
Diversity and Phylogeny of Cattle Ixodid Ticks and Associated Spotted Fever Group Rickettsia spp. in Tunisia

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
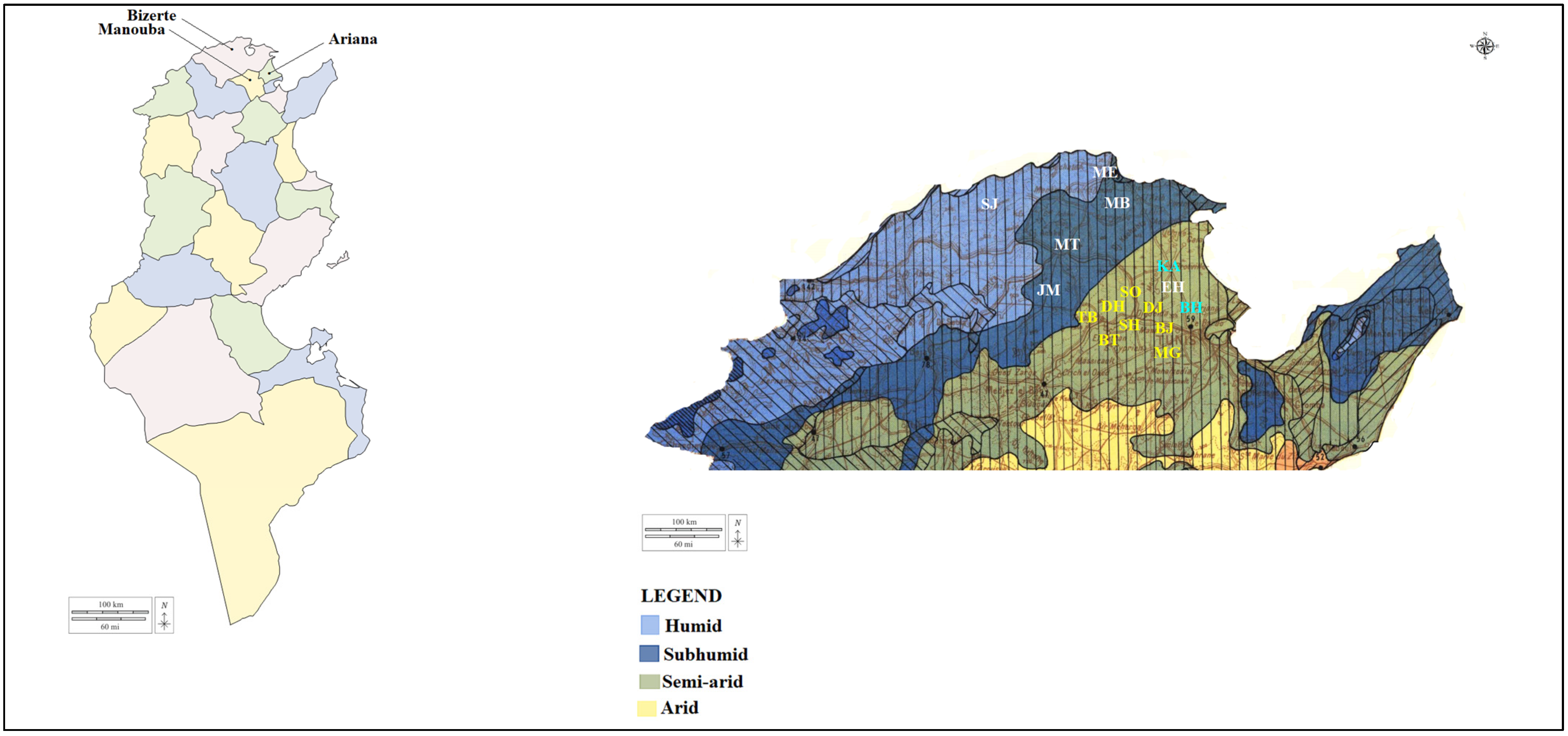
2.1. Study Regions, Tick Collection and Morphological Identification
2.2. Total DNA Extraction and Tick DNA Amplification
2.3. Molecular Detection of Rickettsia Species
2.4. Statistical Analysis
2.5. DNA Sequencing, Sequence Alignment and Phylogenetic Study
3. Results
3.1. Morphological and Molecular Identification of Ticks and Phylogenetic Analysis
3.1.1. Efficiency of DNA Isolation and Distribution of Collected Ticks
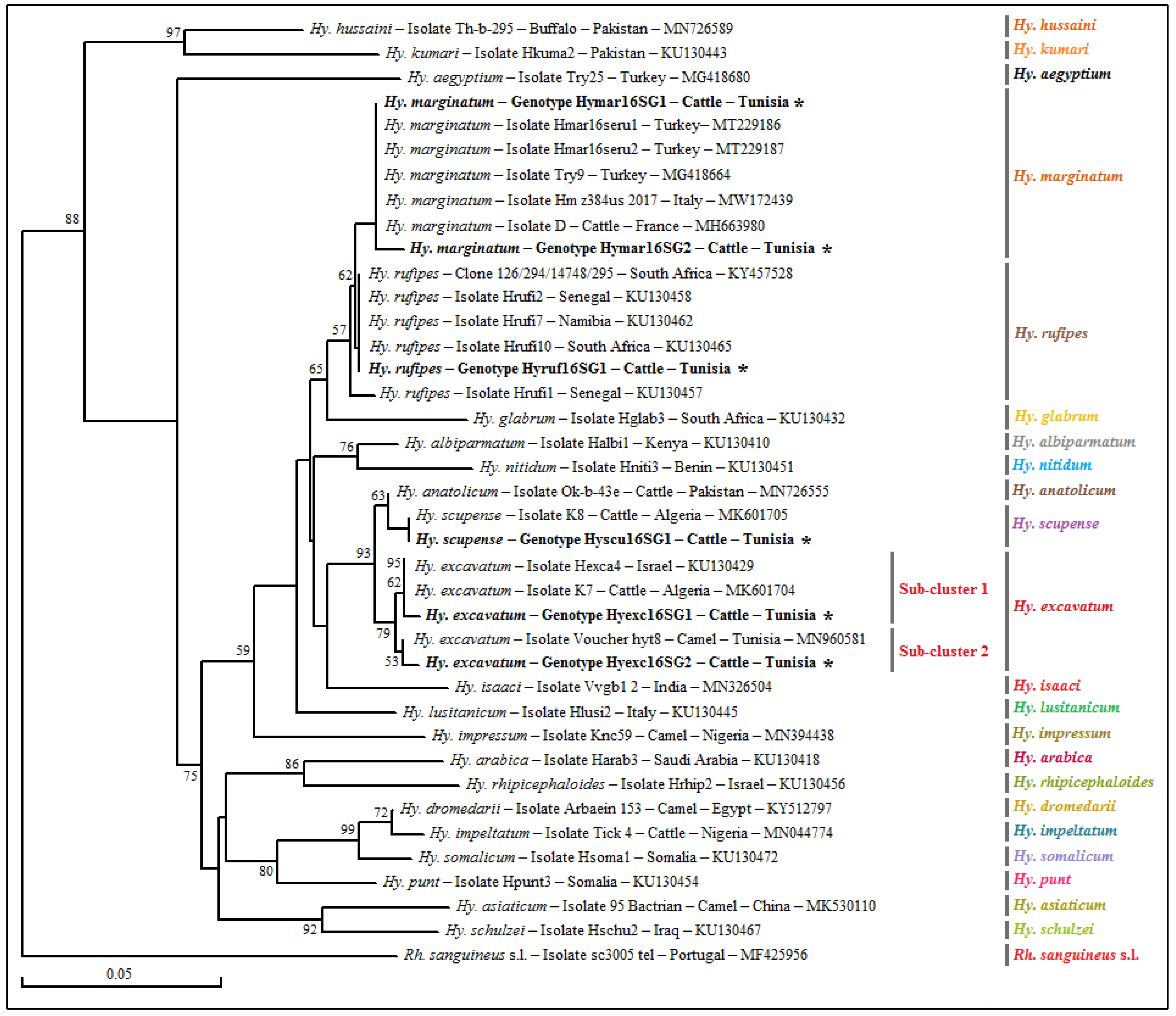
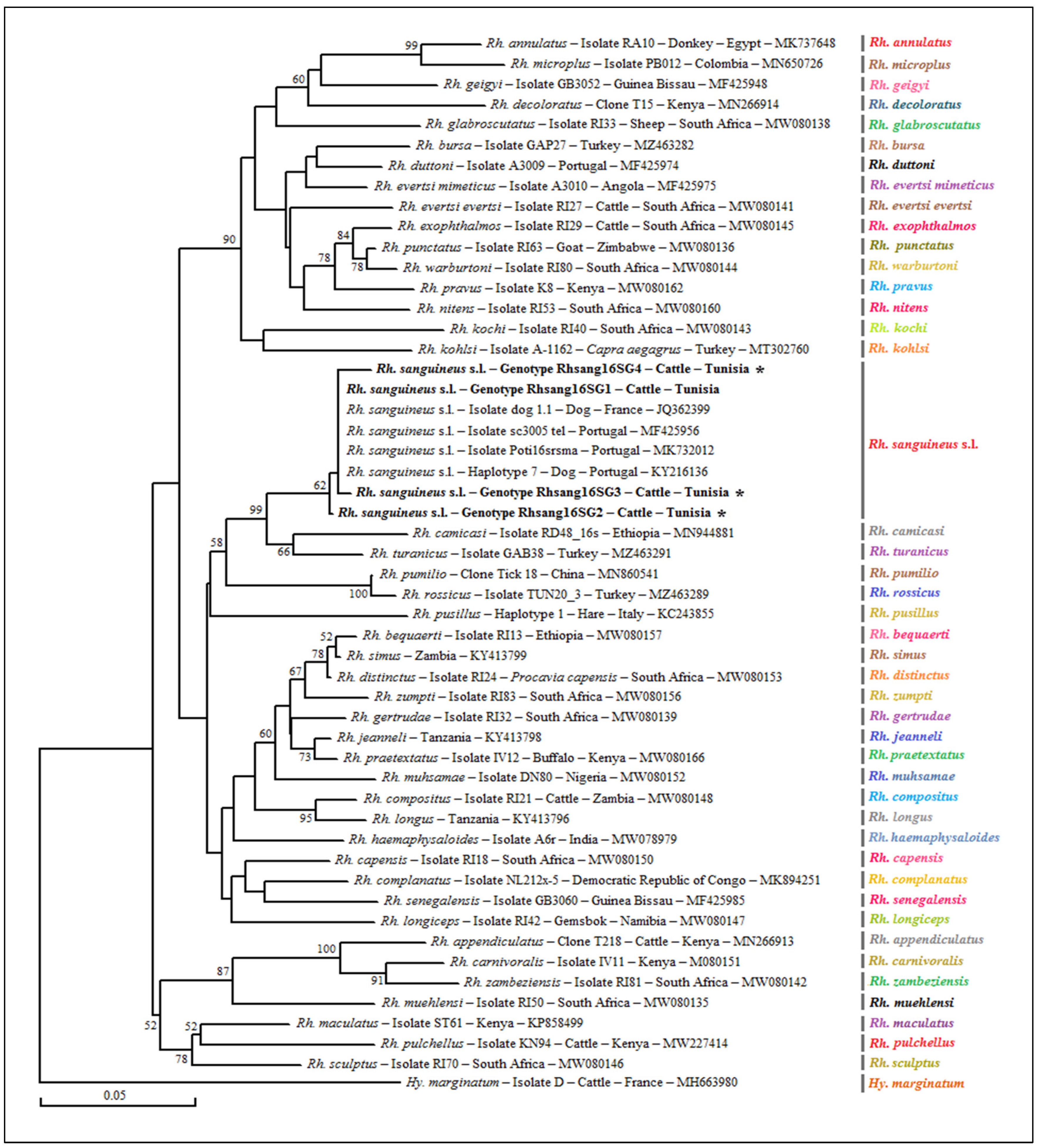
3.1.2. Genotyping and Phylogenetic Analysis of Selected Tick Specimens
3.2. Molecular Prevalence of Rickettsia spp.
3.3. Rickettsia Species Identification
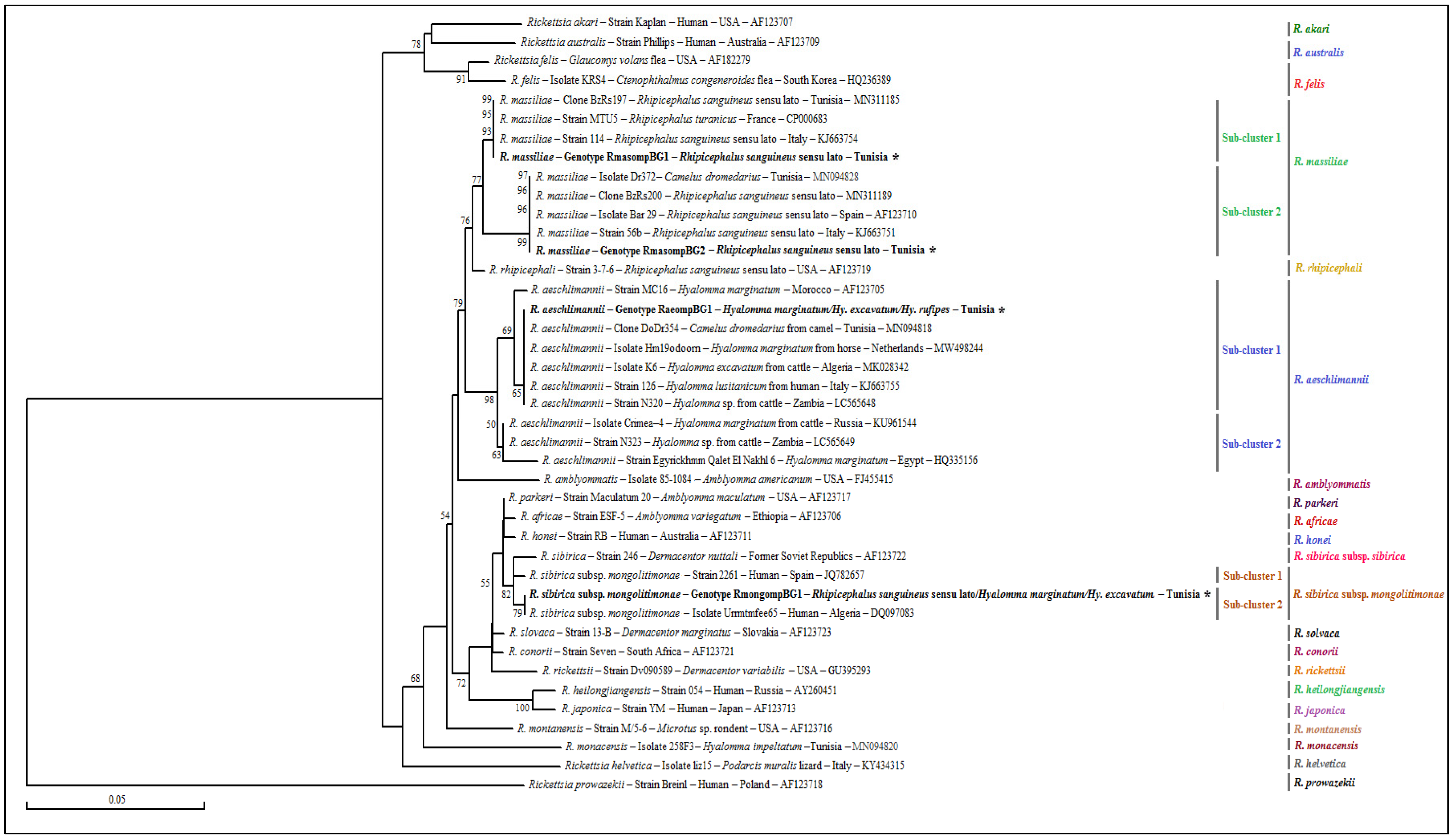
3.4. Genotyping and Phylogenetic Analysis
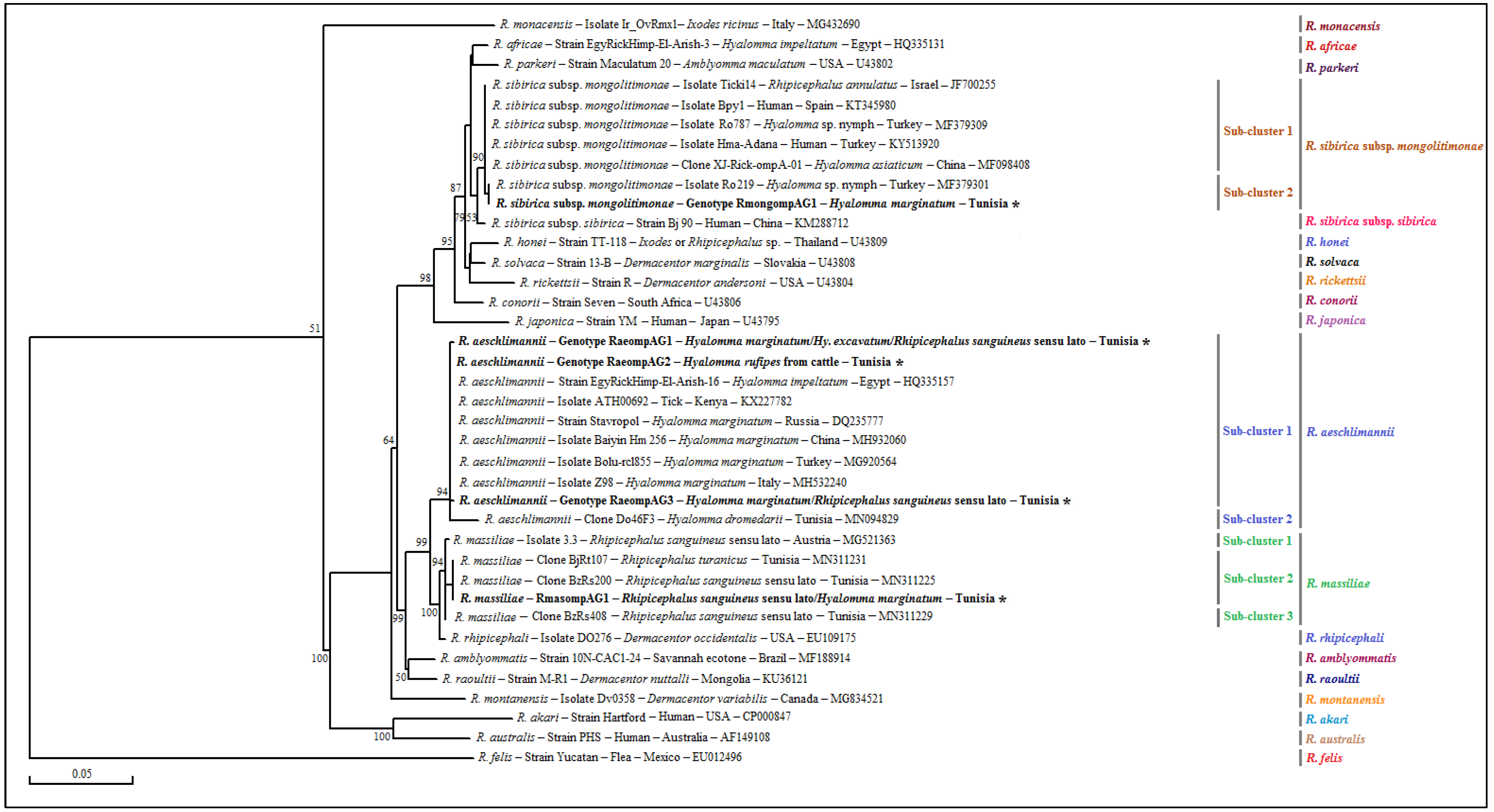
3.4.1. Rickettsia spp. ompB Partial Sequences
3.4.2. Rickettsia spp. ompA Partial Sequences
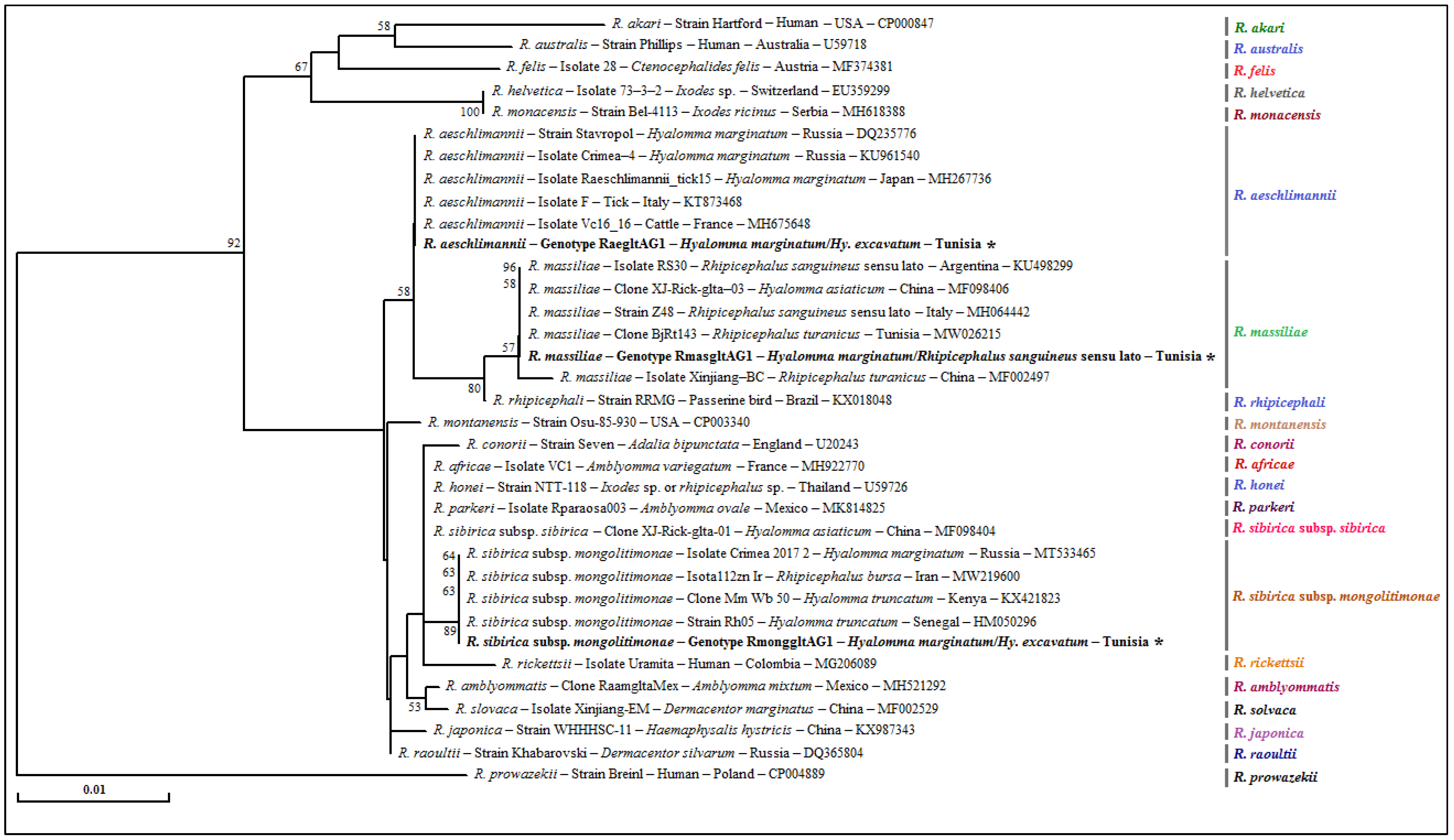
3.4.3. Rickettsia spp. gltA Partial Sequences
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garcia, K.; Weakley, M.; Do, T.; Mir, S. Current and Future Molecular Diagnostics of Tick-Borne Diseases in Cattle. Vet. Sci. 2022, 9, 241. [Google Scholar] [CrossRef] [PubMed]
- Kohli, S.; Atheya, U.K.; Thapliyal, A. Prevalence of Theileriosis in Cross-Bred Cattle: Its Detection through Blood Smear Examination and Polymerase Chain Reaction in Dehradun District, Uttarakhand, India. Vet. World 2014, 7, 168–171. [Google Scholar] [CrossRef] [Green Version]
- García, D.I.D.; Agatón, F.T.; Rosario-Cruz, R. Evaluación Económica Del Control de Garrapatas Rhipicephalus microplus En México/Economic Evaluation of Tick (Rhipicephalus microplus) Control in Mexico. CIBA Rev. Iberoam. Las Cienc. Biológicas Agropecu. 2016, 5, 43–52. [Google Scholar]
- Lagunes, R.L.; Domínguez-García, D.; Quiroz, H.; Martínez-Velázquez, M.; Rosario-Cruz, R. Potential Effects on Rhipicephalus microplus Tick Larvae Fed on Calves Immunized with a Subolesin Peptide Predicted by Epitope Analysis. Trop. Biomed. 2016, 33, 726–738. [Google Scholar] [PubMed]
- Hurtado, O.J.B.; Giraldo-Ríos, C. Economic and Health Impact of the Ticks in Production Animals. In Ticks and Tick-Borne Pathogens; Abubakar, M., Perera, P.K., Eds.; IntechOpen: London, UK, 2018. [Google Scholar]
- Merhej, V.; Raoult, D. Rickettsial Evolution in the Light of Comparative Genomics. Biol. Rev. 2010, 86, 379–405. [Google Scholar] [CrossRef]
- de la Fuente, J.; Antunes, S.; Bonnet, S.; Cabezas-Cruz, A.; Domingos, A.G.; Estrada-Peña, A.; Johnson, N.; Kocan, K.M.; Mansfield, K.L.; Nijhof, A.M.; et al. Tick-Pathogen Interactions and Vector Competence: Identification of Molecular Drivers for Tick-Borne Diseases. Front. Cell Infect. Microbiol. 2017, 7, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merhej, V.; Angelakis, E.; Socolovschi, C.; Raoult, D. Genotyping, Evolution and Epidemiological Findings of Rickettsia Species. Infect. Genet. Evol. 2014, 25, 122–137. [Google Scholar] [CrossRef]
- Alanazi, A.D.; Alanazi, A.D.; Nguyen, V.L.; Alyousif, M.S.; Manoj, R.R.S.; Alouffi, A.S.; Donato, R.; Sazmand, A.; Mendoza-Roldan, J.A.; Dantas-Torres, F.; et al. Ticks and Associated Pathogens in Camels (Camelus dromedarius) from Riyadh Province, Saudi Arabia. Parasit. Vectors 2020, 13, 110. [Google Scholar] [CrossRef] [Green Version]
- Azmat, M.; Ijaz, M.; Farooqi, S.H.; Ghaffar, A.; Ali, A.; Masud, A.; Saleem, S.; Rehman, A.; Ali, M.M.; Mehmood, K.; et al. Molecular Epidemiology, Associated Risk Factors, and Phylogenetic Analysis of Anaplasmosis in Camel. Microb. Pathog. 2018, 123, 377–384. [Google Scholar] [CrossRef]
