

Colonization with Escherichia coli ST131-H30R (H30R) Corresponds with Increased Serum Anti-O25 IgG Levels and Decreased TNFα and IL-10 Responsiveness to H30R


Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects, Fecal Samples, and Strain Typing
2.2. Blood Sample Collection
2.3. O6 and O25b Serology
2.4. Antigen Stimulation of Whole-Blood Samples
2.5. Detection of Cytokines
2.6. Statistical Analysis
3. Results
3.1. Characteristics of the Study Population
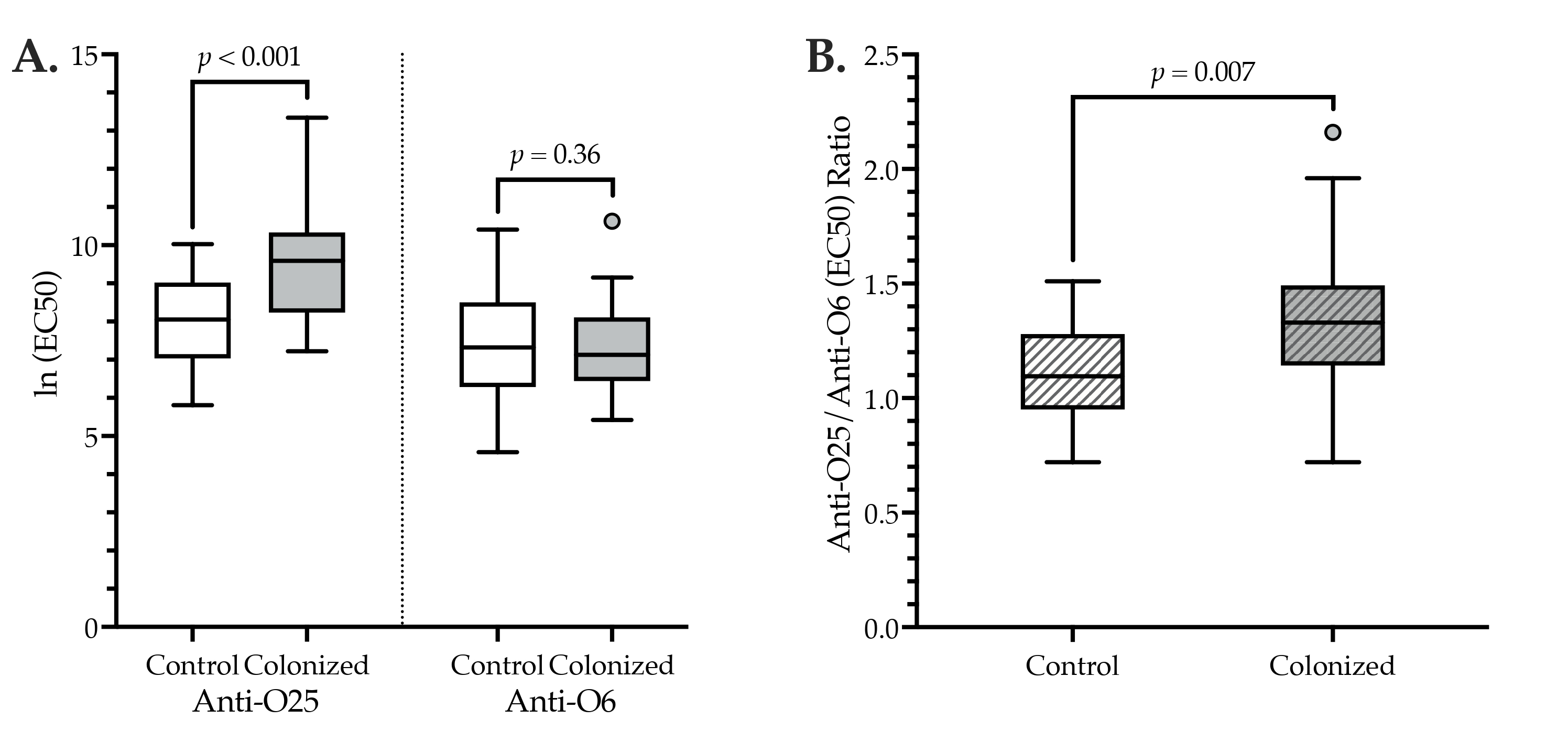
3.2. Anti-O25 and Anti-O6 IgG Levels: Cross-Sectional Comparisons
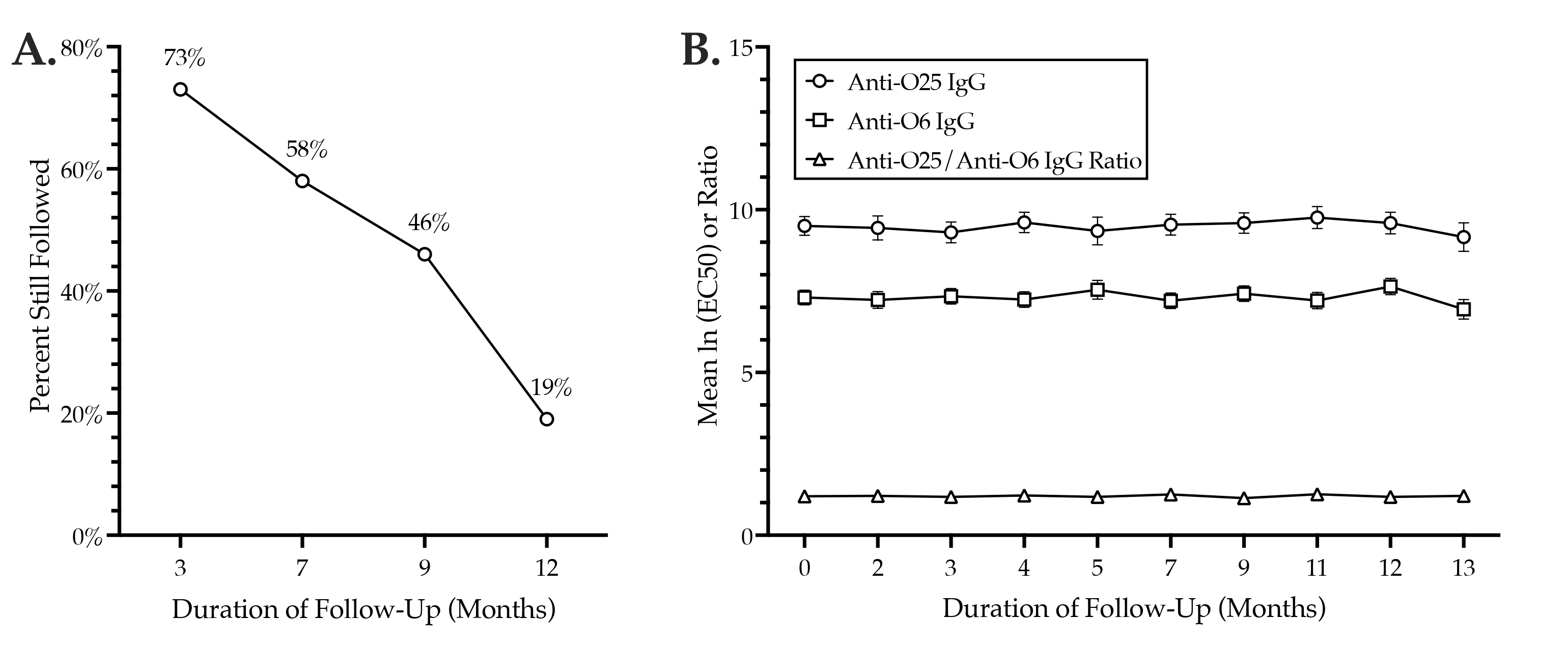
3.3. Anti-O25 and Anti-O6 IgG Levels: Longitudinal Comparisons
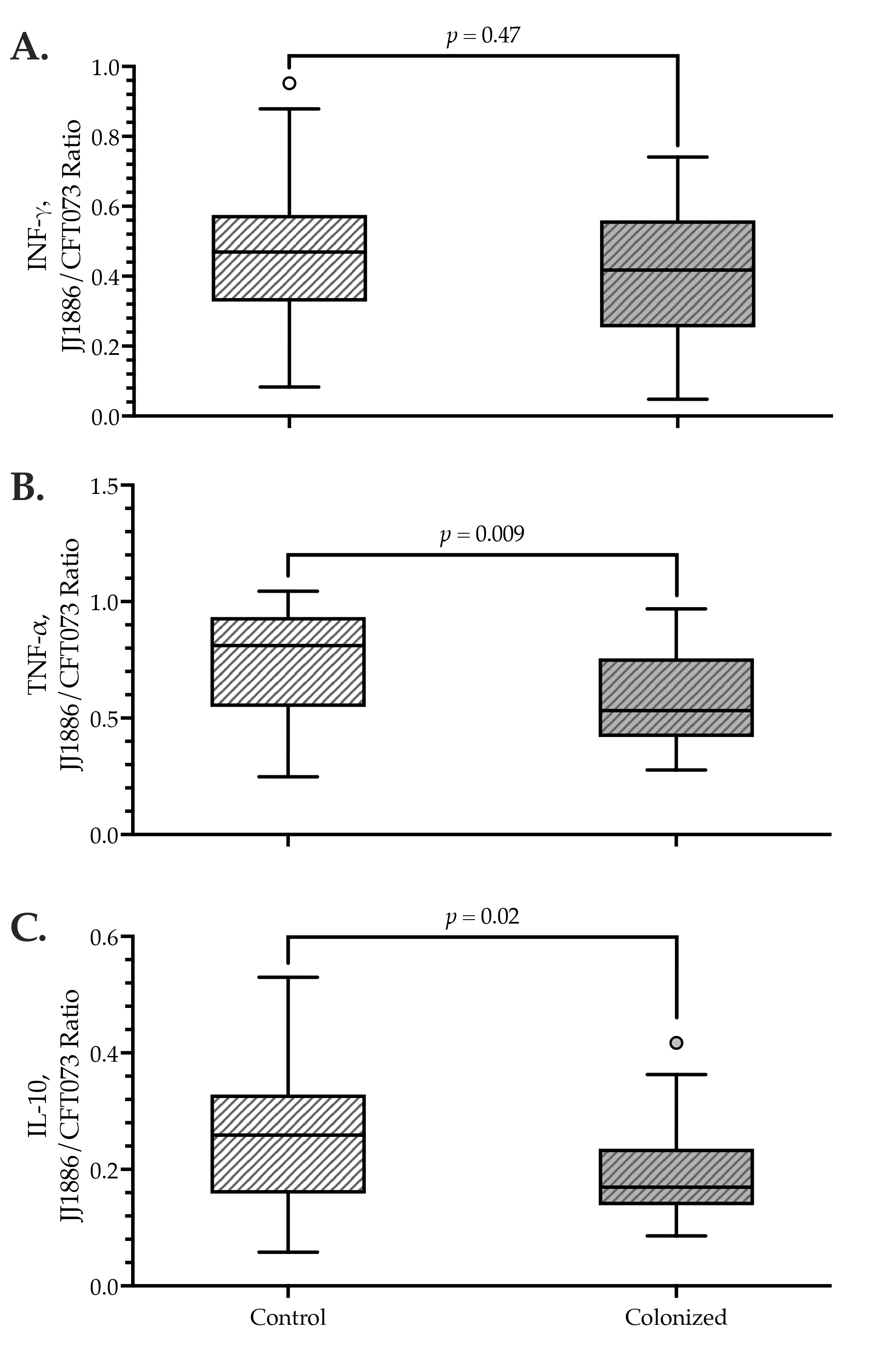
3.4. Antigen-Triggered Cytokine Release: Cross-Sectional Comparisons
3.5. Association of Immune Responses with Host Characteristics
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nicolas-Chanoine, M.-H.; Bertrand, X.; Madec, J.-Y. Escherichia coli ST131, an intriguing clonal group. Clin. Microbiol. Rev. 2014, 27, 543–574. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Porter, S.; Thuras, P.; Castanheira, M. Epidemic emergence in the United States of Escherichia coli sequence type 131-H30 (ST131-H30), 2000–2009. Antimicrob. Agents Chemother. 2017, 61, e00732-00717. [Google Scholar] [CrossRef] [PubMed]
- Tchesnokova, V.; Riddell, K.; Scholes, D.; Johnson, J.; Sokurenko, E. The uropathogenic Escherichia coli subclone ST131-H30 is responsible for most antibiotic prescription errors at an urgent care clinic. Clin. Infect. Dis. 2019, 68, 781–787. [Google Scholar] [CrossRef] [PubMed]
- Price, L.B.; Johnson, J.R.; Aziz, M.; Clabots, C.; Johnston, B.; Tchesnokova, V.; Nordstrom, L.; Billig, M.; Chattopadhyay, S.; Stegger, M.; et al. The epidemic of extended-spectrum-β-lactamase-producing Escherichia coli ST131 is driven by a single highly pathogenic subclone, H30-Rx. mBio 2013, 6, e00377-13. [Google Scholar] [CrossRef]
