
Transmission Electron Microscopy Observation of Morphological Changes to Cryptophlebia Leucotreta Granulovirus following Ultraviolet Irradiation


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Virus Purification
2.2. Enumeration of OBs by Dark Field Microscopy
2.3. UV Irradiation
2.4. Preparation of Samples for TEM
2.5. Data Analysis
3. Results
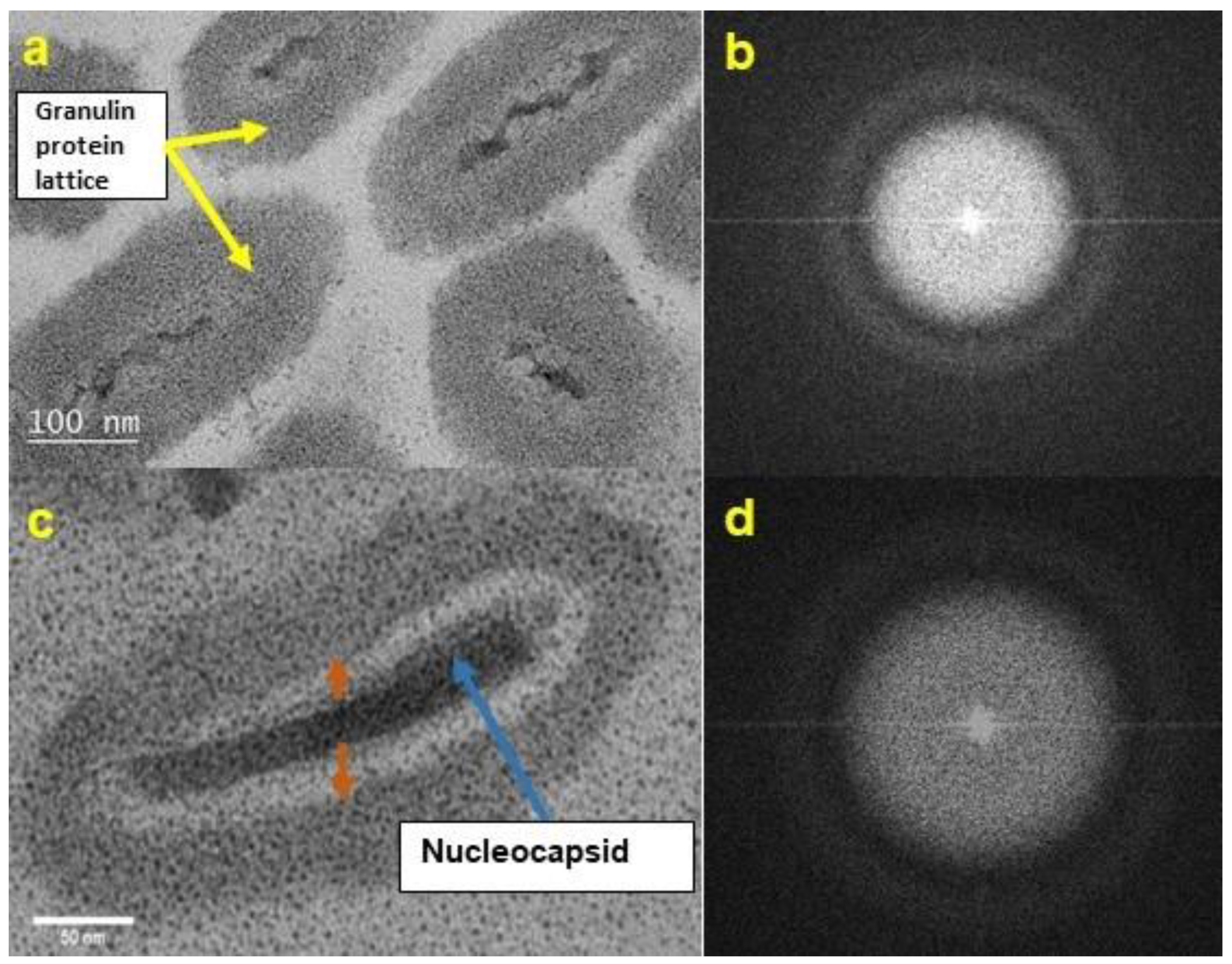
3.1. TEM Morphology of Unexposed CrleGV-SA
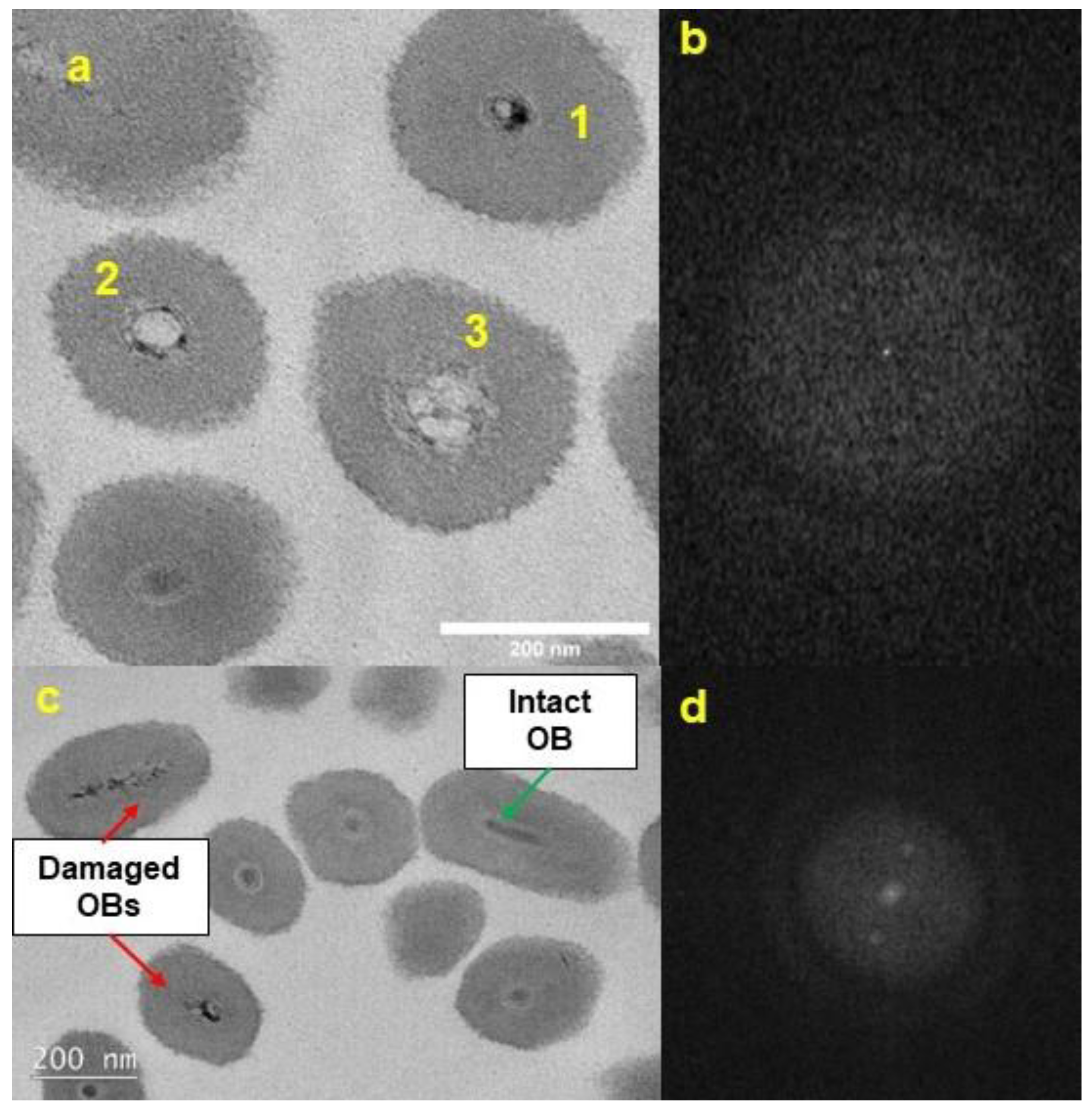
3.2. TEM Morphology of UV-Exposed CrleGV-SA
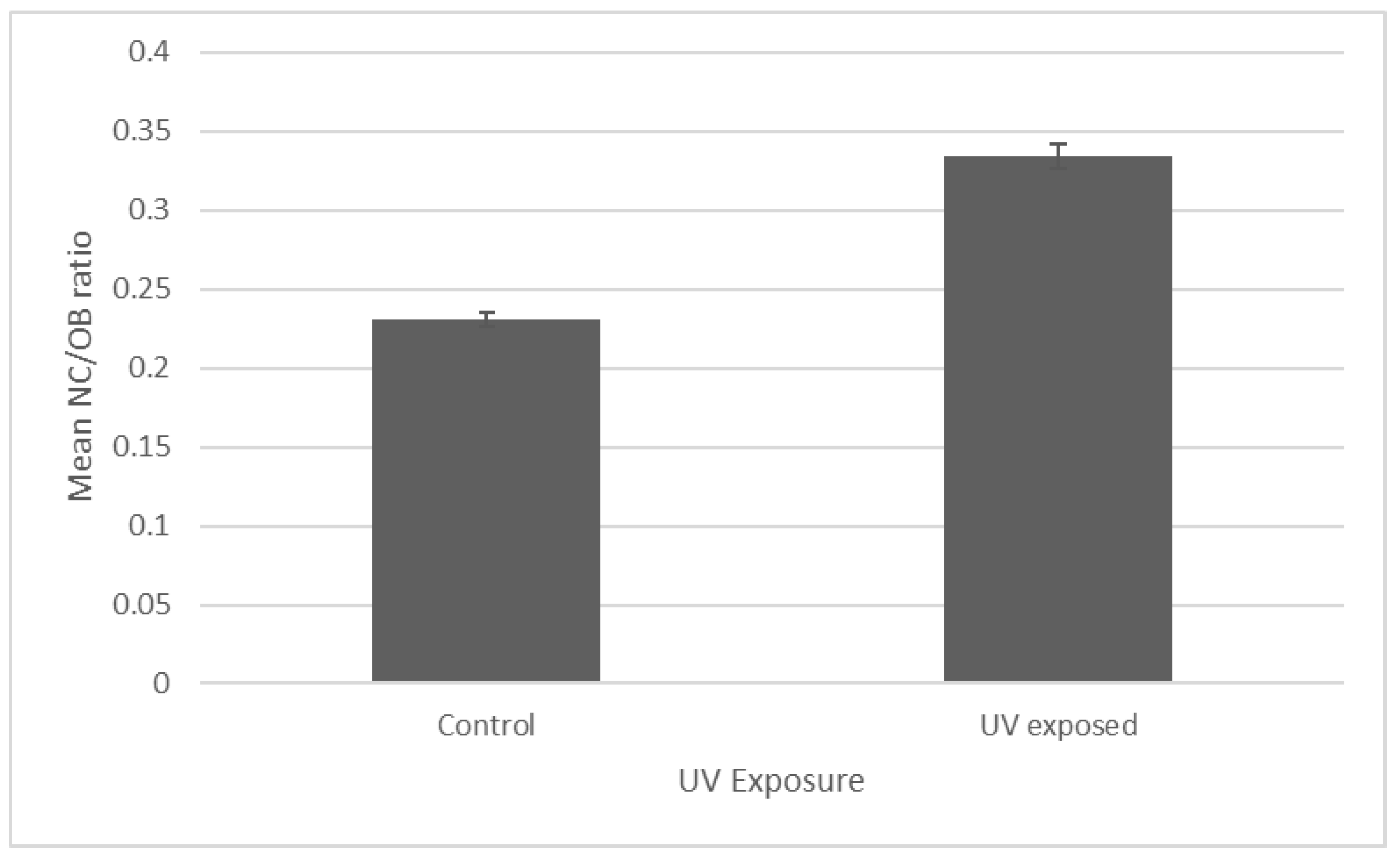
3.3. Measurement of OB Dimensions of UV-Irradiated CrleGV-SA
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Jehle, J.A.; Blissard, G.W.; Bonning, B.C.; Cory, J.S.; Herniou, E.A.; Rohrmann, G.F.; Theilmann, D.A.; Thiem, S.M.; Vlak, J.M. On the classification and nomenclature of baculoviruses: A proposal for revision. Arch. Virol. 2006, 151, 1257–1266. [Google Scholar] [CrossRef] [PubMed]
- Herniou, E.A.; Luque, T.; Chen, X.; Vlak, J.M.; Winstanley, D.; Cory, J.S.; O’Reilly, D.R. Use of whole genome sequence data to infer baculovirus phylogeny. J. Virol. 2001, 75, 8117–8126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beas-Catena, A.; Sanchez-Miron, A.; Garcia-Camacho, F.; Contreras-Gomez, A.; Molina-Grima, E. Baculovirus biopesticides: An overview. J. Anim. Plant Sci. 2014, 24, 362–373. [Google Scholar]
- Szewczyk, B.; Hoyos-Carvajal, L.; Paluszek, M.; Skrzecz, I.; Lobo De Souza, M. Baculoviruses—Re-emerging biopesticides. Biotechnol. Adv. 2006, 24, 143–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajek, A. Natural Enemies: An Introduction to Biological Control; Cambridge University Press: Cambridge, UK, 2004. [Google Scholar]
