



First Report on Detection and Complete Genomic Analysis of a Novel CRESS DNA Virus from Sea Turtles

 ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
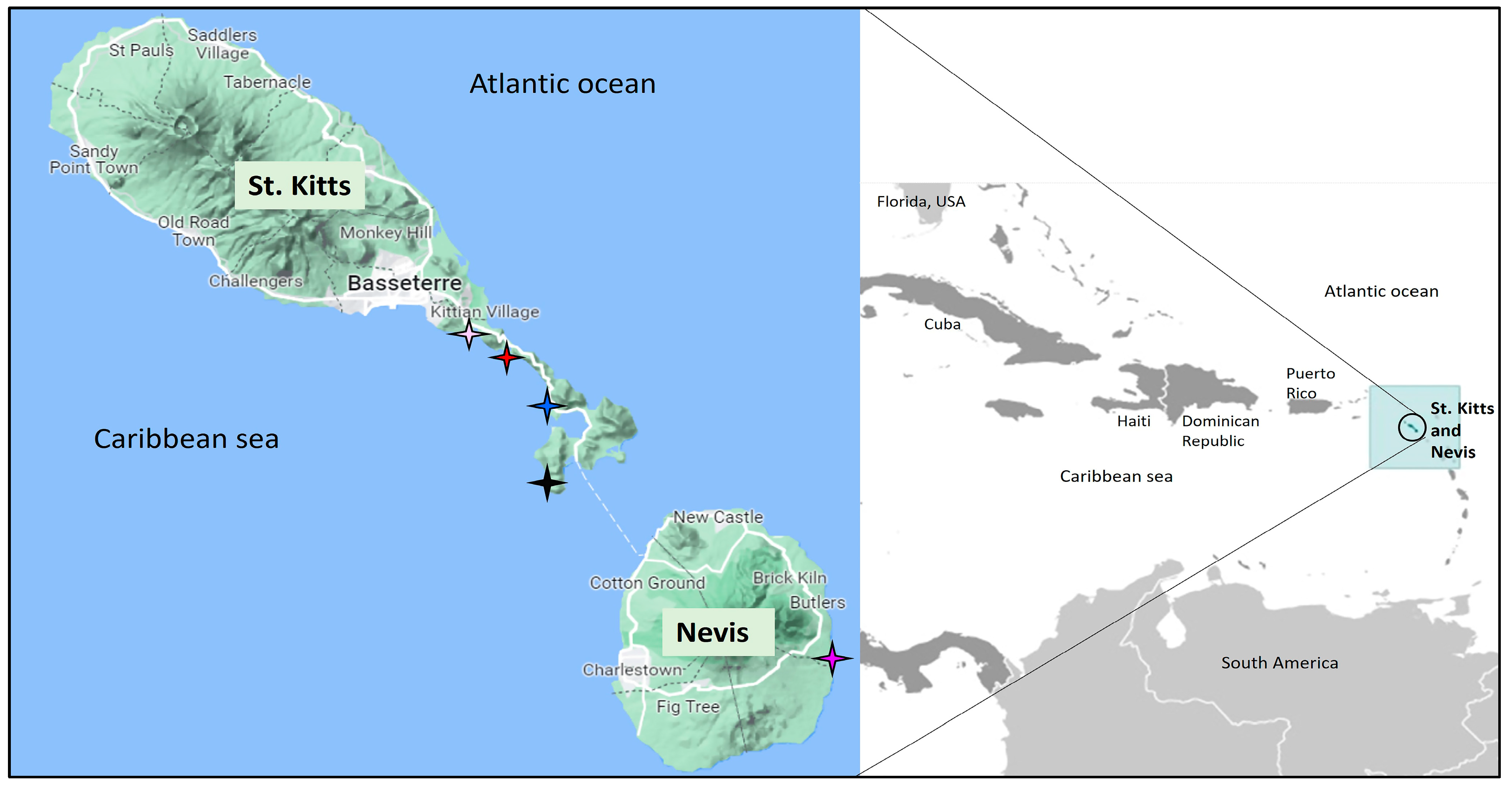
2.1. Sampling
2.2. Amplification of Viral DNA
2.3. Nucleotide Sequencing
2.4. Sequence Analysis
2.5. GenBank Accession Numbers
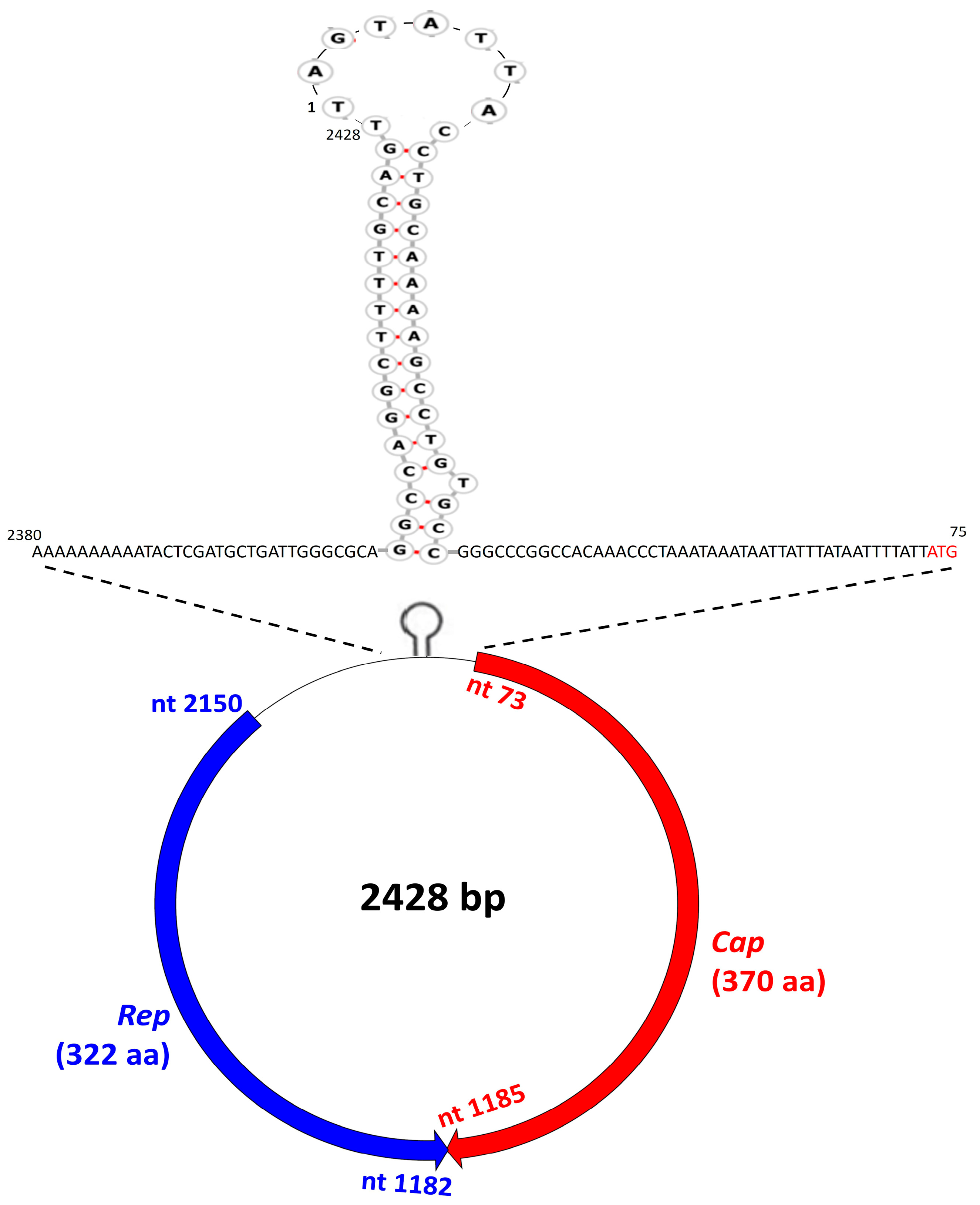
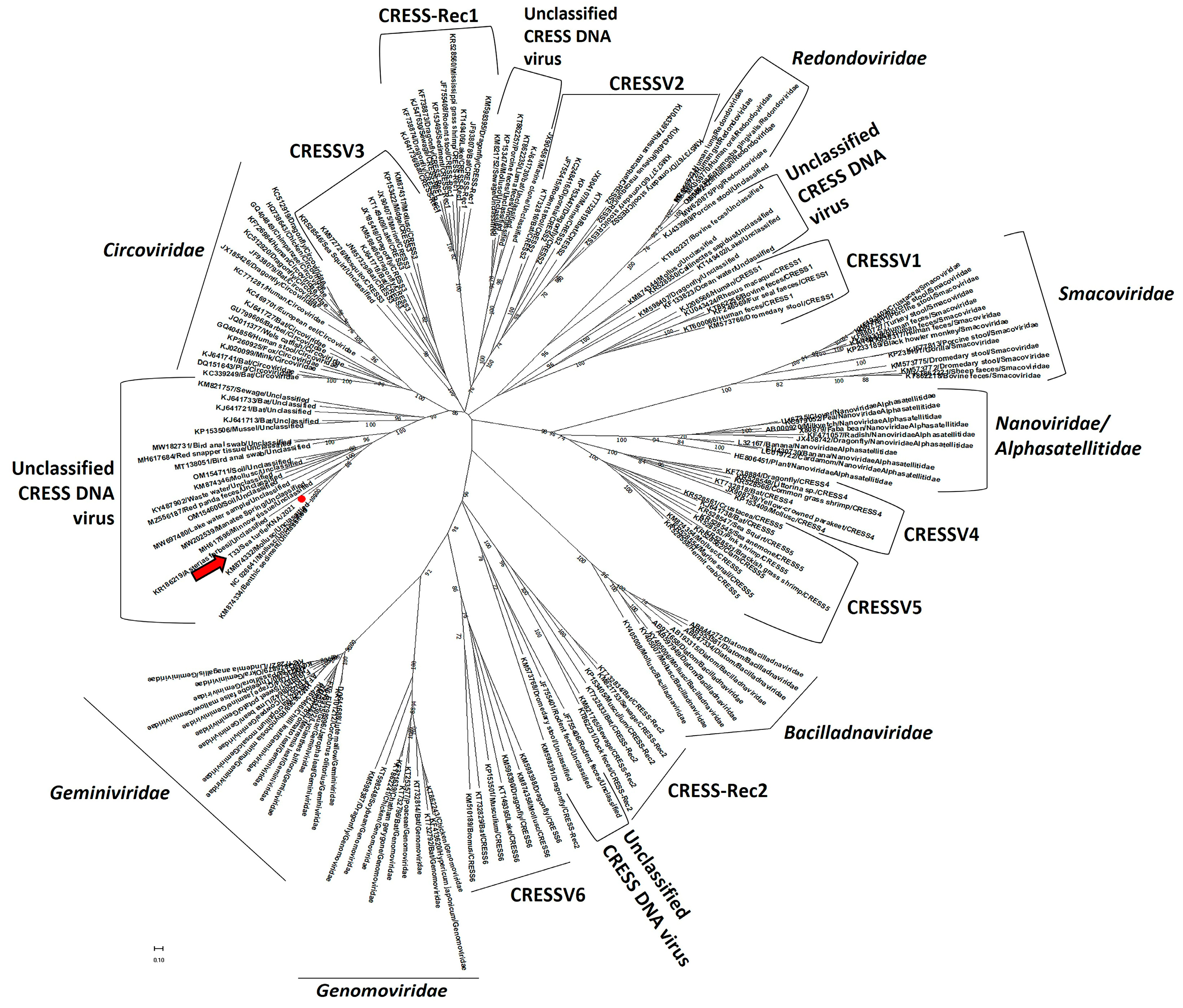
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bennett, L. Sea Turtles. Smithsonian Ocean Portal. Available online: https://ocean.si.edu/ocean-life/reptiles/sea-turtles (accessed on 1 March 2023).
- Sea Turtle Conservancy. Information about Sea Turtles: An Introduction. Available online: https://conserveturtles.org/information-about-sea-turtles-an-introduction/ (accessed on 1 March 2023).
- National Geographic Society. Sea Turtles 101. Available online: https://www.nationalgeographic.com/animals/reptiles/facts/sea-turtles (accessed on 1 March 2023).
- Mashkour, N.; Jones, K.; Kophamel, S.; Hipolito, T.; Ahasan, S.; Walker, G.; Jakob-Hoff, R.; Whittaker, M.; Hamann, M.; Bell, I.; et al. Disease Risk Analysis in Sea Turtles: A Baseline Study to Inform Conservation Efforts. PLoS ONE 2020, 15, e0230760. [Google Scholar] [CrossRef] [PubMed]
- ICUN. The IUCN Red List of Threatened Species. 2023. Available online: https://www.iucnredlist.org/search?query=sea%20turtle&searchType=species (accessed on 1 March 2023).