- Ben Said, M.; Belkahia, H.; Alberti, A.; Abdi, K.; Zhioua, M.; Daaloul-Jedidi, M.; Messadi, L. First Molecular Evidence of Borrelia Burgdorferi Sensu Lato in Goats, Sheep, Cattle and Camels in Tunisia. Ann. Agric. Environ. Med. 2016, 23, 442–447. [Google Scholar] [CrossRef]
- Bahari, A.; Azami, S.; Goudarztalejerdi, A.; Karimi, S.; Esmaeili, S.; Chomel, B.B.; Sazmand, A. Molecular Detection of Zoonotic Pathogens in the Blood and Tissues of Camels (Camelus dromedarius) in Central Desert of Iran. Yale J. Biol. Med. 2021, 94, 249–258. [Google Scholar] [PubMed]
- Selmi, R.; Ben Said, M.; Ben Yahia, H.; Abdelaali, H.; Messadi, L. Molecular Epidemiology and Phylogeny of Spotted Fever Group Rickettsia in Camels (Camelus dromedarius) and Their Infesting Ticks from Tunisia. Transbound. Emerg. Dis. 2019, 67, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Larson, P.S.; Espira, L.; Grabow, C.; Wang, C.A.; Muloi, D.; Browne, A.S.; Deem, S.L.; Fèvre, E.M.; Foufopoulos, J.; Hardin, R.; et al. The Sero-Epidemiology of Coxiella burnetii (Q Fever) across Livestock Species and Herding Contexts in Laikipia County, Kenya. Zoonoses Public Health 2019, 66, 316–324. [Google Scholar] [CrossRef] [Green Version]
- Selmi, R.; Belkahia, H.; Sazmand, A.; Ben Said, M.; Messadi, L. Epidemiology and Genetic Characteristics of Tick-Borne Bacteria in Dromedary Camels of the World. Acta Trop. 2022, 234, 106599. [Google Scholar] [CrossRef]
- Ereqat, S.; Nasereddin, A.; Al-Jawabreh, A.; Azmi, K.; Harrus, S.; Mumcuoglu, K.; Apanaskevich, D.; Abdeen, Z. Molecular Detection and Identification of Spotted Fever Group Rickettsiae in Ticks Collected from the West Bank, Palestinian Territories. PLoS Negl. Trop. Dis. 2016, 10, e0004348. [Google Scholar] [CrossRef]
- Roux, V.; Rydkina, E.; Eremeeva, M.; Raoult, D. Citrate Synthase Gene Comparison, a New Tool for Phylogenetic Analysis, and Its Application for the Rickettsiae. Int. J. Syst. Bacteriol. 1997, 47, 252–261. [Google Scholar] [CrossRef] [Green Version]
- Znazen, A.; Khrouf, F.; Elleuch, N.; Lahiani, D.; Marrekchi, C.; M’Ghirbi, Y.; Ben Jemaa, M.; Bouattour, A.; Hammami, A. Multispacer Typing of Rickettsia Isolates from Humans and Ticks in Tunisia Revealing New Genotypes. Parasit. Vectors 2013, 6, 367. [Google Scholar] [CrossRef] [Green Version]
- Khrouf, F.; Sellami, H.; Elleuch, E.; Hattab, Z.; Ammari, L.; Khalfaoui, M.; Souissi, J.; Harrabi, H.; M’ghirbi, Y.; Tiouiri, H.; et al. Molecular Diagnosis of Rickettsia Infection in Patients from Tunisia. Ticks Tick Borne Dis. 2016, 7, 653–656. [Google Scholar] [CrossRef]
- Demoncheaux, J.-P.; Socolovschi, C.; Davoust, B.; Haddad, S.; Raoult, D.; Parola, P. First Detection of Rickettsia aeschlimannii in Hyalomma dromedarii Ticks from Tunisia. Ticks Tick Borne Dis. 2012, 3, 398–402. [Google Scholar] [CrossRef]
- Khrouf, F.; M’Ghirbi, Y.; Znazen, A.; Ben Jemaa, M.; Hammami, A.; Bouattour, A. Detection of Rickettsia in Rhipicephalus sanguineus Ticks and Ctenocephalides felis Fleas from Southeastern Tunisia by Reverse Line Blot Assay. J. Clin. Microbiol. 2013, 52, 268–274. [Google Scholar] [CrossRef] [Green Version]
- Belkahia, H.; Selmi, R.; Zamiti, S.; Daaloul-Jedidi, M.; Messadi, L.; Ben Said, M. Zoonotic Rickettsia Species in Small Ruminant Ticks from Tunisia. Front. Vet. Sci. 2021, 8, 676896. [Google Scholar] [CrossRef] [PubMed]
- Thrusfield, M. Veterinary Epidemiology; Blackwell Publishing Professional: Ames, IA, USA, 2005; Volume 47, ISBN 0-632-06397-1. [Google Scholar]
- Walker, A.R.; International Consortium on Ticks and Tick-Borne Diseases. Ticks of Domestic Animals in Africa: A Guide to Identification of Species; Bioscience Reports: Edinburgh, UK, 2003; ISBN 9780954517304. [Google Scholar]
