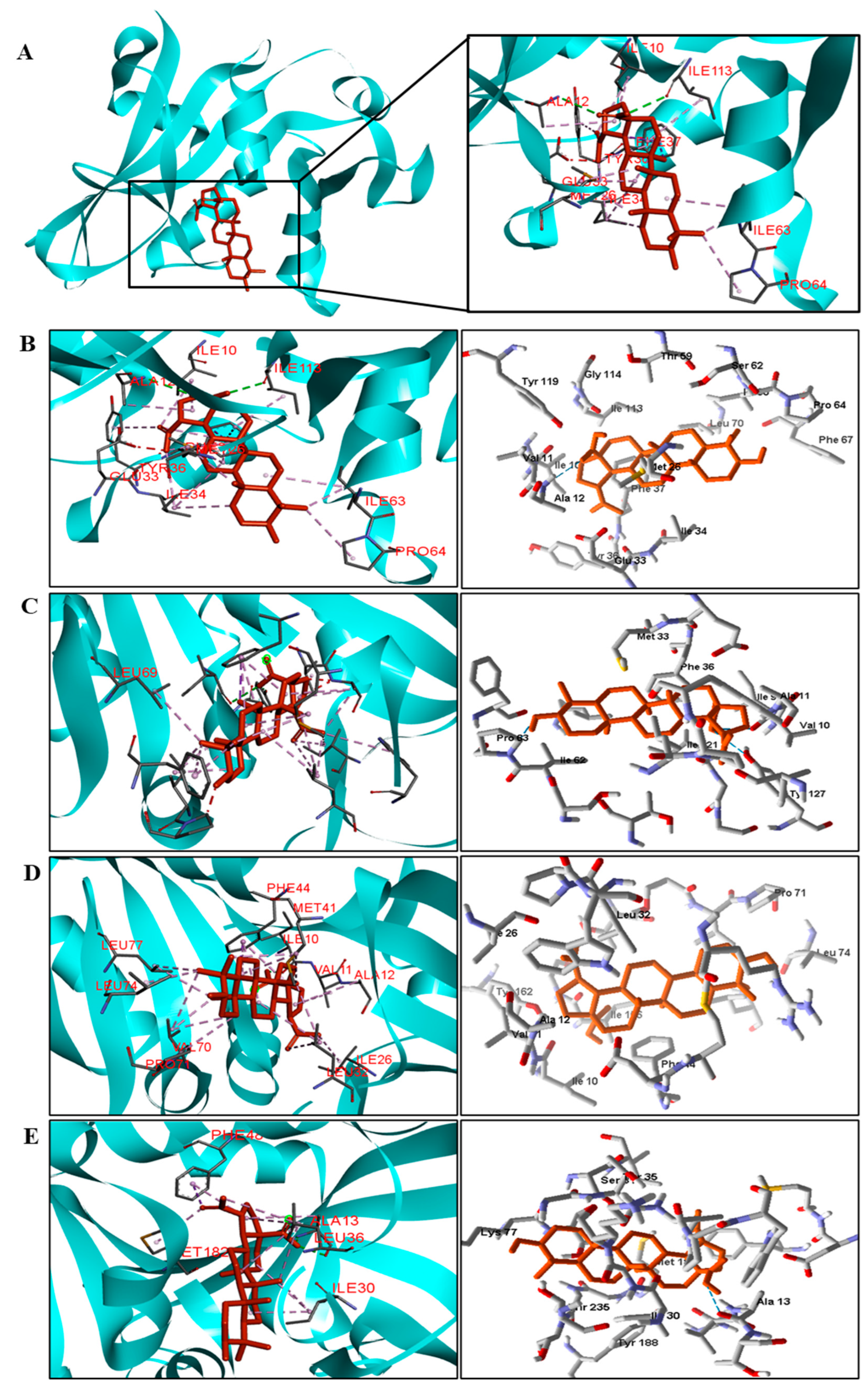{kind=link}
| Species | Protein | PDB ID/Homology | Medication | Resolution |
|---|---|---|---|---|
| DNA gyrase | 6FQM | ciprofloxacin | 3.06 Å | |
| S. aureus | beta-lactamase | 1GHP | sulbactam | 1.76 Å |
| PBP | 3VSL | imipenem | 2.40 Å | |
| DNA gyrase | Homologia | ciprofloxacin | - | |
| S. epidermidis | beta-lactamase | Homologia | sulbactam | - |
| PBP | Homologia | imipenem | - | |
| DNA gyrase | 6M1J | ciprofloxacin | 1.70 Å | |
| P. aeruginosa | beta-lactamase | 5EPH | sulbactam | 1.79 Å |
| PBP | 3PBQ | imipenem | 1.70 Å | |
| DNA gyrase | 5L3J | ciprofloxacin | 2.83 Å | |
| E. coli | beta-lactamase | 1IEM | sulbactam | 2.30 Å |
| PBP | 6G9P | imipenem | 2.10 Å | |
| DNA gyrase | 3IFZ | ciprofloxacin | 2.70 Å | |
| M. tuberculosis | beta-lactamase | 3N7W | sulbactam | 1.70 Å |
| PBP | 6KGW | imipenem | 2.41 Å | |
| CYP | 5FSA | fluconazole | 2.86 Å | |
| C. albicans | SAP-2 | 3PVK | pepstaine | 1.27 Å |
| DHFR | 3QLW | trimethoprim | 2.50 Å | |