- Smith, K.F.; Sax, D.F.; Lafferty, K.D. Evidence for the Role of Infectious Disease in Species Extinction and Endangerment. Conserv. Biol. 2006, 20, 1349–1357. [Google Scholar] [CrossRef] [PubMed]
- Okoh, G.R.; Horwood, P.F.; Whitmore, D.; Ariel, E. Herpesviruses in Reptiles. Front. Vet. Sci. 2021, 8, 642894. [Google Scholar] [CrossRef]
- Jones, K.; Ariel, E.; Burgess, G.; Read, M. A Review of Fibropapillomatosis in Green Turtles (Chelonia mydas). Vet. J. 2016, 212, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Robben, D.M.; Palaniappan, P.; Loganathan, A.L.; Subbiah, V.K. Increased Prevalence and New Evidence of Multi-Species Chelonid Herpesvirus 5 (ChHV5) Infection in the Sea Turtles of Mabul Island, Borneo. Animals 2023, 13, 290. [Google Scholar] [CrossRef]
- Stacy, B.A.; Wellehan, J.F.X.; Foley, A.M.; Coberley, S.S.; Herbst, L.H.; Manire, C.A.; Garner, M.M.; Brookins, M.D.; Childress, A.L.; Jacobson, E.R. Two Herpesviruses Associated with Disease in Wild Atlantic Loggerhead Sea Turtles (Caretta caretta). Vet. Microbiol. 2008, 126, 63–73. [Google Scholar] [CrossRef]
- Manire, C.A.; Stacy, B.A.; Kinsel, M.J.; Daniel, H.T.; Anderson, E.T.; Wellehan, J.F.X. Proliferative Dermatitis in a Loggerhead Turtle, Caretta caretta, and a Green Turtle, Chelonia mydas, Associated with Novel Papillomaviruses. Vet. Microbiol. 2008, 130, 227–237. [Google Scholar] [CrossRef]
- Mashkour, N.; Maclaine, A.; Burgess, G.W.; Ariel, E. Discovery of an Australian Chelonia mydas Papillomavirus via Green Turtle Primary Cell Culture and QPCR. J. Virol. Methods 2018, 258, 13–23. [Google Scholar] [CrossRef]
- Mashkour, N.; Jones, K.; Wirth, W.; Burgess, G.; Ariel, E. The Concurrent Detection of Chelonid Alphaherpesvirus 5 and Chelonia mydas Papillomavirus 1 in Tumoured and Non-Tumoured Green Turtles. Animals 2021, 11, 697. [Google Scholar] [CrossRef]
- Sarker, S.; Hannon, C.; Athukorala, A.; Bielefeldt-Ohmann, H. Emergence of a Novel Pathogenic Poxvirus Infection in the Endangered Green Sea Turtle (Chelonia mydas) Highlights a Key Threatening Process. Viruses 2021, 13, 219. [Google Scholar] [CrossRef]
- Casey, R.; Quackenbush, S.; Work, T.; Balazs, G.; Bowser, P.; Casey, J. Evidence for Retrovirus Infections in Green Turtles Chelonia mydas from the Hawaiian Islands. Dis. Aquat. Organ. 1997, 31, 1–7. [Google Scholar] [CrossRef]
- Fichi, G.; Cardeti, G.; Cersini, A.; Mancusi, C.; Guarducci, M.; di Guardo, G.; Terracciano, G. Bacterial and Viral Pathogens Detected in Sea Turtles Stranded along the Coast of Tuscany, Italy. Vet. Microbiol. 2016, 185, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.F.F.; Manire, C.; Borrowman, K.; Langer, T.; Ehrhart, L.; Breitbart, M. Discovery of a Novel Single-Stranded DNA Virus from a Sea Turtle Fibropapilloma by Using Viral Metagenomics. J. Virol. 2009, 83, 2500–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krupovic, M.; Varsani, A.; Kazlauskas, D.; Breitbart, M.; Delwart, E.; Rosario, K.; Yutin, N.; Wolf, Y.I.; Harrach, B.; Zerbini, F.M.; et al. Cressdnaviricota: A Virus Phylum Unifying Seven Families of Rep-Encoding Viruses with Single-Stranded, Circular DNA Genomes. J. Virol. 2020, 94, e00582-20. [Google Scholar] [CrossRef] [PubMed]
- Kazlauskas, D.; Varsani, A.; Krupovic, M. Pervasive Chimerism in the Replication-Associated Proteins of Uncultured Single-Stranded DNA Viruses. Viruses 2018, 10, 187. [Google Scholar] [CrossRef] [Green Version]
- Desingu, P.A.; Nagarajan, K. Genetic Diversity and Characterization of Circular Replication (Rep)-Encoding Single-Stranded (CRESS) DNA Viruses. Microbiol. Spectr. 2022, 10, e0105722. [Google Scholar] [CrossRef]