- Moscardi, F.; Lobo de Souza, M.; Batista de Castro, M.E.; Moscardi, M.L.; Szewczyk, B. Baculovirus pesticides: Present state and future perspectives. In Microbes and Microbial Technology; Ahmad, I., Ahmad, F., Pichtel, J., Eds.; Springer: New York, NY, USA, 2011; pp. 415–445. [Google Scholar]
- Moore, S.D.; Kirkman, W.; Richards, G.I.; Stephen, P. The Cryptophlebia leucotreta granulovirus—10 years of commercial field use. Viruses 2015, 7, 1284–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohrmann, G.F. Polyhedrin structure. J. Gen. Virol. 1986, 67, 1499–1513. [Google Scholar] [CrossRef]
- Grzywacz, D. Basic and applied research: Baculoviruses. In Microbial Control of Insect and Mite Pests: From Theory to Practice; Lacey, L.A., Ed.; Academic Press: London, UK, 2017; pp. 27–46. [Google Scholar]
- Moore, S.D. Biological control of a phytosanitary pest (Thaumatotibia leucotreta): A case study. Int. J. Environ. Res. Public Health 2021, 18, 1198. [Google Scholar] [CrossRef]
- David, W.A.L.; Gardiner, B.O.C. Persistence of a granulosis virus of Pieris brassicae on cabbage leaves. J. Invertebr. Pathol. 1966, 8, 180–183. [Google Scholar] [CrossRef]
- Young, S.Y.; Yearian, W.C. Persisstence of heliothis NPV on foliage of cotton, soybean and tomato. Environ. Entomol. 1974, 3, 253–255. [Google Scholar] [CrossRef]
- Jacques, R.P. Stability of entomopathogenic viruses in the environment. In Viral Insecticides for Biological Control; Maramorosch, K., Sherman, K.E., Eds.; Academic Press: New York, NY, USA, 1985; pp. 285–360. [Google Scholar]
