
Toll-like Receptor-9 (TLR-9) Signaling Is Crucial for Inducing Protective Immunity following Immunization with Genetically Modified Live Attenuated Leishmania Parasites

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals and Parasites
2.2. Ethics Statement
2.3. Infection of Mice and Isolation of Macrophages and DCs from Ear dLN and Spleen
2.4. Cultivation of Bone Marrow-Derived Dendritic Cells (BMDCs)
2.5. TLR-9 siRNA and MyD88 siRNA Mediated Silencing in DC In Vitro
2.6. NF-κB Nuclear Translocation
2.7. Antigen Presentation Assay: In Vitro DC and T Cell Co-Culture Studies
2.8. RT-PCR
2.9. Flow Cytometry
2.10. Immunization and Challenge Studies
2.11. Statistical Analysis
3. Results
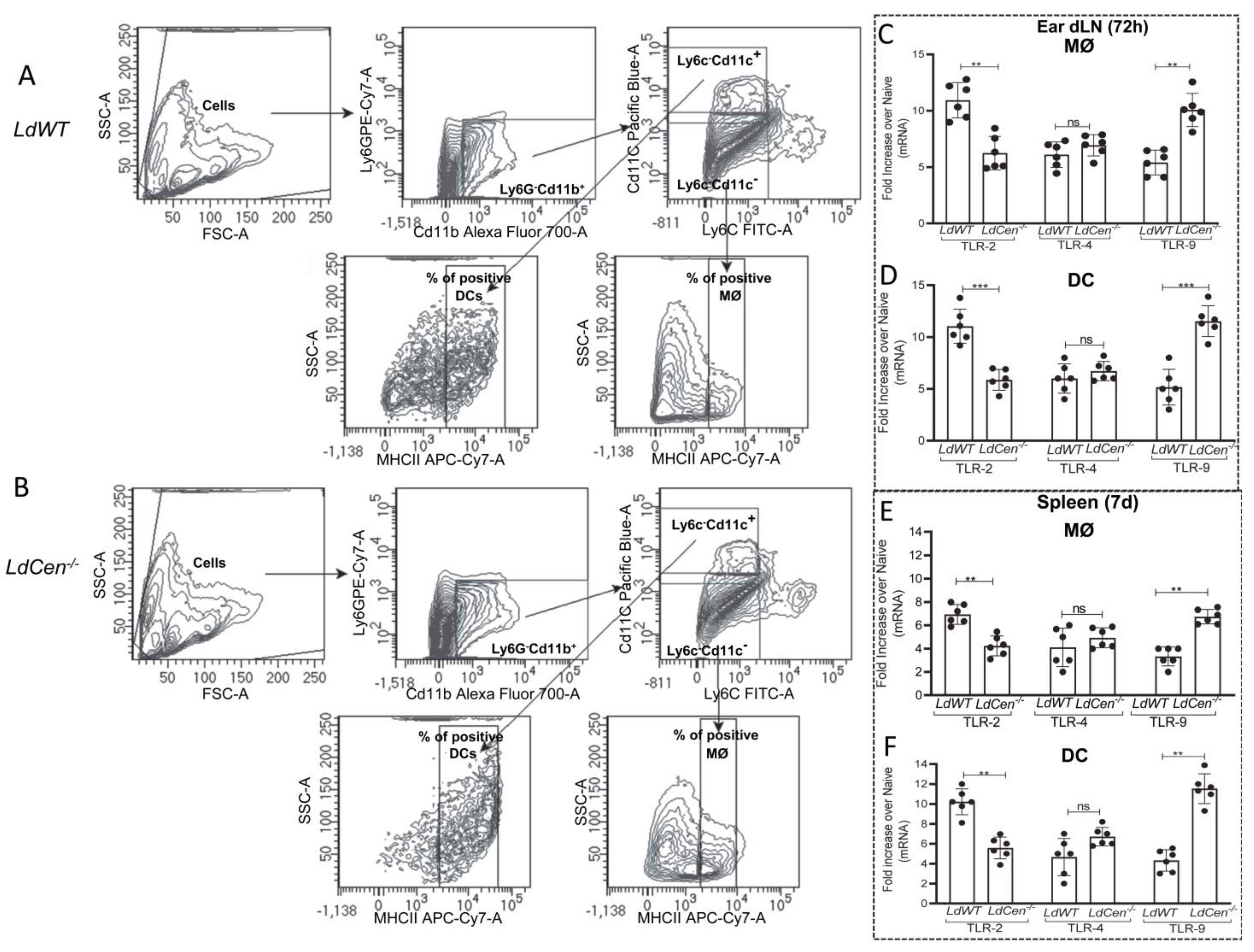
3.1. Infection with LdCen−/− Induces TLR-9 mRNA Expression
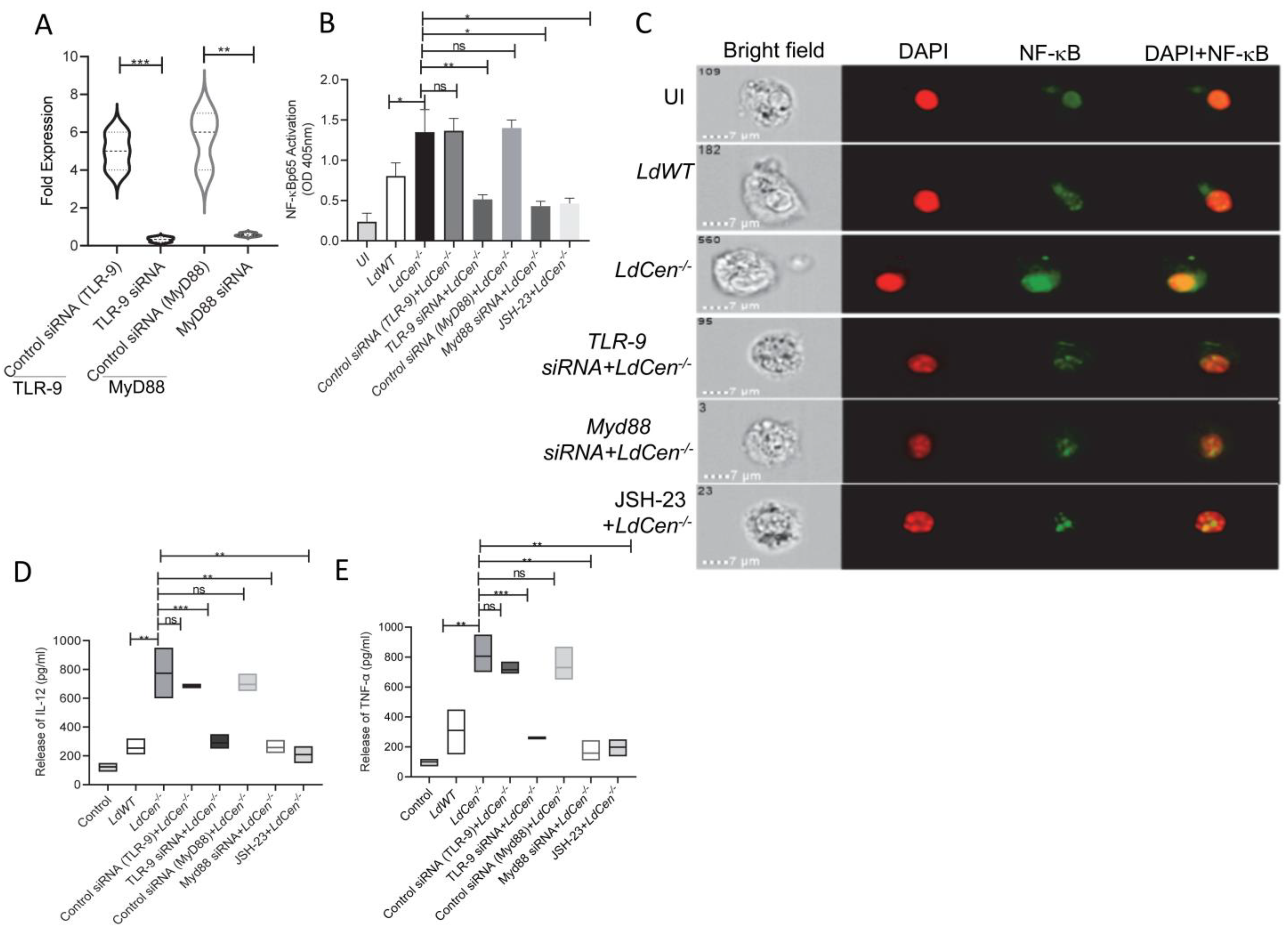
3.2. LdCen−/− Infected DCs Showed Heighted NF-κB Activation and Proinflammatory Cytokine Response via TLR-9- Myd88 Pathway
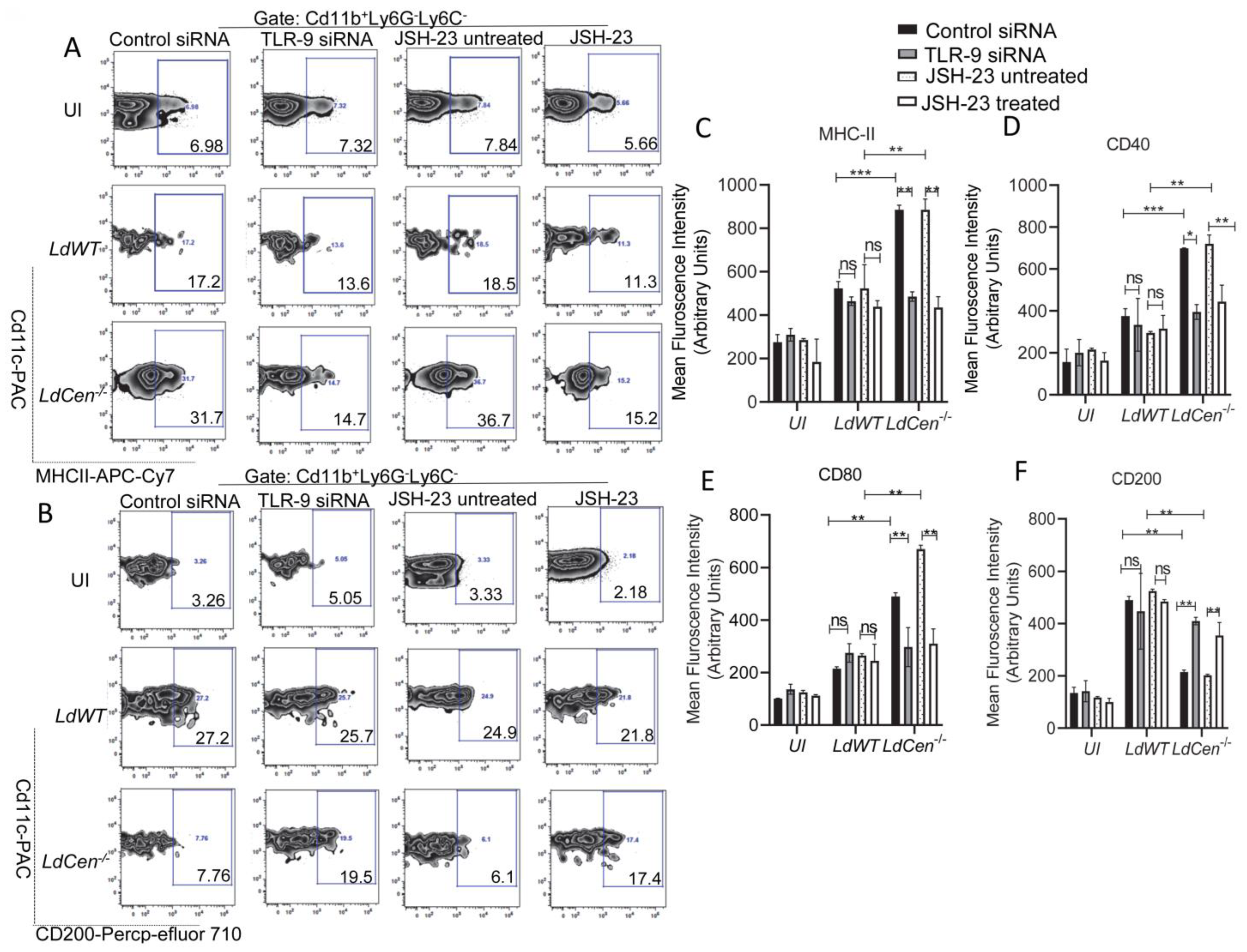
3.3. TLR-9 Silencing or Inhibition of NF-κB Transcriptional Activity Significantly Reduced the Expression of Costimulatory Molecules While Augmenting Expression of Coinhibitory Molecule in LdCen−/− Infected DCs
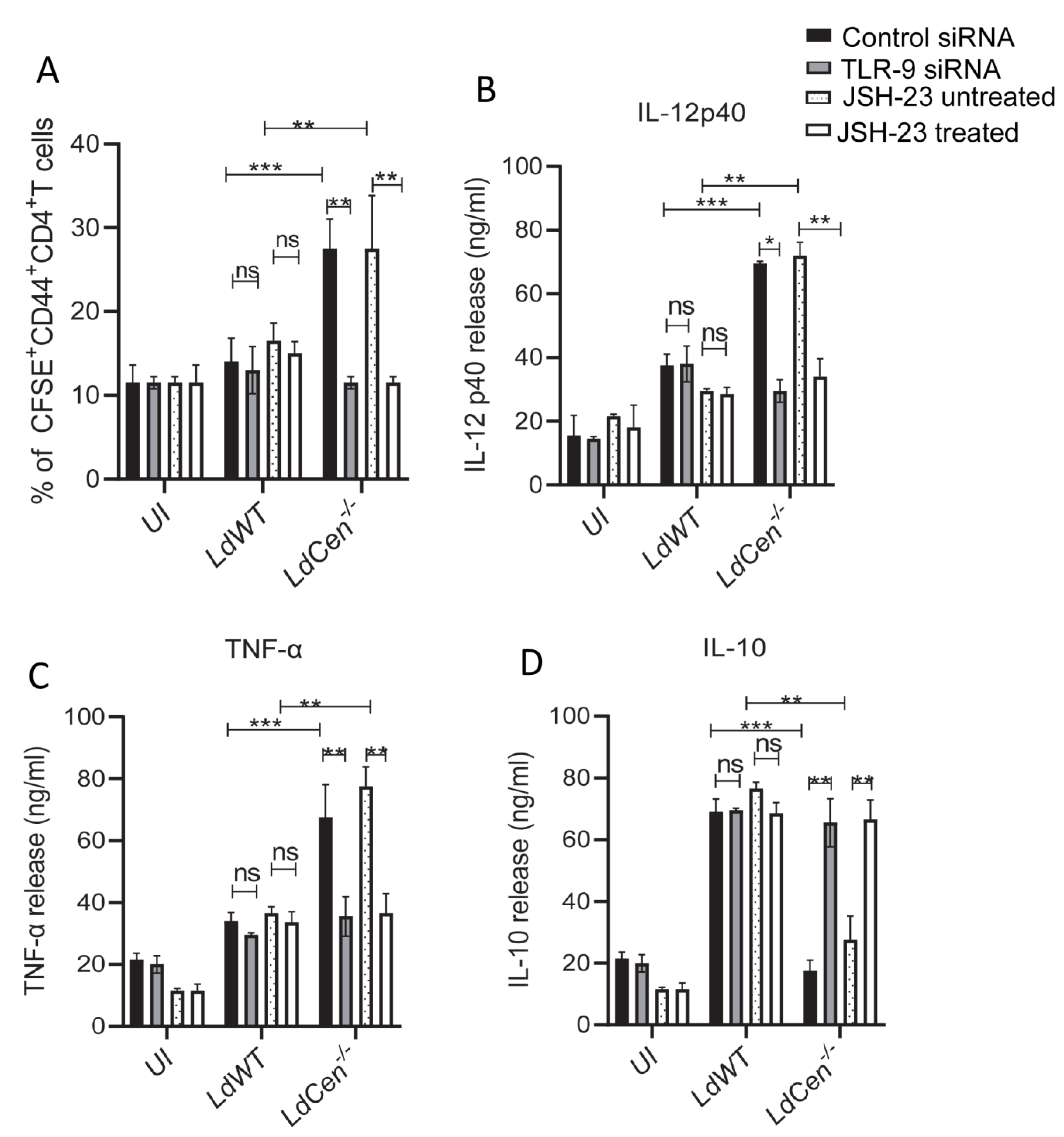
3.4. TLR-9 Silencing or Inhibition of NF-κB Transcriptional Activity Significantly Reduced LdCen−/− Infected DC Mediated T Cell Proliferation and Altered Cytokine Expression
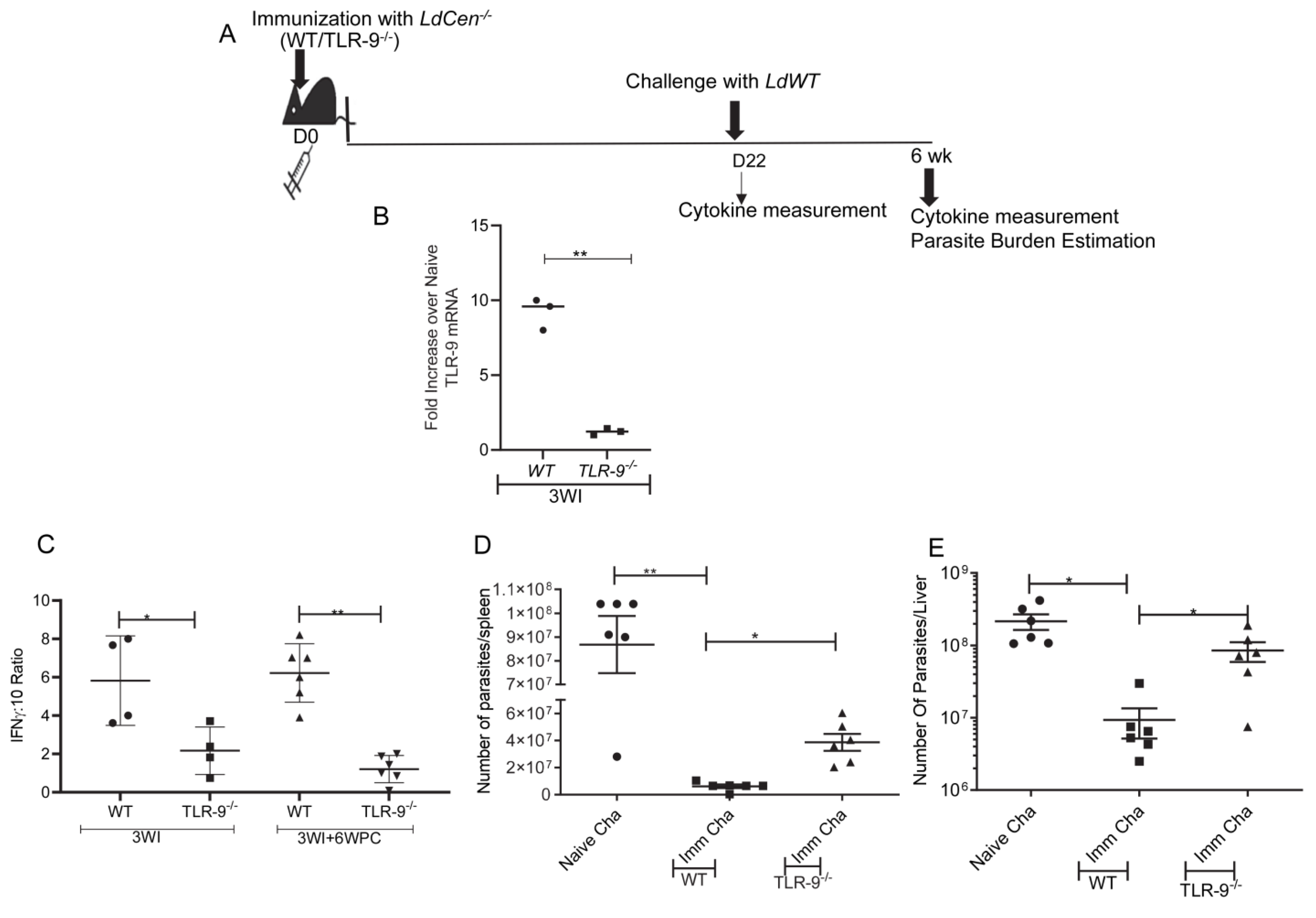
3.5. Absence of TLR-9 Abrogates LdCen−/− Induced Protection against LdWT Infection
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grifferty, G.; Shirley, H.; McGloin, J.; Kahn, J.; Orriols, A.; Wamai, R. Vulnerabilities to and the Socioeconomic and Psychosocial Impacts of the Leishmaniases: A Review. Res. Rep. Trop. Med. 2021, 12, 135–151. [Google Scholar] [CrossRef]
- Elmahallawy, E.K.; Alkhaldi, A.A.M.; Saleh, A.A. Host immune response against leishmaniasis and parasite persistence strategies: A review and assessment of recent research. Biomed. Pharmacother. 2021, 139, 111671. [Google Scholar] [CrossRef]
- Sacramento, L.; Trevelin, S.C.; Nascimento, M.S.; Lima-Junior, D.S.; Costa, D.L.; Almeida, R.P.; Cunha, F.Q.; Silva, J.S.; Carregaro, V. Toll-like receptor 9 signaling in dendritic cells regulates neutrophil recruitment to inflammatory foci following Leishmania infantum infection. Infect. Immun. 2015, 83, 4604–4616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bafica, A.; Santiago, H.C.; Goldszmid, R.; Ropert, C.; Gazzinelli, R.T.; Sher, A. Cutting edge: TLR9 and TLR2 signaling together account for MyD88-dependent control of parasitemia in Trypanosoma cruzi infection. J. Immunol. 2006, 177, 3515–3519. [Google Scholar] [CrossRef] [Green Version]