- Kazlauskas, D.; Varsani, A.; Koonin, E.V.; Krupovic, M. Multiple Origins of Prokaryotic and Eukaryotic Single-Stranded DNA Viruses from Bacterial and Archaeal Plasmids. Nat. Commun. 2019, 10, 3425. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Rosario, K.; Breitbart, M.; Duffy, S. Eukaryotic Circular Rep-Encoding Single-Stranded DNA (CRESS DNA) Viruses: Ubiquitous Viruses with Small Genomes and a Diverse Host Range. Adv. Virus Res. 2019, 103, 71–133. [Google Scholar] [CrossRef]
- Dayaram, A.; Goldstien, S.; Argüello-Astorga, G.R.; Zawar-Reza, P.; Gomez, C.; Harding, J.S.; Varsani, A. Diverse Small Circular DNA Viruses Circulating amongst Estuarine Molluscs. Infect. Genet. Evol. 2015, 31, 284–295. [Google Scholar] [CrossRef]
- Rosario, K.; Duffy, S.; Breitbart, M. Diverse Circovirus-like Genome Architectures Revealed by Environmental Metagenomics. J. General Virol. 2009, 90, 2418–2424. [Google Scholar] [CrossRef]
- Fahsbender, E.; Hewson, I.; Rosario, K.; Tuttle, A.D.; Varsani, A.; Breitbart, M. Discovery of a Novel Circular DNA Virus in the Forbes Sea Star, Asterias Forbesi. Arch. Virol. 2015, 160, 2349–2351. [Google Scholar] [CrossRef] [PubMed]
- Jackson, E.W.; Bistolas, K.S.I.; Button, J.B.; Hewson, I. Novel Circular Single-Stranded DNA Viruses among an Asteroid, Echinoid and Holothurian (Phylum: Echinodermata). PLoS ONE 2016, 11, e0166093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosario, K.; Schenck, R.O.; Harbeitner, R.C.; Lawler, S.N.; Breitbart, M. Novel Circular Single-Stranded DNA Viruses Identified in Marine Invertebrates Reveal High Sequence Diversity and Consistent Predicted Intrinsic Disorder Patterns within Putative Structural Proteins. Front. Microbiol. 2015, 6, 696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landrau-Giovannetti, N.; Subramaniam, K.; Brown, M.A.; Ng, T.F.F.; Rotstein, D.S.; West, K.; Frasca, S.; Waltzek, T.B. Genomic Characterization of a Novel Circovirus from a Stranded Longman’s Beaked Whale (Indopacetus Pacificus). Virus Res. 2020, 277, 197826. [Google Scholar] [CrossRef] [PubMed]
- St. Kitts Sea Turtle Monitoring Network. Available online: https://www.stkittsturtles.org/ (accessed on 1 March 2023).
- Li, L.; Kapoor, A.; Slikas, B.; Bamidele, O.S.; Wang, C.; Shaukat, S.; Masroor, M.A.; Wilson, M.L.; Ndjango, J.-B.N.; Peeters, M.; et al. Multiple Diverse Circoviruses Infect Farm Animals and Are Commonly Found in Human and Chimpanzee Feces. J. Virol. 2010, 84, 1674–1682. [Google Scholar] [CrossRef] [Green Version]
- Gainor, K.; Becker, A.A.M.J.; Malik, Y.S.; Ghosh, S. Detection and Complete Genome Analysis of Circoviruses and Cycloviruses in the Small Indian Mongoose (Urva auropunctata): Identification of Novel Species. Viruses 2021, 13, 1700. [Google Scholar] [CrossRef] [PubMed]
- Gainor, K.; Castillo Fortuna, Y.; Alakkaparambil, A.S.; González, W.; Malik, Y.S.; Ghosh, S. Detection and Complete Genome Analysis of Porcine Circovirus 2 (PCV2) and an Unclassified CRESS DNA Virus from Diarrheic Pigs in the Dominican Republic: First Evidence for Predominance of PCV2d from the Caribbean Region. Viruses 2022, 14, 1799. [Google Scholar] [CrossRef]
- Zuker, M. Mfold Web Server for Nucleic Acid Folding and Hybridization Prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Rosario, K.; Breitbart, M.; Harrach, B.; Segalés, J.; Delwart, E.; Biagini, P.; Varsani, A. Revisiting the Taxonomy of the Family Circoviridae: Establishment of the Genus Cyclovirus and Removal of the Genus Gyrovirus. Arch. Virol. 2017, 162, 1447–1463. [Google Scholar] [CrossRef] [Green Version]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A Virus Classification Tool Based on Pairwise Sequence Alignment and Identity Calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef]
- The Information Architects of Encyclopedia of Britannica. Saint Kitts and Nevis. Available online: https://www.britannica.com/facts/Saint-Kitts-and-Nevis (accessed on 1 March 2023).