- Burges, H.D.; Jones, K.A. Formulation of bacteria, viruses and protozoa to control insects. In Formulation of Microbial Biopesticides; Burges, H.D., Ed.; Springer: Dordrecht, The Netherlands, 1998; pp. 33–127. [Google Scholar]
- Shapiro, M.; Farrar, R.R.; Domek, J.; Javais, I. Effects of virus concentration and ultraviolet irradiation on the activity of corn earworm and beet army worm (Lepidoptera: Noctuidae) nucleopolyhedroviruses. J. Econ. Entomol. 2002, 95, 243–249. [Google Scholar] [CrossRef]
- Jeyarani, S.; Sathiah, N.; Karuppuchamy, P. An in vitro method for increasing UV-tolerance in a strain of Helicoverpa armigera (Lepidoptera: Noctuidae) nucleopolyhedrovirus. Biocontrol Sci. Technol. 2013, 23, 305–316. [Google Scholar] [CrossRef]
- Bergold, G.H. Die isolierung des polyeder-virus and die natur der polyeder. Z. Naturforsch. B 1947, 2, 122–143. [Google Scholar] [CrossRef]
- Ignoffo, C.M. Development of a viral insecticide: Concept to commercialization. Exp. Parasitol. 1973, 33, 380–406. [Google Scholar] [CrossRef] [PubMed]
- Dhladhla, B.I.R.; Mwanza, P.; Lee, M.E.; Moore, S.D.; Dealtry, G.B. Comparison of microscopic and molecular enumeration methods for insect viruses: Cryptophlebia leucotreta granulovirus as a case study. J. Virol. Methods 2018, 256, 107–110. [Google Scholar] [CrossRef]
- Arnott, H.J.; Smith, K.M. An ultrastructural study of the development of a granulosis virus in the cells of the moth Plodia interpunctella (Hbn.). J. Ultrastruct. Res. 1968, 21, 251–268. [Google Scholar] [CrossRef]