| CYP | 6T1U | fluconazol | 1.50 Å | |
| C. tropicalis | SAP-2 | Homologia | pepstaine | - |
| DHFR | Homologia | trimethoprim | - | |
| C. glabrata | CYP | 5JLC | fluconazole | 2.40 Å |
| A. flavus | CYP | Homologia | fluconazole | - |
| DHFR | 6DRS | trimethoprim | 2.00 Å | |
| T. rubrum | DHFR | Homologia | trimethoprim | - |
| Bacterial Strains | Betulinic Acid | Imipenem | Effect | ||
|---|---|---|---|---|---|
| Gram Positive | MIC | MBC | MIC | MBC | |
| Staphylococcus aureus ATCC-13150 | 561 | 1122 | 855 | 1710 | Inhibitory |
| S. epidermidis ATCC-12228 | 561 | 1122 | 855 | 1710 | Inhibitory |
| Gram negative | MIC | MBC | MIC | MBC | |
| Pseudomonas aeruginosa ATCC-25853 | 1122 | 2245 | 1710 | 3420 | - |
| Escherichia coli ATCC-18739 | 561 | 2245 | 1710 | 3420 | Inhibitory |
| Fungal Strains | Betulinic Acid | Fluconazole | Effect | ||
|---|---|---|---|---|---|
| Yeasts | MIC | MFC | MIC | MFC | |
| Candida albicans ATCC-90028 | 561 | 1122 | 835 | 1671 | Inhibitory |
| C. albicans LM-34 | 561 | 1122 | 835 | 1671 | Inhibitory |
