
Snapshot of Phenotypic and Molecular Virulence and Resistance Profiles in Multidrug-Resistant Strains Isolated in a Tertiary Hospital in Romania



 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Evaluation of Antibiotic Susceptibility
2.3. Detection of β-Lactamase Encoding Genes
2.4. Evaluation of the Soluble Enzymatic Factors
2.5. Virulence Genes Detection
2.6. Statistical Analysis
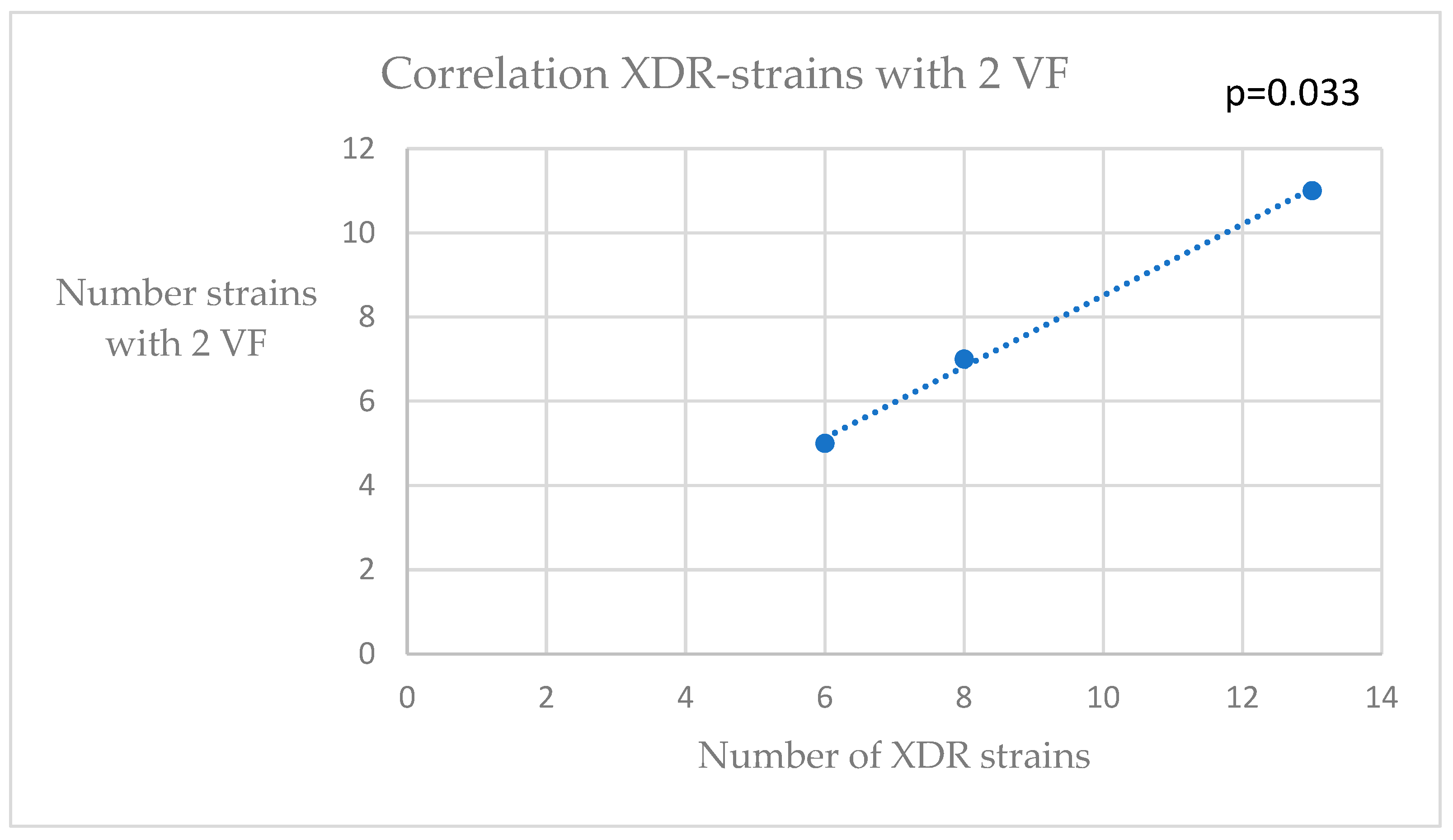
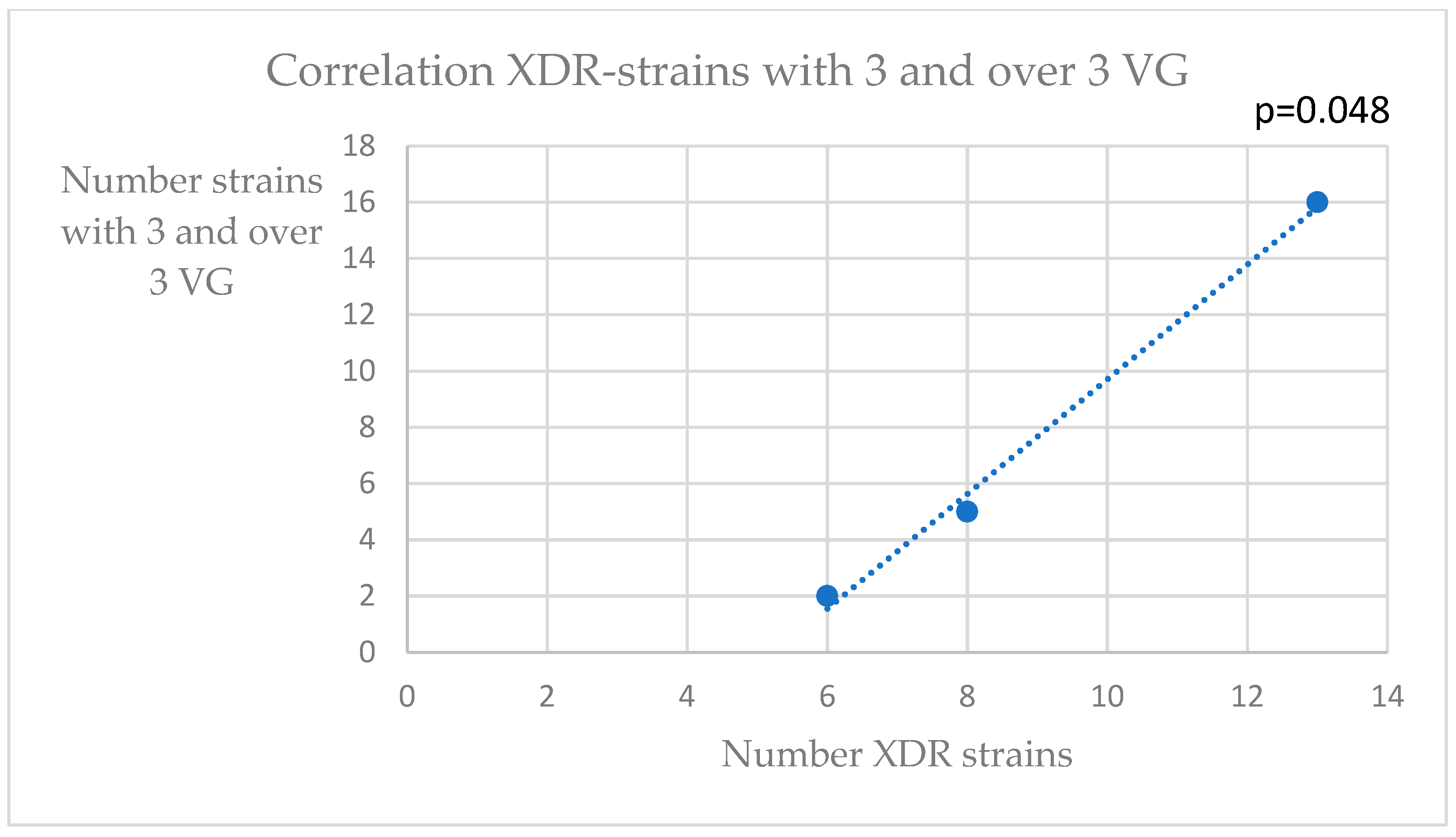
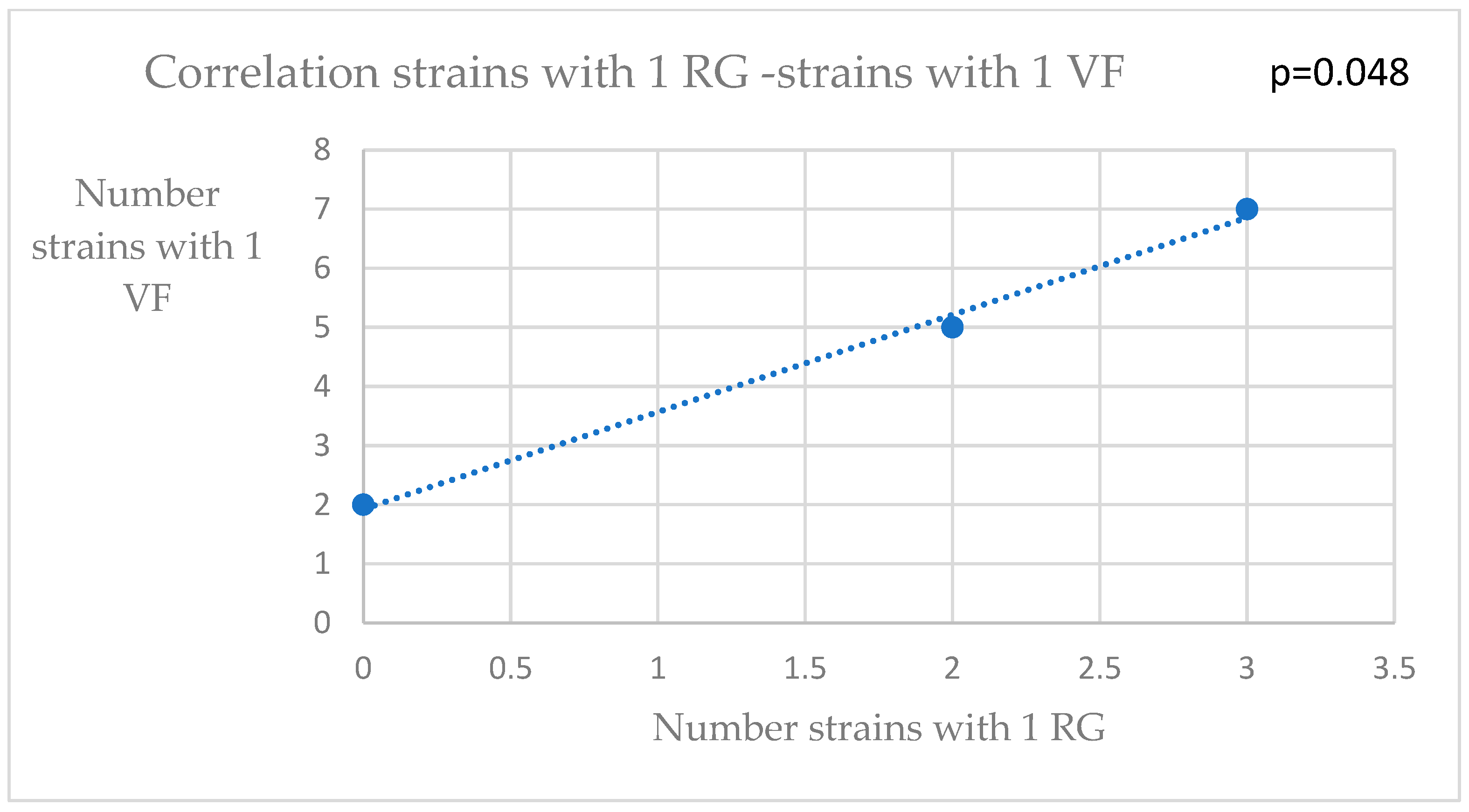
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Madigan, M.T.; Martinko, J.M.; Dunlap, P.V.; Clark, D.P. Brock Biology of Microorganisms, 12th ed.; Pearson Education Inc.: Upper Saddle River, NJ, USA, 2009. [Google Scholar]