- Petty, N.K.; Ben Zakour, N.L.; Stanton-Cook, M.; Skippington, E.; Totsika, M.; Forde, B.M.; Phan, M.-D.; Moriel, D.G.; Peters, K.M.; Davies, M.; et al. Global dissemination of a multidrug resistant Escherichia coli clone. Proc. Natl. Acad. Sci. USA 2014, 111, 5694–5699. [Google Scholar] [CrossRef]
- Bonten, M.; Johnson, J.R.; van den Biggelaar, A.; Georgalis, L.; Geurtsen, J.; Ibarra de Palacios, P.; Gravenstein, S.; Verstraeten, T.; Hermans, P.; Poolman, J. Epidemiology of Escherichia coli bacteremia: A systematic literature review. Clin. Infect. Dis. 2021, 72, 1211–1229. [Google Scholar] [CrossRef]
- Johnson, J.R.; Davis, G.; Clabots, C.; Johnston, B.D.; Porter, S.; Debroy, C.; Pomputius, W.; Ender, P.T.; Cooperstock, M.; Slater, B.C.; et al. Household clustering of Escherichia coli sequence type 131 clinical and fecal isolates according to whole genome se-quence analysis. Open Forum Infect. Dis. 2016, 3, ofw129. [Google Scholar] [CrossRef]
- Moreno, E.; Andreu, A.; Pigrau, C.; Kuskowski, M.A.; Johnson, J.R.; Prats, G. Relationship between Escherichia coli strains causing acute cystitis in women and the fecal E. coli population of the host. J. Clin. Microbiol. 2008, 46, 2529–2534. [Google Scholar] [CrossRef]
- Grüneberg, R.N. Relationship of infecting urinary organism to the faecal flora in patients with symptomatic urinary infection. Lancet 1969, 1, 766–768. [Google Scholar] [CrossRef]
- Hooper, L.V.; Littman, D.R.; MacPherson, A.J. Interactions between the microbiota and the immune system. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef]
- Stecher, B. The roles of inflammation, nutrient availability and the commensal microbiota in enteric pathogen infection. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Mendes, V.; Galvão, I.; Vieira, A.T. Mechanisms by Which the gut microbiota influences cytokine production and modulates host inflammatory responses. J. Interferone Cytokine Res. 2019, 39, 393–409. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.; Clabots, C.; Porter, S.B.; Bender, T.; Thuras, P.; Johnson, J.R. Large fecal reservoir of Escherichia coli sequence type 131-H30 subclone strains that are shared within households and re-semble clinical ST131-H30 isolates. J. Infect. Dis. 2019, 221, 1659–1668. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Clabots, C.; Porter, S.B.; Bender, T.; Johnston, B.D.; Thuras, P. Intestinal persistence of colonizing Escherichia coli strains, especially ST131-H30, in relation to bacterial and host factors. J. Infect. Dis. 2022, 225, 2197–2207. [Google Scholar] [CrossRef] [PubMed]
- Berg, D.E.; Akopyants, N.S.; Kersulyte, D. Fingerprinting microbial genomes using the RAPD or AP-PCR method. Methods Mol. Cell. Biol. 1994, 5, 13–24. [Google Scholar]
- Johnson, J.R.; Menard, M.; Johnston, B.; Kuskowski, M.A.; Nichol, K.; Zhanel, G.G. Epidemic clonal groups of Escherichia coli as a cause of antimicrobial-resistant urinary tract infections in Canada, 2002 to 2004. Antimicrob. Agents Chemother. 2009, 53, 2733–2739. [Google Scholar] [CrossRef]
- Johnson, J.R.; Johnston, B.D.; Porter, S.B.; Clabots, C.; Bender, T.L.; Thuras, P.; Trott, D.J.; Cobbold, R.; Mollinger, J.; Ferrieri, P.; et al. Rapid emergence, subsidence, and molecular detection of Escherichia coli sequence type 1193-fimH64, a new dissemi-nated multidrug-resistant commensal and extraintestinal pathogen. J. Clin. Microbiol. 2019, 57, e01664-18. [Google Scholar] [CrossRef]
- Clermont, O.; Christenson, J.K.; Daubie, A.; Gordon, D.M.; Denamur, E. Development of an allele-specific PCR for Escherichia coli B2 sub-typing, a rapid and easy to perform substitute of multilocus sequence typing. J. Microbiol. Methods 2014, 101, 24–27. [Google Scholar] [CrossRef]
- Johnston, B.D.; Thuras, P.; Porter, S.B.; Anacker, M.; VonBank, B.; Vagnone, P.S.; Witwer, M.; Castanheira, M.; Johnson, J.R. Global molecular epidemiology of carbapenem-resistant Escherichia coli (2002–2017). Eur. J. Clin. Microbiol. Infect. Dis. 2021, 1–13. [Google Scholar] [CrossRef]