| C. tropicalis ATCC-13803 | 561 | 1122 | 835 | 1671 | Inhibitory |
| C. glabrata ATCC-90030 | 561 | 1122 | 835 | 1671 | Inhibitory |
| Filamentous | MIC | MFC | MIC | MFC | Effect |
| Aspergillus flavus ATCC-13013 | R | R | 1671 | 3443 | Resistant |
| Penicillium citrinum ATCC-40011 | 1122 | 2245 | 1671 | 3443 | Inhibitory |
| Trichophyton rubrum LM-34 | 561 | 1122 | 835 | 1671 | Inhibitory |
| Microsporum canis LM-12 | 561 | 1122 | 835 | 1671 | Inhibitory |
| Compounds | MIC in µM |
|---|---|
| betulinic acid | 100 |
| moxifloxacin | 0.19 |
| rifampin | 0.15 |
| Species | DNA Gyrase | Beta-Lactamase | PBP | |||
|---|---|---|---|---|---|---|
| Betulinic Acid | Ciprofloxacin | Betulinic Acid | Sulbactam | Betulinic Acid | Imipenem | |
| S. aureus | −109.22 | −96.78 | −100.68 | −69.29 | −82 | −103.63 |
| S. epidermidis | −73.89 | −101.58 | −118.61 | −68.33 | −76.51 | −100.61 |
| P. aeruginosa | −107.14 | −121.73 | −112.71 | −72.25 | −107.59 | −113.99 |
| E. coli | −99.59 | −89.57 | −101.68 | −68.35 | −71.63 | −91.57 |