- Black, W.C.; Piesman, J. Phylogeny of Hard- and Soft-Tick Taxa (Acari: Ixodida) Based on Mitochondrial 16S RDNA Sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 10034–10038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.-J.; Jang, W.-J.; Kim, J.-H.; Ryu, J.-S.; Lee, S.-H.; Park, K.-H.; Paik, H.-S.; Koh, Y.-S.; Choi, M.-S.; Kim, I.-S. Spotted Fever Group and Typhus Group Rickettsioses in Humans, South Korea. Emerg. Infect. Dis. 2005, 11, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Oteo, J.A.; Portillo, A.; Santibáñez, S.; Blanco, J.R.; Pérez-Martínez, L.; Ibarra, V. Cluster of Cases of Human Rickettsia felis Infection from Southern Europe (Spain) Diagnosed by PCR. J. Clin. Microbiol. 2006, 44, 2669–2671. [Google Scholar] [CrossRef] [Green Version]
- Regnery, R.L.; Olson, J.G.; Perkins, B.A.; Bibb, W. Serological Response to “Rochalimaea henselae” Antigen in Suspected Cat-Scratch Disease. Lancet 1992, 339, 1443–1445. [Google Scholar] [CrossRef]
- Pearson, W.R. An Introduction to Sequence Similarity (“Homology”) Searching. Curr. Protoc. Bioinform. 2013, 42, 3.1.1–3.1.8. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L. Assessing and Improving Methods Used in Operational Taxonomic Unit-Based Approaches for 16S RRNA Gene Sequence Analysis. Appl. Environ. Microbiol. 2011, 77, 3219–3226. [Google Scholar] [CrossRef] [Green Version]
- Desper, R.; Gascuel, O. Getting a Tree Fast: Neighbor Joining, FastME, and Distance-Based Methods. Curr. Protoc. Bioinform. 2006, 15, 6.3.1–6.3.28. [Google Scholar] [CrossRef] [PubMed]
- Mort, M.E.; Soltis, P.S.; Soltis, D.E.; Mabry, M.L. Comparison of Three Methods for Estimating Internal Support on Phylogenetic Trees. Syst. Biol. 2000, 49, 160–171. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Otranto, D. Best Practices for Preventing Vector-Borne Diseases in Dogs and Humans. Trends Parasitol. 2016, 32, 43–55. [Google Scholar] [CrossRef]
- Battisti, E.; Zanet, S.; Boraso, F.; Minniti, D.; Giacometti, M.; Duscher, G.G.; Ferroglio, E. Survey on Tick-Borne Pathogens in Ticks Removed from Humans in Northwestern Italy. Vet Parasitol. Reg. Stud. Rep. 2019, 18, 100352. [Google Scholar] [CrossRef] [PubMed]
- Vanegas, A.; Keller, C.; Krüger, A.; Manchang, T.K.; Hagen, R.M.; Frickmann, H.; Veit, A.; Achukwi, M.D.; Krücken, J.; Poppert, S. Molecular Detection of Spotted Fever Group Rickettsiae in Ticks from Cameroon. Ticks Tick Borne Dis. 2018, 9, 1049–1056. [Google Scholar] [CrossRef]
- Cicculli, V.; Capai, L.; Quilichini, Y.; Masse, S.; Fernández-Alvarez, A.; Minodier, L.; Bompard, P.; Charrel, R.; Falchi, A. Molecular Investigation of Tick-Borne Pathogens in Ixodid Ticks Infesting Domestic Animals (Cattle and Sheep) and Small Rodents (Black Rats) of Corsica, France. Ticks Tick Borne Dis. 2019, 10, 606–613. [Google Scholar] [CrossRef]
- Pesquera, C.; Portillo, A.; Palomar, A.M.; Oteo, J.A. Investigation of Tick-Borne Bacteria (Rickettsia spp., Anaplasma spp., Ehrlichia spp. and Borrelia spp.) in Ticks Collected from Andean Tapirs, Cattle and Vegetation from a Protected Area in Ecuador. Parasit. Vectors 2015, 8, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehlers, J.; Krüger, A.; Rakotondranary, S.J.; Ratovonamana, R.Y.; Poppert, S.; Ganzhorn, J.U.; Tappe, D. Molecular Detection of Rickettsia spp., Borrelia spp., Bartonella spp. And Yersinia pestis in Ectoparasites of Endemic and Domestic Animals in Southwest Madagascar. Acta Trop. 2020, 205, 105339. [Google Scholar] [CrossRef]
- Chisu, V.; Leulmi, H.; Masala, G.; Piredda, M.; Foxi, C.; Parola, P. Detection of Rickettsia hoogstraalii, Rickettsia helvetica, Rickettsia massiliae, Rickettsia slovaca and Rickettsia aeschlimannii in Ticks from Sardinia, Italy. Ticks Tick Borne Dis. 2017, 8, 347–352. [Google Scholar] [CrossRef]
- Chitanga, S.; Chibesa, K.; Sichibalo, K.; Mubemba, B.; Nalubamba, K.S.; Muleya, W.; Changula, K.; Simulundu, E. Molecular Detection and Characterization of Rickettsia Species in Ixodid Ticks Collected from Cattle in Southern Zambia. Front. Vet Sci. 2021, 8, 684487. [Google Scholar] [CrossRef]