- Huttner, A.; Hatz, C.; van den Dobbelstein, G.; Abbanat, D.; Hornacek, A.; Frölich, R.; Dreyer, A.M.; Martin, P.; Davies, T.; Fae, K.; et al. immunogenicity, and preliminary clinical efficacy of a vaccine against extraintestinal pathogenic Escherichia coli in women with a history of recurrent urinary tract infection: A randomised, single-blind, placebo-controlled phase 1b trial. Lancet Infect. Dis. 2017, 17, 528–537. [Google Scholar] [CrossRef]
- van den Dobbelsteen, G.P.J.M.; Faé, K.C.; Serroyen, J.; van den Nieuwenhof, I.M.; Braun, M.; Haeuptle, M.A.; Sirena, D.; Schneider, J.; Alaimo, C.; Lipowsky, G.; et al. Immunogenicity and safety of a tetravalent E. coli O-antigen bioconjugate vaccine in animal models. Vaccine 2016, 34, 4152–4160. [Google Scholar] [CrossRef] [PubMed]
- Frenck, R.W.; Ervin, J.; Chu, L.; Abbanat, D.; Spiessens, B.; Go, O.; Haazen, W.; Dobbelsteen, G.V.D.; Poolman, J.; Thoelen, S.; et al. Safety and immunogenicity of a vaccine for extra-intestinal pathogenic Escherichia coli (ESTELLA): A phase 2 random-ised controlled trial. Lancet Infect. Dis. 2019, 19, 631–640. [Google Scholar] [CrossRef]
- Marin, J.; Clermont, O.; Royer, G.; Mercier-Darty, M.; Decousser, J.-W.; Tenaillon, A.; Denamur, E.; Blanquart, F. The popu-lation genomics of increased virulence and antibiotic resistance in human commensal Escherichia coli over 30 years in France. Appl. Environ. Microbiol. 2022, 88, e0066422. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Porter, S.; Thuras, P.; Castanheira, M. The pandemic H30 subclone of sequence type 131 (ST131) as the leading cause of multidrug-resistant Escherichia coli in-fections in the United States (2011–2012). Open Forum Infect. Dis. 2017, 4, ofx089. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.Y.; Cisalpino, D.; Varadarajan, S.; Hellman, J.; Warren, H.S.; Cascalho, M.; Inohara, N.; Núñez, G. Gut microbiota-induced immunoglobulin G controls systemic infection by symbiotic bacteria and pathogens. Immunity 2016, 44, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Vujkovic-Cvijin, I.; Welles, H.C.; Ha, C.W.Y.; Huq, L.; Mistry, S.; Brenchley, J.M.; Trinchieri, G.; Devkota, S.; Belkaid, Y. The systemic anti-microbiota IgG repertoire can identify gut bacteria that translocate across gut barrier surfaces. Sci. Transl. Med. 2022, 14, eabl3927. [Google Scholar] [CrossRef]
- Cevallos, S.A.; Lee, J.-Y.; Tiffany, C.R.; Bundloss, A.J.; Johnston, L.; Byndloss, M.X.; Bäumler, A.J. Increased epithelial oxygenation links colitis to an expansion of tumorigenic bacteria. mBio 2019, 10, e-02244-19. [Google Scholar] [CrossRef]
- Winter, S.E.; Winter, M.G.; Zavier, M.N.; Thiennimitr, P.; Poon, V.; Keestra, A.M.; Laughlin, R.C.; Gomez, G.; Wu, J.; Lawhon, S.D.; et al. Host-derived nitrate boosts growth of E. coli in the inflamed gut. Science 2013, 339, 708–711. [Google Scholar] [CrossRef]
- Becattini, S.; Sorbara, M.T.; Kim, S.G.; Littmann, E.L.; Dong, Q.; Walsh, G.; Wright, R.; Amoretti, L.; Fontana, E.; Hohl, T.M.; et al. Rapid transcriptional and metabolic adaptation of intestinal microbes to host immune activation. Cell Host Microbe 2021, 29, 378–393.e5. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Cevallos, S.A.; Byndloss, M.X.; Tiffany, C.R.; Olsan, E.E.; Butler, B.P.; Young, B.M.; Rogers, A.W.; Nguyen, H.; Kim, K.; et al. High-fat diet and antibiotics cooperatively impair mitochondrial bioenergetics to trigger dysbiosis that exacerbates pre-inflammatory bowel disease. Cell Host Microbe 2020, 28, 273–284.e6. [Google Scholar] [CrossRef]
- Costello, E.K.; Stagaman, K.; Dethlefsen, L.; Bohannan, B.J.M.; Relman, D.A. The application of ecological theory toward an understanding of the human microbiome. Science 2012, 336, 1255–1262. [Google Scholar] [CrossRef]