- Debierre-Grockiego, F.; Azzouz, N.; Schmidt, J.; Dubremetz, J.F.; Geyer, H.; Geyer, R.; Weingart, R.; Schmidt, R.R.; Schwarz, R.T. Roles of glycosylphosphatidylinositols of Toxoplasma gondii. Induction of tumor necrosis factor-alpha production in macrophages. J. Biol. Chem. 2003, 278, 32987–32993. [Google Scholar] [CrossRef] [Green Version]
- Shoda, L.K.; Kegerreis, K.A.; Suarez, C.E.; Roditi, I.; Corral, R.S.; Bertot, G.M.; Norimine, J.; Brown, W.C. DNA from protozoan parasites Babesia bovis, Trypanosoma cruzi, and T. brucei is mitogenic for B lymphocytes and stimulates macrophage expression of interleukin-12, tumor necrosis factor alpha, and nitric oxide. Infect. Immun. 2001, 69, 2162–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, M.A.; Almeida, I.C.; Takeuchi, O.; Akira, S.; Valente, E.P.; Procopio, D.O.; Travassos, L.R.; Smith, J.A.; Golenbock, D.T.; Gazzinelli, R.T. Activation of Toll-like receptor-2 by glycosylphosphatidylinositol anchors from a protozoan parasite. J. Immunol. 2001, 167, 416–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijay, K. Toll-like receptors in immunity and inflammatory diseases: Past, present, and future. Int. Immunopharmacol. 2018, 59, 391–412. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Faria, M.S.; Reis, F.C.; Lima, A.P. Toll-like receptors in leishmania infections: Guardians or promoters? J. Parasitol. Res. 2012, 2012, 930257. [Google Scholar] [CrossRef] [Green Version]
- Muraille, E.; De Trez, C.; Brait, M.; De Baetselier, P.; Leo, O.; Carlier, Y. Genetically resistant mice lacking MyD88-adapter protein display a high susceptibility to Leishmania major infection associated with a polarized Th2 response. J. Immunol. 2003, 170, 4237–4241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Trez, C.; Brait, M.; Leo, O.; Aebischer, T.; Torrentera, F.A.; Carlier, Y.; Muraille, E. Myd88-dependent in vivo maturation of splenic dendritic cells induced by Leishmania donovani and other Leishmania species. Infect. Immun. 2004, 72, 824–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flandin, J.F.; Chano, F.; Descoteaux, A. RNA interference reveals a role for TLR2 and TLR3 in the recognition of Leishmania donovani promastigotes by interferon-gamma-primed macrophages. Eur. J. Immunol. 2006, 36, 411–420. [Google Scholar] [CrossRef]