- Azagi, T.; Klement, E.; Perlman, G.; Lustig, Y.; Mumcuoglu, K.Y.; Apanaskevich, D.A.; Gottlieb, Y. Francisella-like Endosymbionts and Rickettsia Species in Local and Imported Hyalomma Ticks. Appl. Environ. Microbiol. 2017, 83, 18. [Google Scholar] [CrossRef] [Green Version]
- Kumsa, M.K.; Chambon, A.; Yan, M.C.; Maiter, S. Catching the Shimmers of the Social: From the Limits of Reflexivity to Methodological Creativity. Qual. Res. J. 2014, 15, 419–436. [Google Scholar] [CrossRef]
- Leulmi, H.; Aouadi, A.; Bitam, I.; Bessas, A.; Benakhla, A.; Raoult, D.; Parola, P. Detection of Bartonella tamiae, Coxiella burnetii and Rickettsiae in Arthropods and Tissues from Wild and Domestic Animals in Northeastern Algeria. Parasit. Vectors 2016, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.-E.; et al. Update on Tick-Borne Rickettsioses around the World: A Geographic Approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, P.-E. Evidence of Infection in Humans with Rickettsia helvetica in Eastern France. Emerg. Infect. Dis. 2000, 6, 389–392. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, A.-M.; Birtles, R.J. Rickettsia mongolotimonae Infection in South Africa. Emerg. Infect. Dis. 2004, 10, 126–129. [Google Scholar] [CrossRef]
- Germanakis, A.; Chochlakis, D.; Angelakis, E.; Tselentis, Y.; Psaroulaki, A. Skin Lesions and Inoculation Eschars at the Tick Bite Site in Spotted Fever Group Rickettsioses: Experience from a Patient Series in Eastern Crete, Greece. Dermatology 2014, 228, 332–337. [Google Scholar] [CrossRef]
- Aguirrebengoa, K.; Portillo, A.; Santibáñez, S.; Marín, J.J.; Montejo, M.; Oteo, J.A.; Aguirrebengoa, K.; Portillo, A.; Santibáñez, S.; Marín, J.J.; et al. Human Rickettsia sibirica mongolitimonae Infection, Spain. Emerg. Infect. Dis. 2008, 14, 528–529. [Google Scholar] [CrossRef]
- Chochlakis, D.; Ioannou, I.; Sandalakis, V.; Dimitriou, T.; Kassinis, N.; Papadopoulos, B.; Tselentis, Y.; Psaroulaki, A. Spotted Fever Group Rickettsiae in Ticks in Cyprus. Microb. Ecol. 2011, 63, 314–323. [Google Scholar] [CrossRef]
- Boulkaboul, A. Parasitisme Des Tiques (Ixodidae) Des Bovins à Tiaret, Algérie. Rev. D’élevage Médecine Vétérinaire Pays Trop. 2003, 56, 157. [Google Scholar] [CrossRef] [Green Version]
- Awa, D.N.; Adakal, H.; Luogbou, N.D.D.; Wachong, K.H.; Leinyuy, I.; Achukwi, M.D. Cattle Ticks in Cameroon: Is Rhipicephalus (Boophilus) microplus Absent in Cameroon and the Central African Region? Ticks Tick Borne Dis. 2015, 6, 117–122. [Google Scholar] [CrossRef]
- Chisu, V.; Foxi, C.; Mannu, R.; Satta, G.; Masala, G. A Five-Year Survey of Tick Species and Identification of Tick-Borne Bacteria in Sardinia, Italy. Ticks Tick Borne Dis. 2018, 9, 678–681. [Google Scholar] [CrossRef]
- Fournier, P.-E.; Zhu, Y.; Yu, X.; Raoult, D. Proposal to Create Subspecies of Rickettsia sibirica and an Emended Description of Rickettsia sibirica. Ann. N. Y. Acad. Sci. 2006, 1078, 597–606. [Google Scholar] [CrossRef]
- Orkun, Ö.; Çakmak, A. Molecular Identification of Tick-Borne Bacteria in Wild Animals and Their Ticks in Central Anatolia, Turkey. Comp. Immunol. Microbiol. Infect. Dis. 2019, 63, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Alieva, E.E.; Bondarenko, E.I.; Maliy, K.D.; Shvalov, A.N.; Verbenets, E.A.; Gafarova, M.T. The Role of Rhipicephalus sanguineus Ticks Parasitizing Dogs in the Spread of Tick-Borne Rickettsial Pathogens in the City of Sevastopol. New. Microbes. New. Infect. 2020, 36, 100704. [Google Scholar] [CrossRef] [PubMed]
- Kernif, T.; Socolovschi, C.; Bitam, I.; Raoult, D.; Parola, P. Vector-Borne Rickettsioses in North Africa. Infect Dis Clin N. Am. 2012, 26, 455–478. [Google Scholar] [CrossRef] [PubMed]
- Chisu, V.; Masala, G.; Foxi, C.; Socolovschi, C.; Raoult, D.; Parola, P.; Chisu, V.; Masala, G.; Foxi, C.; Socolovschi, C.; et al. Rickettsia conorii israelensis in Rhipicephalus sanguineus Ticks, Sardinia, Italy. Ticks Tick Borne Dis. 2014, 5, 446–448. [Google Scholar] [CrossRef]