| M. tuberculosis | −114.43 | −78.99 | −86.77 | −73.68 | −73.29 | −100 |
| Species | CYP51 | SAP−2 | DHFR | |||
|---|---|---|---|---|---|---|
| Betulinic Acid | Fluconazole | Betulinic Acid | Pepstaine | Betulinic Acid | Trimethoprim | |
| C. albicans | −120.98 | −112.67 | −46.73 | −84.10 | −68.47 | −82.96 |
| C. tropicalis | −113.31 | −119.54 | −99.36 | −129.96 | −87.25 | −92.43 |
| C. glabrata | −130.95 | −77.29 | − | − | −96.25 | −83.97 |
| Aspergillus flavus | −120.26 | −125.17 | − | − | −103.16 | −87.71 |
| Penicillium citrinum | − | − | − | − | − | − |
| Trichophyton rubrum | − | − | − | − | −151.48 | −108.78 |
| Microsporum canis | − | − | − | − | − | − |
| Species | DNA Gyrase | |
|---|---|---|
| Hydrogen Bonds | Hydrophobic Interactions | |
| S. aureus | Glu88. | Arg458 and Glu477. |
| E. coli | − | Val43, His55, Arg76, Pro79, Val120, Val167. |
| M. tuberculosis | − | Pro42, Leu48, Lys49, His52, Arg98, Leu105 e Tyr276. |
| Species | Beta−Lactamase | |
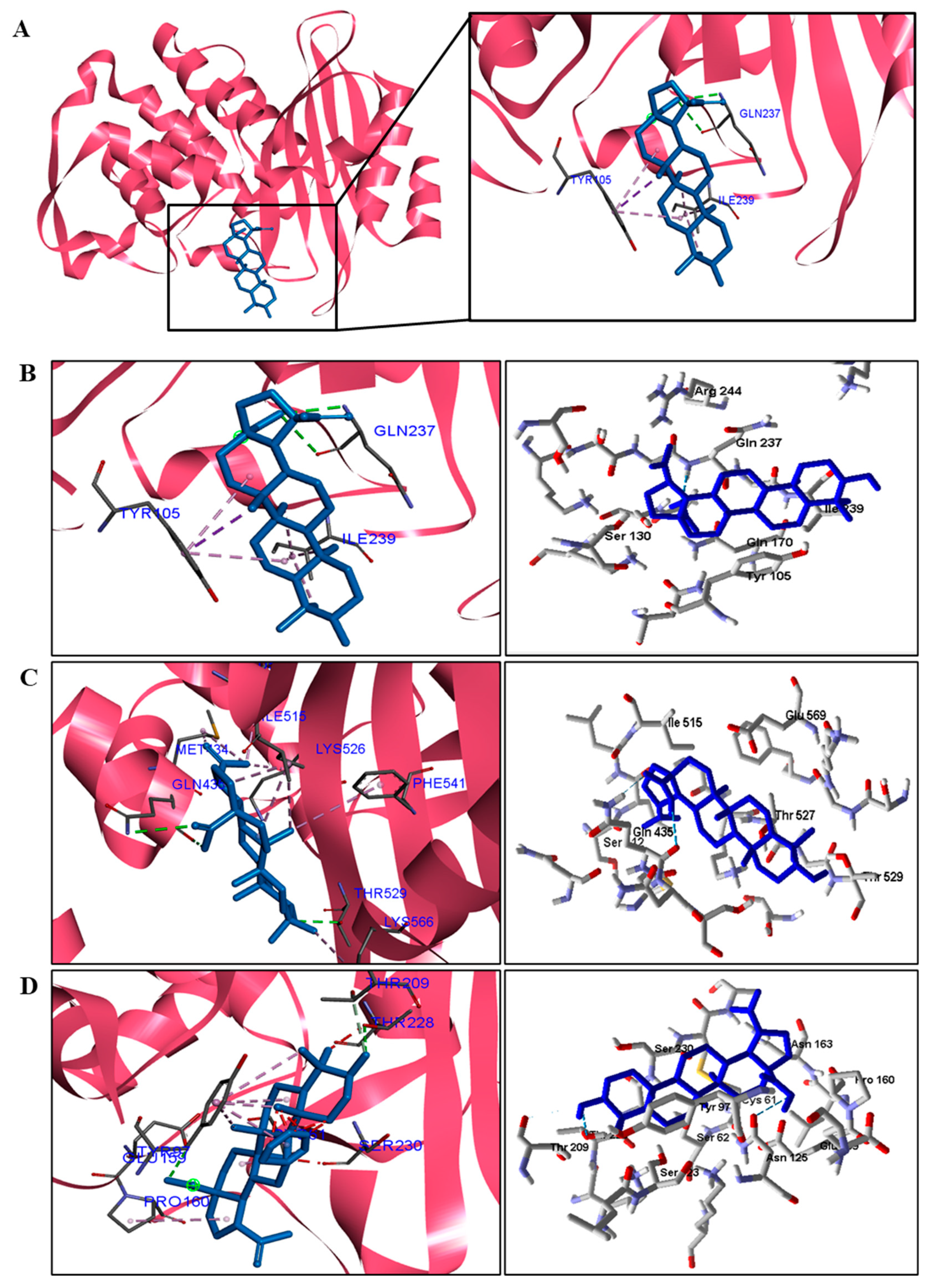