- de Veer, M.J.; Curtis, J.M.; Baldwin, T.M.; DiDonato, J.A.; Sexton, A.; McConville, M.J.; Handman, E.; Schofield, L. MyD88 is essential for clearance of Leishmania major: Possible role for lipophosphoglycan and Toll-like receptor 2 signaling. Eur. J. Immunol. 2003, 33, 2822–2831. [Google Scholar] [CrossRef]
- Osanya, A.; Song, E.H.; Metz, K.; Shimak, R.M.; Boggiatto, P.M.; Huffman, E.; Johnson, C.; Hostetter, J.M.; Pohl, N.L.; Petersen, C.A. Pathogen-derived oligosaccharides improve innate immune response to intracellular parasite infection. Am. J. Pathol. 2011, 179, 1329–1337. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, P.; Bhattacharjee, S.; Gupta, G.; Majumder, S.; Adhikari, A.; Mukherjee, A.; Majumdar, S.B.; Saha, B.; Majumdar, S. Arabinosylated lipoarabinomannan-mediated protection in visceral leishmaniasis through up-regulation of toll-like receptor 2 signaling: An immunoprophylactic approach. J. Infect. Dis. 2010, 202, 145–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kropf, P.; Freudenberg, M.A.; Modolell, M.; Price, H.P.; Herath, S.; Antoniazi, S.; Galanos, C.; Smith, D.F.; Muller, I. Toll-like receptor 4 contributes to efficient control of infection with the protozoan parasite Leishmania major. Infect. Immun. 2004, 72, 1920–1928. [Google Scholar] [CrossRef] [Green Version]
- Kropf, P.; Freudenberg, N.; Kalis, C.; Modolell, M.; Herath, S.; Galanos, C.; Freudenberg, M.; Muller, I. Infection of C57BL/10ScCr and C57BL/10ScNCr mice with Leishmania major reveals a role for Toll-like receptor 4 in the control of parasite replication. J. Leukoc. Biol. 2004, 76, 48–57. [Google Scholar] [CrossRef]
- Abou Fakher, F.H.; Rachinel, N.; Klimczak, M.; Louis, J.; Doyen, N. TLR9-dependent activation of dendritic cells by DNA from Leishmania major favors Th1 cell development and the resolution of lesions. J. Immunol. 2009, 182, 1386–1396. [Google Scholar] [CrossRef] [Green Version]
- Liese, J.; Schleicher, U.; Bogdan, C. The innate immune response against Leishmania parasites. Immunobiology 2008, 213, 377–387. [Google Scholar] [CrossRef]
- Schleicher, U.; Liese, J.; Knippertz, I.; Kurzmann, C.; Hesse, A.; Heit, A.; Fischer, J.A.; Weiss, S.; Kalinke, U.; Kunz, S.; et al. NK cell activation in visceral leishmaniasis requires TLR9, myeloid DCs, and IL-12, but is independent of plasmacytoid DCs. J. Exp. Med. 2007, 204, 893–906. [Google Scholar] [CrossRef] [PubMed]