- Yoo, W.; Zieba, J.K.; Foegeding, N.J.; Torres, T.P.; Shelton, C.D.; Shealy, N.G.; Byndloss, A.J.; Cevallos, S.A.; Gertz, E.; Tiffany, C.R.; et al. High-fat diet–induced colonocyte dysfunction escalates microbiota-derived trimethylamine N-oxide. Science 2021, 373, 813–818. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Casey, J.; Almudevar, A.; Pichichero, M. Correlation of higher antibody levels to pneumococcal proteins with protection from pneumococcal acute otitis media but not protection from nasopharyngeal colonization in young children. Clin. Microbiol. Infect. 2017, 23, 487.e1–487.e6. [Google Scholar] [CrossRef] [PubMed]
- Nurkka, A.; Lahdenkari, M.; Palmu, A.A.; Käyhty, H. The FinOM Study Group Salivary antibodies induced by the seven-valent PncOMPC conjugate vaccine in the Finnish Otitis Media Vaccine Trial. BMC Infect. Dis. 2005, 5, 41. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Subject Characteristics within Subgroup a | ||||||
|---|---|---|---|---|---|---|
| Serum IgG Substudy | Stimulated Cytokine Release Substudy | |||||
| Variable | Total (n = 52) | Control (n = 26) | H30R-Colonized (n = 26) | Total (n = 61) | Control (n = 32) | H30R-Colonized (n = 29) |
| Age, years: median (IQR) | 69 (64,74) | 73 (64,77) | 68 (64,72) | 68 (64,72) | 69 (60.5,77) | 68 (65,72) |
| Male, no. (column percent) | 39 (75) | 19 (73) | 20 (77) | 51 (84) | 28 (88) | 23 (79) |
| Veteran, no. (column percent) | 38 (73) | 18 (69) | 20 (77) | 50 (82) | 27 (84) | 23 (79) |
| ln (EC50 a) or Ratio, Mean (SD b) | ||||
|---|---|---|---|---|
| Total | Control (n = 26) | Colonized (n = 26) | p Value c, Colonized vs. Control | |
| anti-O25b IgG, ln (EC50 a) | 8.8 (1.6) | 8.0 (1.2) | 9.5 (1.6) | <0.001 |
| anti-O6 IgG, ln (EC50 a) | 7.3 (1.3) | 7.3 (1.4) | 7.3 (1.2) | 0.36 |
| anti-O25b/anti-O6 IgG ratio | 1.2 (0.3) | 1.1 (0.2) | 1.3 (0.3) | 0.007 |
| Cytokine Release after Stimulation with JJ1886 (ST131-H30R, O25b:K+:H4) a | Cytokine Release after Stimulation with CFT073 (ST73, O6:K2:H1) a | JJ1886/CFT073 Cytokine Release Ratio a | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Median (IQR b) | Median (IQR b) | Median (IQR b) | |||||||
| Variable | Control | H30R-Colonized | p Value c | Control | H30R- colonized | p Value c | Control | H30R-colonized | p Value c |