- St. Kitts Sea Turtle Monitoring Network. Kayden’s Journey. Available online: https://197497-28bf.icpage.net/kaydens-journey (accessed on 2 March 2023).
- Dayaram, A.; Galatowitsch, M.L.; Argüello-Astorga, G.R.; van Bysterveldt, K.; Kraberger, S.; Stainton, D.; Harding, J.S.; Roumagnac, P.; Martin, D.P.; Lefeuvre, P.; et al. Diverse Circular Replication-Associated Protein Encoding Viruses Circulating in Invertebrates within a Lake Ecosystem. Infect. Genet. Evol. 2016, 39, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Gainor, K.; Becker, A.A.M.J.; Malik, Y.S.; Ghosh, S. First Report on Detection and Molecular Characterization of Adenoviruses in the Small Indian Mongoose (Urva auropunctata). Viruses 2021, 13, 2194. [Google Scholar] [CrossRef] [PubMed]
- Kleymann, A.; Becker, A.A.M.J.; Malik, Y.S.; Kobayashi, N.; Ghosh, S. Detection and Molecular Characterization of Picobirnaviruses (PBVs) in the Mongoose: Identification of a Novel PBV Using an Alternative Genetic Code. Viruses 2020, 12, 99. [Google Scholar] [CrossRef] [Green Version]
- Rosario, K.; Duffy, S.; Breitbart, M. A Field Guide to Eukaryotic Circular Single-Stranded DNA Viruses: Insights Gained from Metagenomics. Arch. Virol. 2012, 157, 1851–1871. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, C.E.; Alexander, K.A. Unchartered Waters: Climate Change Likely to Intensify Infectious Disease Outbreaks Causing Mass Mortality Events in Marine Mammals. Glob. Chang. Biol. 2020, 26, 4284–4301. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gainor, K.; Stewart, K.M.; Picknell, A.; Russ, M.; Makela, N.; Watson, K.; Mancuso, D.M.; Malik, Y.S.; Ghosh, S. First Report on Detection and Complete Genomic Analysis of a Novel CRESS DNA Virus from Sea Turtles. Pathogens 2023, 12, 601. https://doi.org/10.3390/pathogens12040601
Gainor K, Stewart KM, Picknell A, Russ M, Makela N, Watson K, Mancuso DM, Malik YS, Ghosh S. First Report on Detection and Complete Genomic Analysis of a Novel CRESS DNA Virus from Sea Turtles. Pathogens. 2023; 12(4):601. https://doi.org/10.3390/pathogens12040601
Chicago/Turabian StyleGainor, Kerry, Kimberly M. Stewart, Angela Picknell, Morgan Russ, Noah Makela, Kierra Watson, Diana M. Mancuso, Yashpal Singh Malik, and Souvik Ghosh. 2023. "First Report on Detection and Complete Genomic Analysis of a Novel CRESS DNA Virus from Sea Turtles" Pathogens 12, no. 4: 601. https://doi.org/10.3390/pathogens12040601
APA StyleGainor, K., Stewart, K. M., Picknell, A., Russ, M., Makela, N., Watson, K., Mancuso, D. M., Malik, Y. S., & Ghosh, S. (2023). First Report on Detection and Complete Genomic Analysis of a Novel CRESS DNA Virus from Sea Turtles. Pathogens, 12(4), 601. https://doi.org/10.3390/pathogens12040601