- Raman, V.S.; Duthie, M.S.; Fox, C.B.; Matlashewski, G.; Reed, S.G. Adjuvants for Leishmania vaccines: From models to clinical application. Front. Immunol. 2012, 3, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noorpisheh Ghadimi, S.; Farjadian, S.; Hatam, G.R.; Kalani, M.; Sarkari, B. Vaccination with Live Attenuated L. Major and TLR4 Agonist Promotes a Th1 Immune Response and Induces Protection against L. Major Infection in BALB/c Mice. Iran. J. Immunol. 2018, 15, 74–83. [Google Scholar] [PubMed]
- Saljoughian, N.; Taheri, T.; Rafati, S. Live vaccination tactics: Possible approaches for controlling visceral leishmaniasis. Front. Immunol. 2014, 5, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabala-Penafiel, A.; Todd, D.; Daneshvar, H.; Burchmore, R. The potential of live attenuated vaccines against Cutaneous Leishmaniasis. Exp. Parasitol. 2020, 210, 107849. [Google Scholar] [CrossRef]
- Gannavaram, S.; Dey, R.; Avishek, K.; Selvapandiyan, A.; Salotra, P.; Nakhasi, H.L. Biomarkers of safety and immune protection for genetically modified live attenuated leishmania vaccines against visceral leishmaniasis—Discovery and implications. Front. Immunol. 2014, 5, 241. [Google Scholar] [CrossRef] [Green Version]
- Selvapandiyan, A.; Debrabant, A.; Duncan, R.; Muller, J.; Salotra, P.; Sreenivas, G.; Salisbury, J.L.; Nakhasi, H.L. Centrin gene disruption impairs stage-specific basal body duplication and cell cycle progression in Leishmania. J. Biol. Chem. 2004, 279, 25703–25710. [Google Scholar] [CrossRef] [Green Version]
- Dey, R.; Natarajan, G.; Bhattacharya, P.; Cummings, H.; Dagur, P.K.; Terrazas, C.; Selvapandiyan, A.; McCoy, J.P., Jr.; Duncan, R.; Satoskar, A.R.; et al. Characterization of cross-protection by genetically modified live-attenuated Leishmania donovani parasites against Leishmania mexicana. J. Immunol. 2014, 193, 3513–3527. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.W.; Karmakar, S.; Gannavaram, S.; Dey, R.; Lypaczewski, P.; Ismail, N.; Siddiqui, A.; Simonyan, V.; Oliveira, F.; Coutinho-Abreu, I.V.; et al. A second generation leishmanization vaccine with a markerless attenuated Leishmania major strain using CRISPR gene editing. Nat. Commun. 2020, 11, 3461. [Google Scholar] [CrossRef]
- Gannavaram, S.; Davey, S.; Lakhal-Naouar, I.; Duncan, R.; Nakhasi, H.L. Deletion of ubiquitin fold modifier protein Ufm1 processing peptidase Ufsp in L. donovani abolishes Ufm1 processing and alters pathogenesis. PLoS Negl. Trop. Dis. 2014, 8, e2707. [Google Scholar] [CrossRef]