| S. aureus | Gln237. | Ala69, Tyr105, Asn132, and Ile239. |
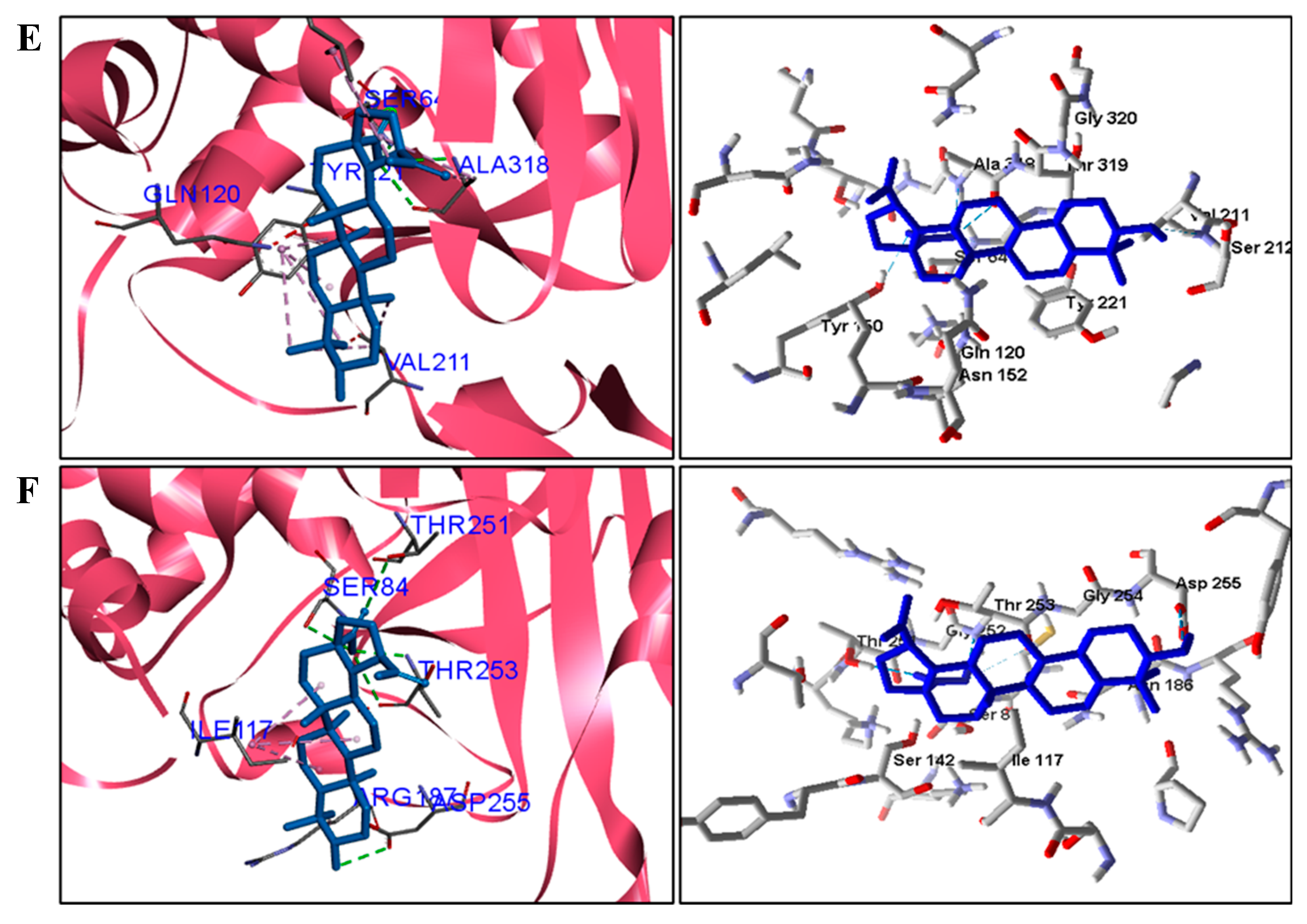
| S. epidermidis | Gln435 and Thr529. | Met434, Ile515, Lys526, Phe541 e Lys566. |
| P. aeruginosa | Thr159 and Thr228. | Cys61, Tyr97 e Pro160. |
| E. coli | Ser64 and Ala318. | Tyr221 and Leu293. |
| M. tuberculosis | Ser84, Thr251, and Asp255. | Ile117 e Arg187. |
| Species | PBP | |
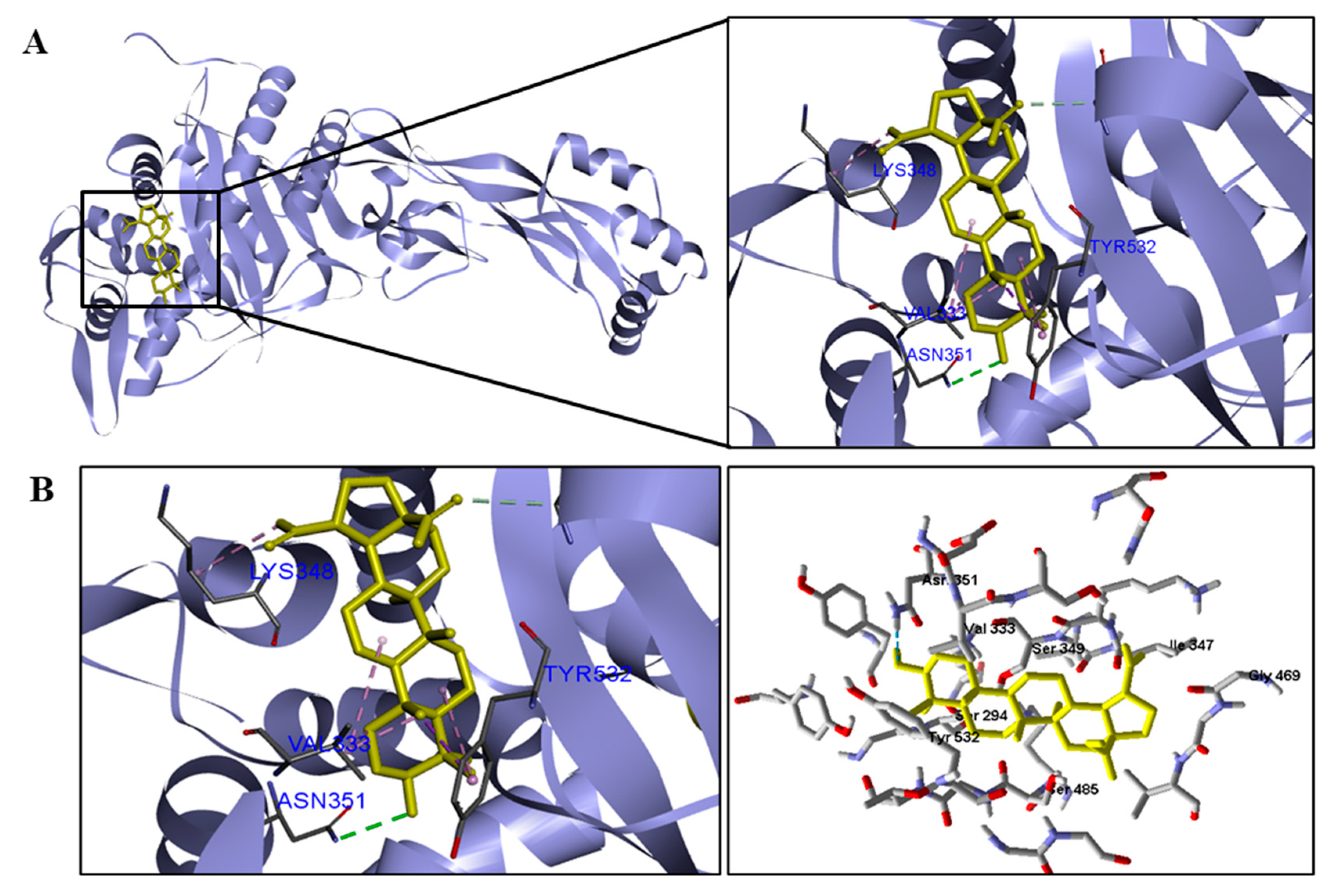
| P. aeruginosa | Asn351. | Val333, Lys348 and Tyr532. |
| Species | CYP51 | |
|---|---|---|
| Hydrogen Bonds | Hydrophobic Interactions | |
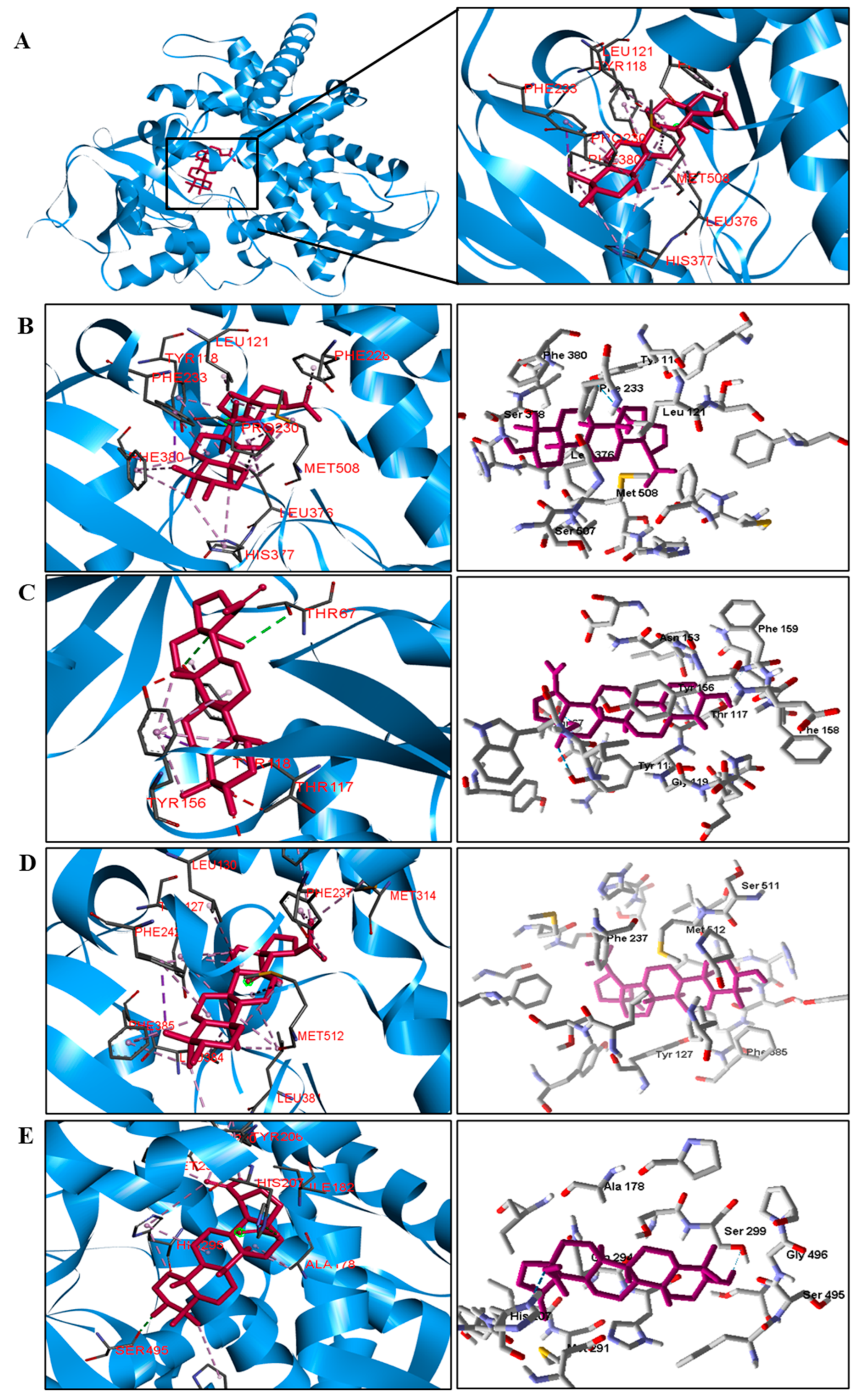
| C. albicans | Tyr118, Phe228, Pro230, Leu376, Hist377, Phe380 and Met508. | |
| C. tropicalis | Tyr 156. | Thr67, and Tyr118. |
| C. glabrata | − | Tyr127, Leu130, Phe135, Phe237, Met314, Phe385, Leu381, His382, and Met512. |
| His207 and Ser495. | Ala178, Ile182, Tyr206, His295, Leu290, Met291, and Pro497. | |
| Species | DHFR | |
| C. tropicalis | Ala12 and Ile113. | Ile10, Met26, Ile34, Tyr36, Ile63, and Pro64. |