| IFNγ (pg/mL) or JJ1886/CFT073 IFNγ ratio | 184 (518) d | 194 (538) e | 0.61 | 426 (743) d | 442 (978) e | 0.58 | 0.50 (0.26) | 0.44 (0.32) | 0.47 |
| TNFα (pg/mL) or JJ1886/CFT073 TNFα ratio | 9353 (5429) f | 7804 (6204) g | 0.48 | 11,451 (2092) f | 12,546 (4606) g | 0.04 | 0.83 (0.39) | 0.53 (0.34) | 0.009 |
| IL-10 (pg/mL) or JJ1886/CFT073 IL-10 ratio | 623 (664) | 408 (323) | 0.06 | 2590 (2252) | 2485 (1332) | 0.87 | 0.26 (0.18) | 0.17 (0.10) | 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnston, B.D.; Clabots, C.; Bender, T.; Porter, S.B.; van den Dobbelsteen, G.; Poolman, J.; Thuras, P.; Johnson, J.R. Colonization with Escherichia coli ST131-H30R (H30R) Corresponds with Increased Serum Anti-O25 IgG Levels and Decreased TNFα and IL-10 Responsiveness to H30R. Pathogens 2023, 12, 603. https://doi.org/10.3390/pathogens12040603
Johnston BD, Clabots C, Bender T, Porter SB, van den Dobbelsteen G, Poolman J, Thuras P, Johnson JR. Colonization with Escherichia coli ST131-H30R (H30R) Corresponds with Increased Serum Anti-O25 IgG Levels and Decreased TNFα and IL-10 Responsiveness to H30R. Pathogens. 2023; 12(4):603. https://doi.org/10.3390/pathogens12040603
Chicago/Turabian StyleJohnston, Brian D., Connie Clabots, Tricia Bender, Stephen B. Porter, Germie van den Dobbelsteen, Jan Poolman, Paul Thuras, and James R. Johnson. 2023. "Colonization with Escherichia coli ST131-H30R (H30R) Corresponds with Increased Serum Anti-O25 IgG Levels and Decreased TNFα and IL-10 Responsiveness to H30R" Pathogens 12, no. 4: 603. https://doi.org/10.3390/pathogens12040603
APA StyleJohnston, B. D., Clabots, C., Bender, T., Porter, S. B., van den Dobbelsteen, G., Poolman, J., Thuras, P., & Johnson, J. R. (2023). Colonization with Escherichia coli ST131-H30R (H30R) Corresponds with Increased Serum Anti-O25 IgG Levels and Decreased TNFα and IL-10 Responsiveness to H30R. Pathogens, 12(4), 603. https://doi.org/10.3390/pathogens12040603