- Selvapandiyan, A.; Dey, R.; Nylen, S.; Duncan, R.; Sacks, D.; Nakhasi, H.L. Intracellular replication-deficient Leishmania donovani induces long lasting protective immunity against visceral leishmaniasis. J. Immunol. 2009, 183, 1813–1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiuza, J.A.; Santiago Hda, C.; Selvapandiyan, A.; Gannavaram, S.; Ricci, N.D.; Bueno, L.L.; Bartholomeu, D.C.; Correa-Oliveira, R.; Nakhasi, H.L.; Fujiwara, R.T. Induction of immunogenicity by live attenuated Leishmania donovani centrin deleted parasites in dogs. Vaccine 2013, 31, 1785–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiuza, J.A.; Gannavaram, S.; Santiago Hda, C.; Selvapandiyan, A.; Souza, D.M.; Passos, L.S.; de Mendonca, L.Z.; Lemos-Giunchetti Dda, S.; Ricci, N.D.; Bartholomeu, D.C.; et al. Vaccination using live attenuated Leishmania donovani centrin deleted parasites induces protection in dogs against Leishmania infantum. Vaccine 2015, 33, 280–288. [Google Scholar] [CrossRef]
- Fiuza, J.A.; Dey, R.; Davenport, D.; Abdeladhim, M.; Meneses, C.; Oliveira, F.; Kamhawi, S.; Valenzuela, J.G.; Gannavaram, S.; Nakhasi, H.L. Intradermal Immunization of Leishmania donovani Centrin Knock-Out Parasites in Combination with Salivary Protein LJM19 from Sand Fly Vector Induces a Durable Protective Immune Response in Hamsters. PLoS Negl. Trop. Dis. 2016, 10, e0004322. [Google Scholar] [CrossRef]
- Bhattacharya, P.; Dey, R.; Saxena, A.; Karmakar, S.; Ismail, N.; Gannavaram, S.; Dagur, P.K.; Satoskar, M.; Satoskar, S.; De Paoli, S.; et al. Essential Role of Neutrophils in the Protective Immune Response Induced by a Live Attenuated Leishmania Vaccine. J. Immunol. 2020, 205, 3333–3347. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, P.; Dey, R.; Dagur, P.K.; Kruhlak, M.; Ismail, N.; Debrabant, A.; Joshi, A.B.; Akue, A.; Kukuruga, M.; Takeda, K.; et al. Genetically Modified Live Attenuated Leishmania donovani Parasites Induce Innate Immunity through Classical Activation of Macrophages That Direct the Th1 Response in Mice. Infect. Immun. 2015, 83, 3800–3815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, P.; Ismail, N.; Saxena, A.; Gannavaram, S.; Dey, R.; Oljuskin, T.; Akue, A.; Takeda, K.; Yu, J.; Karmakar, S.; et al. Neutrophil-dendritic cell interaction plays an important role in live attenuated Leishmania vaccine induced immunity. PLoS Negl. Trop. Dis. 2022, 16, e0010224. [Google Scholar] [CrossRef]
- Mogensen, T.H. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef] [Green Version]
- Daifalla, N.S.; Bayih, A.G.; Gedamu, L. Leishmania donovani recombinant iron superoxide dismutase B1 protein in the presence of TLR-based adjuvants induces partial protection of BALB/c mice against Leishmania major infection. Exp. Parasitol. 2012, 131, 317–324. [Google Scholar] [CrossRef]