- Blanc, G.; Ogata, H.; Robert, C.; Audic, S.; Claverie, J.M.; Raoult, D.; Blanc, G.; Ogata, H.; Robert, C.; Audic, S.; et al. Lateral Gene Transfer between Obligate Intracellular Bacteria: Evidence from the Rickettsia massiliae Genome. Genome. Res. 2007, 17, 1657–1664. [Google Scholar] [CrossRef] [Green Version]
- Roux, V.; Raoult, D. Phylogenetic Analysis of Members of the Genus Rickettsia Using the Gene Encoding the Outer-Membrane Protein ROmpB (OmpB). Int. J. Syst. Evol. Microbiol. 2000, 50, 1449–1455. [Google Scholar] [CrossRef] [Green Version]
- Abdelkadir, K.; Palomar, A.M.; Portillo, A.; Oteo, J.A.; Ait-Oudhia, K.; Khelef, D. Presence of Rickettsia aeschlimannii, “Candidatus Rickettsia Barbariae” and Coxiella burnetii in Ticks from Livestock in Northwestern Algeria. Ticks Tick Borne Dis. 2019, 10, 924–928. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assays (Reference) | Target Genes | Primers | Sequences (5′-3′) | Amplicon Size (bp) |
|---|---|---|---|---|
| Single PCR 1 [25] | ||||
| 16S rRNA | TQ16S+1F | CTGCTCAATGATTTTTTAAATTGCTGTGG | 324 | |
| TQ16S-2R | ACGCTGTTATCCCTAGAG | |||
| Nested PCR 2 [26] | ||||
| First PCR | ompB | rompB_OF | GTAACCGGAAGTAATCGTTTCGTAA | 511 |
| rompB OR | GCTTTATAACCAGCTAAACCACC | |||
| Second PCR | rompB_SFG_IF | GTTTAATACGTGCTGCTAACCAA | 425 | |
| rompB SFG-IR | GGTTTGGCCCATATACCATAAG | |||
| Semi-nested PCR 3 [27] | ||||
| First PCR | ompA | Rr190.70p | ATGGCGAATATTTCTCCAAAA | 631 |
| Rr190.701n | GTTCCGTTAATGGCAGCATCT | |||
| Second PCR | Rr190.70p | ATGGCGAATATTTCTCCAAAA | 532 | |
| Rr190.602n | AGTGCAGCATTCGCTCCCCCT | |||
| Single PCR 3 [28] | ||||
| gltA | RpCS.877p | GGGGGCCTGCTCACGGCGG | 381 | |
| RpCS.1258n | ATTGCAAAAAGTACAGTGAACA |
| Factors | Number (%) | Positive (% ± C.I. 1) | p-Value (Chi2) |
|---|---|---|---|
| Tick species | |||
| Hyalommaexcavatum | 129 (38.2) | 26 (20.1 ± 0.06) | 0.022 * (11.39) |
| Rhipicephalus sanguineus sensu lato | 111 (32.8) | 38 (34.2 ± 0.08) | |
| Hyalommamarginatum | 84 (24.9) | 25 (29.8 ± 0.09) | |
| Hyalommascupense | 12 (3.6) | 0 (0) | |
| Hyalommarufipes | 2 (0.6) | 1 (50.0 ± 0.69) | |
| Tick gender | |||
| Male | 268 (79.3) | 67 (25.0 ± 0.05) | 0.186 (1.75) |
| Female | 70 (20.7) | 23 (32.8 ± 0.10) | |
| Bioclimatic area | |||
| Subhumid | 197 (58.3) | 45 (22.8 ± 0.05) | 0.063 (3.45) |
| Higher semi-arid | 141 (41.7) | 45 (31.9 ± 0.07) | |
| Governorate | |||
| Bizerte | 197 (58.3) | 45 (22.8 ± 0.05) | 0.157 (3.70) |
| Manouba | 116 (34.3) | 38 (32.7 ± 0.08) | |
| Ariana | 25 (7.4) | 7 (28.0 ± 0.17) | |
| District | |||
| Menzel Bourguiba | 113 | 28 (24.7 ± 0.08) | 0.696 (0.152) |
| Sidi Othman | 39 | 11 (28.2 ± 0.14) | |
| Mornaguia | 35 | 10 (28.5 ± 0.15) | |
| Tebourba | 31 | 13 (41.9 ± 0.17) | |
| Kalaât El Andalous | 20 | 4 (20.0 ± 0.17) | |
| Djedeida | 19 | 6 (21.5 ± 0.21) | |
| Bach Hamba | 16 | 0 (0) | |
| Dhniba | 14 | 4 (28.5 ± 0.24) | |
| Battan | 12 | 3 (25.0 ± 0.26) | |
| Mabtouh | 10 | 1 (10 ± 0.19) | |
| Sejnane | 9 | 1 (11.1 ± 0.2) | |
| Metline | 6 | 4 (66.66 ± 0.38) | |
| Bjaoua | 5 | 3 (60.0 ± 0.43) | |
| Sanhaja | 5 | 2 (40.0 ± 0.43) | |
| Joumine | 3 | 0 (0) | |
| Mateur | 1 | 0 (0) | |
| Total | 338 | 90 (26.6 ± 0.05) |
| Sample (District) | Morp. Id. | BLAST 1 (GenBank 2, Genotype) | BLAST 3 (GenBank 2, Genotype) | ||
|---|---|---|---|---|---|
| ompB | ompA | gltA | |||
| Hyma83 (Tebourba) | Hy. sp. | 100% Hy. marg (OQ109189, Hymar16SG1) | 100% R. aesch (OQ123608, RaeompBG1) | 100% R. aesch (OQ123655, RaeompAG1) | - |
| Hyma161 (Jdaida) | Hy. marg | 100% Hy. marg (OQ109190, Hymar16SG1) | 100% R. aesch (OQ123609, RaeompBG1) | - | - |
| Hyma173 (Mornaguia) | Hy. marg | 100% Hy. marg (OQ109191, Hymar16SG1) | 100% R. aesch (OQ123610, RaeompBG1) | - | 100% R. aesch (OQ123685, RaegltAG1) |
| Hyma151 (Sidi Othmen) | Hy. marg | 100% Hy. marg (OQ109192, Hymar16SG1) | 100% R. aesch (OQ123611, RaeompBG1) | - | - |