| C. glabrata | Ile121 and Tyr127. | Val10, Ile19, Leu25, Trp27, Met33, Ile62, Phe66, and Leu69. |
| A. flavus | − | Val11, Ala12, Ile26, Leu32, Met41, Phe44, Val70, Leu74 e Leu77. |
| T. rubrum | − | Ala13, Ile30, Leu36, Phe48, and Met182. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, G.C.S.; dos Santos Maia, M.; de Souza, T.A.; de Oliveira Lima, E.; dos Santos, L.E.C.G.; Silva, S.L.; da Silva, M.S.; Filho, J.M.B.; da Silva Rodrigues Junior, V.; Scotti, L.; et al. Antimicrobial Potential of Betulinic Acid and Investigation of the Mechanism of Action against Nuclear and Metabolic Enzymes with Molecular Modeling. Pathogens 2023, 12, 449. https://doi.org/10.3390/pathogens12030449
Rodrigues GCS, dos Santos Maia M, de Souza TA, de Oliveira Lima E, dos Santos LECG, Silva SL, da Silva MS, Filho JMB, da Silva Rodrigues Junior V, Scotti L, et al. Antimicrobial Potential of Betulinic Acid and Investigation of the Mechanism of Action against Nuclear and Metabolic Enzymes with Molecular Modeling. Pathogens. 2023; 12(3):449. https://doi.org/10.3390/pathogens12030449
Chicago/Turabian StyleRodrigues, Gabriela Cristina Soares, Mayara dos Santos Maia, Thalisson Amorim de Souza, Edeltrudes de Oliveira Lima, Luiz Eduardo Carneiro Gomes dos Santos, Shellygton Lima Silva, Marcelo Sobral da Silva, José Maria Barbosa Filho, Valnês da Silva Rodrigues Junior, Luciana Scotti, and et al. 2023. "Antimicrobial Potential of Betulinic Acid and Investigation of the Mechanism of Action against Nuclear and Metabolic Enzymes with Molecular Modeling" Pathogens 12, no. 3: 449. https://doi.org/10.3390/pathogens12030449
APA StyleRodrigues, G. C. S., dos Santos Maia, M., de Souza, T. A., de Oliveira Lima, E., dos Santos, L. E. C. G., Silva, S. L., da Silva, M. S., Filho, J. M. B., da Silva Rodrigues Junior, V., Scotti, L., & Scotti, M. T. (2023). Antimicrobial Potential of Betulinic Acid and Investigation of the Mechanism of Action against Nuclear and Metabolic Enzymes with Molecular Modeling. Pathogens, 12(3), 449. https://doi.org/10.3390/pathogens12030449