- Rhee, E.G.; Mendez, S.; Shah, J.A.; Wu, C.Y.; Kirman, J.R.; Turon, T.N.; Davey, D.F.; Davis, H.; Klinman, D.M.; Coler, R.N.; et al. Vaccination with heat-killed leishmania antigen or recombinant leishmanial protein and CpG oligodeoxynucleotides induces long-term memory CD4+ and CD8+ T cell responses and protection against leishmania major infection. J. Exp. Med. 2002, 195, 1565–1573. [Google Scholar] [CrossRef] [Green Version]
- Tewary, P.; Pandya, J.; Mehta, J.; Sukumaran, B.; Madhubala, R. Vaccination with Leishmania soluble antigen and immunostimulatory oligodeoxynucleotides induces specific immunity and protection against Leishmania donovani infection. FEMS Immunol. Med. Microbiol. 2004, 42, 241–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tewary, P.; Sukumaran, B.; Saxena, S.; Madhubala, R. Immunostimulatory oligodeoxynucleotides are potent enhancers of protective immunity in mice immunized with recombinant ORFF leishmanial antigen. Vaccine 2004, 22, 3053–3060. [Google Scholar] [CrossRef]
- Lanza, J.S.; Vucen, S.; Flynn, O.; Donadei, A.; Cojean, S.; Loiseau, P.M.; Fernandes, A.; Frezard, F.; Moore, A.C. A TLR9-adjuvanted vaccine formulated into dissolvable microneedle patches or cationic liposomes protects against leishmaniasis after skin or subcutaneous immunization. Int. J. Pharm. 2020, 586, 119390. [Google Scholar] [CrossRef] [PubMed]
- Shah, J.A.; Darrah, P.A.; Ambrozak, D.R.; Turon, T.N.; Mendez, S.; Kirman, J.; Wu, C.Y.; Glaichenhaus, N.; Seder, R.A. Dendritic cells are responsible for the capacity of CpG oligodeoxynucleotides to act as an adjuvant for protective vaccine immunity against Leishmania major in mice. J. Exp. Med. 2003, 198, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Spath, G.F.; Beverley, S.M. A lipophosphoglycan-independent method for isolation of infective Leishmania metacyclic promastigotes by density gradient centrifugation. Exp. Parasitol. 2001, 99, 97–103. [Google Scholar] [CrossRef]
- Campos, M.B.; Lima, L.; de Lima, A.C.S.; Vasconcelos Dos Santos, T.; Ramos, P.K.S.; Gomes, C.M.C.; Silveira, F.T. Toll-like receptors 2, 4, and 9 expressions over the entire clinical and immunopathological spectrum of American cutaneous leishmaniasis due to Leishmania (V.) braziliensis and Leishmania (L.) amazonensis. PLoS ONE 2018, 13, e0194383. [Google Scholar] [CrossRef]