| Hyma108 (Dhniba) | Hy. marg | 100% Hy. marg (OQ109193, Hymar16SG1) | 100% R. aesch (OQ123612, RaeompBG1) | - | 100% R. aesch (OQ123686, RaegltAG1) |
| Hyma109 (Dhniba) | Hy. marg | 100% Hy. marg (OQ109194, Hymar16SG1) | 100% R. aesch (OQ123613, RaeompBG1) | - | - |
| Hyma334 (Metline) | Hy. marg | 100% Hy. marg (OQ109195, Hymar16SG1) | 100% R. aesch (OQ123614, RaeompBG1) | - | - |
| Hyma113 (Dhniba) | Hy. marg | 100% Hy. marg (OQ109196, Hymar16SG1) | 100% R. aesch (OQ123615, RaeompBG1) | 100% R. aesch (OQ123656, RaeompAG1) | - |
| Hyma333 (Metline) | Hy. marg | - | 100% R. aesch (OQ123616, RaeompBG1) | - | - |
| Hyma140 (Sidi Othmen) | Hy. marg | 100% Hy. marg (OQ109197, Hymar16SG1) | 100% R. aesch (OQ123617, RaeompBG1) | - | 100% R. aesch (OQ123687, RaegltAG1) |
| Hyma96 (Jdaida) | Hy. marg | 100% Hy. marg (OQ109198, Hymar16SG1) | 100% R. aesch (OQ123618, RaeompBG1) | - | 100% R. aesch (OQ123688, RaegltAG1) |
| Hyma209 (Jdaida) | Hy. marg | 100% Hy. marg (OQ109199, Hymar16SG1) | - | 100% R. aesch (OQ123657, RaeompAG1) | - |
| Hyma68 (Battan) | Hy. marg | 99.6% Hy. marg (OQ109200, Hymar16SG2) | - | 100% R. aesch (OQ123658, RaeompAG1) | - |
| Hyma336 (Metline) | Hy. marg | 100% Hy. marg (OQ109201, Hymar16SG1) | - | 100% R. mas (OQ123676, RmasompAG1) | - |
| Hyma25 (Sejnane) | Hy. marg | - | - | 100% R. aesch (OQ123659, RaeompAG1) | - |
| Hyma5 (K. El Andalous) | Hy. marg | 100% Hy. marg (OQ109202, Hymar16SG1) | - | 100% R. aesch (OQ123660, RaeompAG1) | - |
| Hyma174 (Mornaguia) | Hy. marg | 100% Hy. marg (OQ109203, Hymar16SG1) | - | 99.8% R. aesch (OQ123661, RaeompAG2) | - |
| Hyma72 (Tebourba) | Hy. marg | 100% Hy. marg (OQ109204, Hymar16SG1) | - | - | 100% R. mas (OQ123696, RmasgltAG1) |
| Hyma198 (Tebourba) | Hy. sp. | 100% Hy. marg (OQ109205, Hymar16SG1) | 100% R. aesch (OQ123619, RaeompBG1) | - | 100% R. aesch (OQ123689, RaegltAG1) |
| Hyma226 (Tebourba) | Hy. sp. | 100% Hy. marg (OQ109206, Hymar16SG1) | 100% R. sib subsp. mong (OQ123639, RmongompBG1) | 100% R. sib subsp. mong (OQ123675, RmongompAG1) | 100% R. sib subsp. mong (OQ123694, RmonggltAG1) |
| Hyma156 (Sidi Othmen) | Hy. marg | - | 100% R. aesch (OQ123620, RaeompBG1) | - | - |
| Gene | Tick or Rickettsia Species | Size (pb) | N | VS | GC% | G | Gd | Pi | k |
|---|---|---|---|---|---|---|---|---|---|
| Mito 16S rRNA | Hy. scupense | 273 | 12 | 0 | 48.7 | 1 | 0 | 0 | 0 |
| Hy. marginatum | 272 | 24 | 1 | 47.8 | 2 | 0.159 | 0.00059 | 0.159 | |
| Hy. rufipes | 272 | 1 | 0 | 47.8 | 1 | 0 | 0 | 0 | |
| Rh. sanguineus s.l. | 272 | 15 | 3 | 48.5 | 4 | 0.467 | 0.00189 | 0.514 | |
| Hy. excavatum | 270 | 31 | 3 | 49.3 | 2 | 0.452 | 0.00502 | 1.355 | |
| ompB | R. massiliae | 382 | 13 | 5 | 51.3 | 2 | 0.385 | 0.00503 | 1.923 |
| R. aeschlimannii | 382 | 31 | 0 | 51.3 | 1 | 0 | 0 | 0 | |
| R. sibirica subsp. mongolitimonae | 382 | 3 | 0 | 51.3 | 1 | 0 | 0 | 0 | |
| ompA | R. massiliae | 490 | 9 | 0 | 54.0 | 1 | 0 | 0 | 0 |
| R. aeschlimannii | 491 | 20 | 2 | 53.8 | 3 | 0.542 | 0.00123 | 0.605 | |
| R. sibirica subsp. mongolitimonae | 490 | 1 | 0 | 53.3 | 1 | 0 | 0 | 0 | |
| gltA | R. massiliae | 341 | 16 | 0 | 49.0 | 1 | 0 | 0 | 0 |
| R. aeschlimannii | 341 | 9 | 0 | 49.2 | 1 | 0 | 0 | 0 | |
| R. sibirica subsp. mongolitimonae | 341 | 2 | 0 | 48.4 | 1 | 0 | 0 | 0 |
| Sample (District) | Morp. Id. | BLAST 1 (GenBank 2, Genotype) | BLAST 3 (GenBank 2, Genotype) | ||
|---|---|---|---|---|---|
| ompB | ompA | gltA | |||
| Hyex143 (Sidi Othmen) | Hy. sp. | 99.2% Hy. exc (OQ109213, Hyexc16SG1) | - | 99.8% R. aesch (OQ123662, RaeompAG2) | - |
| Hyex206 (Sanhaja) | Hy. sp. | 99.2% Hy. exc (OQ109214, Hyexc16SG1) | 100% R. aesch (OQ123621, RaeompBG1) | 99.8% R. aesch (OQ123663, Raeomp AG2) | - |
| Hyex90 (Battan) | Hy. exc | 99.6% Hy. exc (OQ109215, Hyexc16SG2) | 100% R. aesch (OQ123622, RaeompBG1) | - | - |
| Hyex167 (Mornaguia) | Hy. exc | 99.6% Hy. exc (OQ109216, Hyexc16SG2) | 100% R. aesch (OQ123623, RaeompBG1) | - | - |