- Akermann, H.W.; Smirnoff, W.A. A morphological investigation of 23 baculoviruses. J. Invertebr. Pathol. 1983, 41, 269–280. [Google Scholar] [CrossRef]
- Grasela, J.J.; McIntosh, A.H.; Shelby, K.S.; Long, S. Isolation and characterization of a baculovirus associated with the insect parasitoid wasp, Cotesia marginiventris, or its host, Trichoplusia ni. J. Insect Sci. 2008, 8, 42. [Google Scholar] [CrossRef]
- Bayramoglu, Z.; Gencer, D.; Muratoglu, H.; Efe, D.; Nalcacioglu, R.; Demirbag, Z.; Demir, I. Characterization of a nucleopolyhedrovirus variant of the gypsy moth, Lymantria dispar (Lepidoptera: Lymantriidae) in Turkey. Int. J. Pest Manag. 2018, 64, 119–127. [Google Scholar] [CrossRef]
- Hughes, K.M.; Addison, R.B. Two nuclear polyhedrosis viruses of the Douglas-fir tussock moth. J. Invertebr. Pathol. 1970, 16, 196–204. [Google Scholar] [CrossRef]
- Matilainen, H.; Rinne, J.; Gilbert, L.; Marjomäki, V.; Reunanen, H.; Oker-Blom, C. Baculovirus Entry into Human Hepatoma Cells. J. Virol. 2005, 79, 15452–15459. [Google Scholar] [CrossRef] [Green Version]
- Evans, H.F.; Shapiro, M. Viruses. In Manual of Techniques in Insect Pathology; Lacey, L.A., Ed.; Academic Press: San Diego, CA, USA, 1997; pp. 17–53. [Google Scholar]
- Hunter-Fujita, F.R.; Entwistle, P.F.; Evans, H.F.; Crook, N.E. Insect Viruses and Pest Management; Wiley: Chichester, UK, 1998. [Google Scholar]
- Brassel, J.; Benz, G. Selection of a strain of the granulosis virus of the codling moth with improved resistance against artificial ultraviolet radiation and sunlight. J. Invertebr. Pathol. 1979, 33, 358–363. [Google Scholar] [CrossRef]