- Tiburcio, R.; Nunes, S.; Nunes, I.; Rosa Ampuero, M.; Silva, I.B.; Lima, R.; Machado Tavares, N.; Brodskyn, C. Molecular Aspects of Dendritic Cell Activation in Leishmaniasis: An Immunobiological View. Front. Immunol. 2019, 10, 227. [Google Scholar] [CrossRef] [Green Version]
- Reinhard, K.; Huber, M.; Lohoff, M.; Visekruna, A. The role of NF-kappaB activation during protection against Leishmania infection. Int. J. Med. Microbiol. 2012, 302, 230–235. [Google Scholar] [CrossRef]
- Martinez-Orellana, P.; Montserrat-Sangra, S.; Quirola-Amores, P.; Gonzalez, N.; Solano-Gallego, L. Cytokine Effect of TLR3, TLR4, and TLR7 Agonists Alone or Associated with Leishmania infantum Antigen on Blood from Dogs. BioMed Res. Int. 2018, 2018, 5693736. [Google Scholar] [CrossRef] [Green Version]
- Daifalla, N.S.; Bayih, A.G.; Gedamu, L. Immunogenicity of Leishmania donovani iron superoxide dismutase B1 and peroxidoxin 4 in BALB/c mice: The contribution of Toll-like receptor agonists as adjuvant. Exp. Parasitol. 2011, 129, 292–298. [Google Scholar] [CrossRef]
- Pratti, J.E.S.; da Fonseca Martins, A.M.; da Silva, J.P.; Ramos, T.D.; Pereira, J.C.; Firmino-Cruz, L.; Oliveira-Maciel, D.; Vieira, T.S.S.; Lacerda, L.L.; Vale, A.M.; et al. The role of TLR9 on Leishmania amazonensis infection and its influence on intranasal LaAg vaccine efficacy. PLoS Negl. Trop. Dis. 2019, 13, e0007146. [Google Scholar] [CrossRef]
- Kumar, R.; Singh, O.P.; Gautam, S.; Nylen, S.; Sundar, S. Enhanced expression of Toll-like receptors 2 and 4, but not 9, in spleen tissue from patients with visceral leishmaniasis. Parasite Immunol. 2014, 36, 721–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babiker, D.T.; Bakhiet, S.M.; Mukhtar, M.M. Leishmania donovani influenced cytokines and Toll-like receptors expression among Sudanese visceral leishmaniasis patients. Parasite Immunol. 2015, 37, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Gupta, C.L.; Akhtar, S.; Waye, A.; Pandey, N.R.; Pathak, N.; Bajpai, P. Cross talk between Leishmania donovani CpG DNA and Toll-like receptor 9: An immunoinformatics approach. Biochem. Biophys. Res. Commun. 2015, 459, 424–429. [Google Scholar] [CrossRef]
- Bamigbola, I.E.; Ali, S. Paradoxical immune response in leishmaniasis: The role of toll-like receptors in disease progression. Parasite Immunol. 2022, 44, e12910. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Gupta, G.; Adhikari, A.; Majumder, S.; Kar Mahapatra, S.; Bhattacharyya Majumdar, S.; Majumdar, S. Miltefosine triggers a strong proinflammatory cytokine response during visceral leishmaniasis: Role of TLR4 and TLR9. Int. Immunopharmacol. 2012, 12, 565–572. [Google Scholar] [CrossRef]