| Hyex141 (Sidi Othmen) | Hy. exc | 99.2% Hy. exc (OQ109217, Hyexc16SG1) | 100% R. aesch (OQ123624, RaeompBG1) | - | - |
| Hyex115 (Tebourba) | Hy. exc | 99.2% Hy. exc (OQ109218, Hyexc16SG1) | 100% R. aesch (OQ123625, RaeompBG1) | - | - |
| Hyex16 (K. El Andalous) | Hy. exc | 99.6% Hy. exc (OQ109219, Hyexc16SG2) | 100% R. aesch (OQ123626, RaeompBG1) | - | 100% R. aesch (OQ123690, RaegltAG1) |
| Hyex48 (M. Bourguiba) | Hy. exc | 99.6% Hy. exc (OQ109220, Hyexc16SG2) | 100% R. aesch (OQ123627, RaeompBG1) | - | 100% R. aesch (OQ123691, RaegltAG1) |
| Hyex78 (Battan) | Hy. exc | 99.6% Hy. exc (OQ109221, Hyexc16SG2) | 100% R. aesch (OQ123628, RaeompBG1) | - | 100% R. aesch (OQ123692, RaegltAG1) |
| Hyex171 (Mornaguia) | Hy. exc | 99.6% Hy. exc (OQ109222, Hyexc16SG2) | 100% R. aesch (OQ123629, RaeompBG1) | - | - |
| Hyex250 (M. Bourguiba) | Hy. exc | 99.2% Hy. exc (OQ109223, Hyexc16SG1) | 100% R. aesch (OQ123630, RaeompBG1) | 99.8% R. aesch (OQ123664, RaeompAG2) | - |
| Hyex237 (Sidi Othmen) | Hy. exc | 99.2% Hy. exc (OQ109224, Hyexc16SG1) | 100% R. sib subsp. mong (OQ123640, RmongompBG1) | - | 100% R. sib subsp. mong (OQ123695, RmonggltAG1) |
| Hyex129 (Sidi Othmen) | Hy. exc | 99.2% Hy. exc (OQ109225, Hyexc16SG1) | 100% R. aesch (OQ123631, RaeompBG1) | - | - |
| Hyex211 (Jdaida) | Hy. exc | 99.2% Hy. exc (OQ109226, Hyexc16SG1) | 100% R. aesch (OQ123632, RaeompBG1) | 99.8% R. aesch (OQ123665, RaeompAG2) | - |
| Hyex195 (Mornaguia) | Hy. exc | 99.2% Hy. exc (OQ109227, Hyexc16SG1) | 100% R. aesch (OQ123633, RaeompBG1) | 99.8% R. aesch (OQ123666, RaeompAG2) | 100% R. aesch (OQ123693, RaegltAG1) |
| Hyex175 (Mornaguia) | Hy. exc | 99.2% Hy. exc (OQ109228, Hyexc16SG1) | 100% R. aesch (OQ123634, RaeompBG1) | 99.8% R. aesch (OQ123667, RaeompAG2) | - |
| Hyex188 (Mornaguia) | Hy. exc | 99.2% Hy. exc (OQ109229, Hyexc16SG1) | 100% R. aesch (OQ123635, RaeompBG1) | 99.8% R. aesch (OQ123668, RaeompAG2) | - |
| Hyex164 (Mornaguia) | Hy. exc | 99.2% Hy. exc (OQ109230, Hyexc16SG1) | - | 99.8% R. aesch (OQ123669, RaeompAG2) | - |
| Hyex177 (Mornaguia) | Hy. exc | 99.2% Hy. exc (OQ109231, Hyexc16SG1) | 100% R. aesch (OQ123636, RaeompBG1) | 99.8% R. aesch (OQ123670, RaeompAG2) | - |
| Hyex170 (Mornaguia) | Hy. exc | 99.2% Hy. exc (OQ109232, Hyexc16SG1) | - | 99.8% R. aesch (OQ123671, RaeompAG2) | - |
| Hyex148 (Sidi Othmen) | Hy. exc | 99.2% Hy. exc (OQ109233, Hyexc16SG1) | 100% R. aesch (OQ123637, RaeompBG1) | - | - |
| Hyru97 (Jdaida) | Hy. sp. | 100% Hy. ruf (OQ109244, Hyruf16SG1) | 100% R. aesch (OQ123638, RaeompBG1) | 100% R. aesch (OQ123672, RaeompAG3) | - |
| Sample | Morp. Id. | BLAST 1 (GenBank 2, Genotype) | BLAST 3 (GenBank 2, Genotype) | ||
|---|---|---|---|---|---|
| ompB | ompA | gltA | |||
| Rhsa275 (M. Bourguiba) | Rh. sang s.l. | 100% Rh. sang s.l. (OQ109257, Rhsang16SG1) | 100% R. mas (OQ123642, RmasompBG1) | 100% R. mas (OQ123677, RmasompAG1) | 100% R. mas (OQ123697, RmasgltAG1) |
| Rhsa282 (M. Bourguiba) | Rh. sang s.l. | 100% Rh. sang s.l. (OQ109258, Rhsang16SG1) | 100% R. mas (OQ123643, RmasompBG2) | - | 100% R. mas (OQ123698, RmasgltAG1) |
| Rhsa77 (Bjaoua) | Rh. sang s.l. | 100% Rh. sang s.l. (OQ109259, Rhsang16SG1) | 100% R. mas (OQ123644, RmasompBG2) | 100% R. mas (OQ123678, RmasompAG1) | 100% R. mas (OQ123699, RmasgltAG1) |
| Rhsa1 (K. El Andalous) | Rh. sang s.l. | 99.6% Rh. sang s.l. (OQ109260, Rhsang16SG2) | 100% R. mas (OQ123645, RmasompBG2) | 100% R. mas (OQ123679, RmasompAG1) | 100% R. mas (OQ123700, RmasgltAG1) |
| Rhsa73 (Bjaoua) | Rh. sang s.l. | 99.6% Rh. sang s.l. (OQ109261, Rhsang16SG3) | 100% R. mas (OQ123646, RmasompBG1) | 99.8% R. aesch (OQ123673, RaeompAG2) | 100% R. mas (OQ123701, RmasgltAG1) |
| Rhsa9 (K. El Andalous) | Rh. sang s.l. | 100% Rh. sang s.l. (OQ109262, Rhsang16SG1) | 100% R. mas (OQ123647, RmasompBG2) | - | 100% R. mas (OQ123702, RmasgltAG1) |