- Jehle, J.A.; Baekhaus, H.; Fritsch, E.; Huber, J. Physical map of the Cryptophlebia leucotreta granulosis virus genome and its relationship to the genome of Cydia pomoneila granulosis virus. J. Gen. Virol. 1992, 73, 1621–1626. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.D. The Development and Evaluation of Cryptophlebia leucotreta Granulovirus (CrleGV) as a Biological Control Agent for the Management of False Codling Moth, Cryptophlebia leucotreta, on Citrus. Ph.D. Thesis, Rhodes University, Grahamstown, South Africa, 2002. [Google Scholar]
- Wolff, J.L.; Moraes, R.H.P.; Kitajima, P.; de Souza Leal, E.; Zanotto, P.M.A. Identification and characterization of a baculovirus from Lonomia obliqua (Lepidoptera: Saturniidae). J. Invertebr. Pathol. 2002, 79, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Dhladhla, B.I.R. Enumeration of Insect Viruses Using Microscopic and Molecular Analyses; South African Isolate of Cryptophlebia leucotreta Granulovirus As a Case Study. Master’s Thesis, Nelson Mandela Metropolitan University, Gqeberha, South Africa, 2012. [Google Scholar]
- Rohrmann, G.F. Baculovirus structural proteins. J. Gen. Virol. 1992, 73, 749–761. [Google Scholar] [CrossRef] [PubMed]
- Mwanza, P. Determination of the Effects of Sunlight and UV Irradiation on the Structure, Viability and Reapplication Frequency of the Biopesticide Cryptophlebia leucotreta Granulovirus in the Protection against False Codling Moth Infestation of Citrus Crops. Master’s Thesis, Nelson Mandela Metropolitan University, Gqeberha, South Africa, 2015. [Google Scholar]