- Ives, A.; Masina, S.; Castiglioni, P.; Prevel, F.; Revaz-Breton, M.; Hartley, M.A.; Launois, P.; Fasel, N.; Ronet, C. MyD88 and TLR9 dependent immune responses mediate resistance to Leishmania guyanensis infections, irrespective of Leishmania RNA virus burden. PLoS ONE 2014, 9, e96766. [Google Scholar] [CrossRef]
- Srivastava, S.; Pandey, S.P.; Jha, M.K.; Chandel, H.S.; Saha, B. Leishmania expressed lipophosphoglycan interacts with Toll-like receptor (TLR)-2 to decrease TLR-9 expression and reduce anti-leishmanial responses. Clin. Exp. Immunol. 2013, 172, 403–409. [Google Scholar] [CrossRef]
- Murray, H.W.; Zhang, Y.; Zhang, Y.; Raman, V.S.; Reed, S.G.; Ma, X. Regulatory actions of Toll-like receptor 2 (TLR2) and TLR4 in Leishmania donovani infection in the liver. Infect. Immun. 2013, 81, 2318–2326. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, R.K.; Gupta, C.L.; Bajpai, P. Impelling TLR9: Road to perspective vaccine for visceral leishmaniasis. Drug Dev. Res. 2022, 83, 222–224. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhattacharya, P.; Gannavaram, S.; Ismail, N.; Saxena, A.; Dagur, P.K.; Akue, A.; KuKuruga, M.; Nakhasi, H.L. Toll-like Receptor-9 (TLR-9) Signaling Is Crucial for Inducing Protective Immunity following Immunization with Genetically Modified Live Attenuated Leishmania Parasites. Pathogens 2023, 12, 534. https://doi.org/10.3390/pathogens12040534
Bhattacharya P, Gannavaram S, Ismail N, Saxena A, Dagur PK, Akue A, KuKuruga M, Nakhasi HL. Toll-like Receptor-9 (TLR-9) Signaling Is Crucial for Inducing Protective Immunity following Immunization with Genetically Modified Live Attenuated Leishmania Parasites. Pathogens. 2023; 12(4):534. https://doi.org/10.3390/pathogens12040534
Chicago/Turabian StyleBhattacharya, Parna, Sreenivas Gannavaram, Nevien Ismail, Ankit Saxena, Pradeep K. Dagur, Adovi Akue, Mark KuKuruga, and Hira L. Nakhasi. 2023. "Toll-like Receptor-9 (TLR-9) Signaling Is Crucial for Inducing Protective Immunity following Immunization with Genetically Modified Live Attenuated Leishmania Parasites" Pathogens 12, no. 4: 534. https://doi.org/10.3390/pathogens12040534
APA StyleBhattacharya, P., Gannavaram, S., Ismail, N., Saxena, A., Dagur, P. K., Akue, A., KuKuruga, M., & Nakhasi, H. L. (2023). Toll-like Receptor-9 (TLR-9) Signaling Is Crucial for Inducing Protective Immunity following Immunization with Genetically Modified Live Attenuated Leishmania Parasites. Pathogens, 12(4), 534. https://doi.org/10.3390/pathogens12040534