| Rhsa284 (Mornaguia) | Rh. sang s.l. | 99.6% Rh. sang s.l. (OQ109263, Rhsang16SG4) | 100% R. sib subsp. mong (OQ123641, RmongompBG1) | 100% R. mas (OQ123680, RmasompAG1) | 100% R. mas (OQ123703, RmasgltAG1) |
| Rhsa252 (Sanhaja) | Rh. sang s.l. | 100% Rh. sang s.l. (OQ109264, Rhsang16SG1) | 100% R. mas (OQ123648, RmasompBG1) | - | 100% R. mas (OQ123704, RmasgltAG1) |
| Rhsa273 (M. Bourguiba) | Rh. sang s.l. | 99.6% Rh. sang s.l. (OQ109265, Rhsang16SG3) | 100% R. mas (OQ123649, RmasompBG2) | 99.8% R. aesch (OQ123674, RaesompAG1) | 100% R. mas (OQ123705, RmasgltAG1) |
| Rhsa203 (Tebourba) | Rh. sang s.l. | 100% Rh. sang s.l. (OQ109266, Rhsang16SG1) | - | 100% R. mas (OQ123681, RmasompAG1) | 100% R. mas (OQ123706, RmasgltAG1) |
| Rhsa122 (Tebourba) | Rh. sang s.l. | - | - | 100% R. mas (OQ123682, RmasompAG1) | - |
| Rhsa121 (Tebourba) | Rh. sang s.l. | - | - | 100% R. mas (OQ123683, RmasompAG1) | 100% R. mas (OQ123707, RmasgltAG1) |
| Rhsa254 (Sanhaja) | Rh. sang s.l. | - | - | 100% R. mas (OQ123684, RmasompAG1) | - |
| Rhsa119 (Tebourba) | Rh. sang s.l. | - | - | - | 100% R. mas (OQ123708, RmasgltAG1) |
| Rhsa120 (Tebourba) | Rh. sang s.l. | - | - | - | 100% R. mas (OQ123709, RmasgltAG1) |
| Rhsa57 (M. Bourguiba) | Rh. sang s.l. | - | - | - | 100% R. mas (OQ123710, RmasgltAG1) |
| Rhsa268 (Mornaguia) | Rh. sp. | 100% Rh. sang s. l. (OQ109267, Rhsang16SG1) | 100% R. mas (OQ123650, RmasompBG2) | - | - |
| Rhsa261 (M. Bourguiba) | Rh. sang s.l. | 100% Rh. sang s.l. (OQ109268, Rhsang16SG1) | - | - | 100% R. mas (OQ123711, RmasgltAG1) |
| Rhsa303 (M. Bourguiba) | Rh. sang s.l. | - | 100% R. mas (OQ123651, RmasompBG2) | - | - |
| Rhsa293 (M. Bourguiba) | Rh. sang s.l. | - | 100% R. mas (OQ123652, RmasompBG2) | - | - |
| Rhsa322 (M. Bourguiba) | Rh. sang s.l. | - | 100% R. mas (OQ123653, RmasompBG2) | - | - |
| Rhsa327 (M. Bourguiba) | Rh. sang s.l. | - | 100% R. mas (OQ123654, RmasompBG2) | - | - |
| Tick Species | ompB PCR Positive/ Sequenced | ompA PCR Positive/Sequenced | gltA PCR Positive/Sequenced | Rickettsia spp. |
|---|---|---|---|---|
| Hyalomma excavatum | 17 | 10 | 4 | R. aeschlimannii |
| 0 | 0 | 0 | R. massiliae | |
| 1 | 0 | 1 | R. sibirica subsp. mongolitimonae | |
| Hyalomma marginatum | 13 | 7 | 5 | R. aeschlimannii |
| 0 | 1 | 1 | R. massiliae | |
| 1 | 1 | 1 | R. sibirica subsp. mongolitimonae | |
| Rhipicephalus sanguineus sensu lato | 0 | 2 | 0 | R. aeschlimannii |
| 13 | 8 | 15 | R. massiliae | |
| 1 | 0 | 0 | R. sibirica subsp. mongolitimonae | |
| Hyalomma rufipes | 1 | 1 | 0 | R. aeschlimannii |
| 0 | 0 | 0 | R. massiliae | |
| 0 | 0 | 0 | R. sibirica subsp. mongolitimonae | |
| Total | 47 | 30 | 27 | Rickettsia spp. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kratou, M.; Belkahia, H.; Selmi, R.; Andolsi, R.; Dhibi, M.; Mhadhbi, M.; Messadi, L.; Ben Said, M. Diversity and Phylogeny of Cattle Ixodid Ticks and Associated Spotted Fever Group Rickettsia spp. in Tunisia. Pathogens 2023, 12, 552. https://doi.org/10.3390/pathogens12040552
Kratou M, Belkahia H, Selmi R, Andolsi R, Dhibi M, Mhadhbi M, Messadi L, Ben Said M. Diversity and Phylogeny of Cattle Ixodid Ticks and Associated Spotted Fever Group Rickettsia spp. in Tunisia. Pathogens. 2023; 12(4):552. https://doi.org/10.3390/pathogens12040552
Chicago/Turabian StyleKratou, Myriam, Hanene Belkahia, Rachid Selmi, Rihab Andolsi, Mokhtar Dhibi, Moez Mhadhbi, Lilia Messadi, and Mourad Ben Said. 2023. "Diversity and Phylogeny of Cattle Ixodid Ticks and Associated Spotted Fever Group Rickettsia spp. in Tunisia" Pathogens 12, no. 4: 552. https://doi.org/10.3390/pathogens12040552
APA StyleKratou, M., Belkahia, H., Selmi, R., Andolsi, R., Dhibi, M., Mhadhbi, M., Messadi, L., & Ben Said, M. (2023). Diversity and Phylogeny of Cattle Ixodid Ticks and Associated Spotted Fever Group Rickettsia spp. in Tunisia. Pathogens, 12(4), 552. https://doi.org/10.3390/pathogens12040552