- Anduleit, K.; Sutton, G.; Diprose, J.M.; Mertens, P.P.C.; Grimes, J.M.; Stuart, D.I. Crystal lattice as biological phenotype for insect viruses. Protein Sci. 2005, 14, 2741–2743. [Google Scholar] [CrossRef] [Green Version]
- Coulibaly, F.; Chiu, E.; Gutmann, S.; Rajendran, C.; Haebel, P.W.; Ikeda, K.; Metcalf, P. The atomic structure of baculovirus polyhedra reveals the independent emergence of infectious crystals in DNA and RNA viruses. Proc. Natl. Acad. Sci. USA 2009, 106, 22205–22210. [Google Scholar] [CrossRef] [Green Version]
- Gati, C.; Oberthuer, D.; Yefanov, O.; Bunker, R.D.; Stellato, F.; Chiu, E.; Yeh, S.; Aquila, A.; Basu, S.; Bean, R.; et al. Atomic structure of granulin determined from native nanocrystalline granulovirus using an X-ray free-electron laser. Proc. Natl. Acad. Sci. USA 2017, 114, 2247–2252. [Google Scholar] [CrossRef] [Green Version]
- Ignoffo, C.M.; Garcia, C. UV photo inactivation of cells and spores Bacillus thuringiensis and effects of peroxidase on inactivation. Environ. Entomol. 1978, 7, 270–272. [Google Scholar] [CrossRef]
- Ignoffo, C.M.; Rice, W.C.; Mcintosh, A.H. Inactivation of nonoccluded and occluded baculoviruses and baculovirus-DNA exposed to simulated sunlight. Environ. Entomol. 1989, 18, 177–183. [Google Scholar] [CrossRef]
- Ravanat, J.L.; Douki, T.; Cadet, J. Direct and indirect effects of UV radiation on DNA and its components. J. Photochem.Photobiol. B 2001, 63, 88–102. [Google Scholar] [CrossRef]
- Rohrmann, G.F. Baculovirus Molecular Biology, 3rd ed.; National Center for Biotechnology Information: Bethesda, MD, USA, 2013. Available online: https://www.ncbi.nlm.nih.gov/books/NBK138298/ (accessed on 5 November 2016).
- Angelini, A.; Amargier, A.; Vandamme, P.; Dutoit, J.L. Une virose a granules chez Ie lèpidoptére Argyroploce leucotreta. Coton Fibres Trop. 1965, 20, 277–282. [Google Scholar]
- Mück, O. Biologie, Verhalten und Wirtschaftliche Bedeutung von Parasiten Schädlicher Lepidopteren auf den Kapverden; Bauer: Exeter, NH, USA, 1985. [Google Scholar]
- Singh, S.; Moore, S.D.; Spillings, B.; Hendry, D. South African Isolate of Cryptophlebia leucotreta granulovirus. J. Invertebr. Pathol. 2003, 83, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Opoku-Debrah, J.K.; Hill, M.P.; Knox, C.; Moore, S.D. Heterogeneity in virulence relationships between Cryptophlebia leucotreta granulovirus isolates and geographically distinct host populations: Lessons from codling moth resistance to CpGV-M. BioControl 2016, 61, 449–459. [Google Scholar] [CrossRef]
- Hatting, J.L.; Moore, S.D.; Malan, A.P. Microbial control of phytophagous invertebrate pests in South Africa: Current status and future prospects. J. Invertebr. Pathol. 2018, 165, 54–66. [Google Scholar] [CrossRef]
- Shapiro, M.; Bell, R.A. Selection of a UV-tolerant strain of gypsy moth, Lymantria dispar (L.) (Lepidoptera: Lymantriidae), nucleopolyhedrosis virus. Environ. Entomol. 1984, 13, 1522–1526. [Google Scholar] [CrossRef]
- Mwanza, P.; Jukes, M.; Dealtry, G.; Lee, M.; Moore, S. Selection for and Analysis of UV-Resistant Cryptophlebia leucotreta granulovirus-SA as a Biopesticide for Thaumatotibia leucotreta. Viruses 2022, 14, 28. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dimension (nm) | Control Non-Exposed CrleGV-SA 1 | UV Exposed CrleGV-SA 1 |
|---|---|---|
| OB length | 365.31 ± 4.91 | 301.30 ± 6.03 |
| OB width | 213.47 ± 3.16 | 184.03 ± 3.60 |
| NC length | 210.16 ± 3.89 | 182.76 ± 5.56 |
| NC width | 48.87 ± 4.89 | 58.57 ± 1.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mwanza, P.; Dealtry, G.; Lee, M.; Moore, S. Transmission Electron Microscopy Observation of Morphological Changes to Cryptophlebia Leucotreta Granulovirus following Ultraviolet Irradiation. Pathogens 2023, 12, 590. https://doi.org/10.3390/pathogens12040590
Mwanza P, Dealtry G, Lee M, Moore S. Transmission Electron Microscopy Observation of Morphological Changes to Cryptophlebia Leucotreta Granulovirus following Ultraviolet Irradiation. Pathogens. 2023; 12(4):590. https://doi.org/10.3390/pathogens12040590
Chicago/Turabian StyleMwanza, Patrick, Gill Dealtry, Michael Lee, and Sean Moore. 2023. "Transmission Electron Microscopy Observation of Morphological Changes to Cryptophlebia Leucotreta Granulovirus following Ultraviolet Irradiation" Pathogens 12, no. 4: 590. https://doi.org/10.3390/pathogens12040590
APA StyleMwanza, P., Dealtry, G., Lee, M., & Moore, S. (2023). Transmission Electron Microscopy Observation of Morphological Changes to Cryptophlebia Leucotreta Granulovirus following Ultraviolet Irradiation. Pathogens, 12(4), 590. https://doi.org/10.3390/pathogens12040590