- Otto, M. Staphylococcal Biofilms. Microbiol. Spectr. 2018, 6, 1–26. [Google Scholar] [CrossRef]
- Cepas, V.; Soto, S.M. Relationship between Virulence and Resistance among Gram-Negative Bacteria. Antibiotics 2020, 9, 719. [Google Scholar] [CrossRef]
- Beceiro, A.; Tomás, M.; Bou, G. Antimicrobial Resistance and Virulence: A Successful or Deleterious Association in the Bacterial World? Clin. Microbiol. Rev. 2013, 26, 185–230. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.B. The Antibiotic Paradox. How Miracle Drugs Are Destroying the Miracle; Springer: New York, NY, USA, 1992; ISBN 978-1-4899-6042-9. Available online: https://link.springer.com/book/10.1007/978-1-4899-6042-9 (accessed on 15 February 2023).
- Wang, G.; Zhao, G.; Chao, X.; Xie, L.; Wang, H. The Characteristic of Virulence, Biofilm and Antibiotic Resistance of Klebsiella pneumoniae. Int. J. Environ. Res. Public Health 2020, 17, 6278. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.G.; Rosa, A.C.; Cardoso, O. Virulence factors as predictive tools for drug resistance in Pseudomonas aeruginosa. Virulence 2015, 6, 679–683. [Google Scholar] [CrossRef]
- Viveiros, M.; Dupont, M.; Rodrigues, L.; Couto, I.; Davin-Regli, A.; Martins, M.; Pagès, J.M.; Amaral, L. Antibiotic stress, genetic response and altered permeability of E. coli. PLoS ONE 2007, 2, e365. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Q.; Huang, H.; Huang, W.; Chen, Y.; McGarvey, P.B.; Wu, C.H.; Arighi, C.N. UniProt Consortium. A crowdsourcing open platform for literature curation in UniProt. PLoS Biol. 2021, 19, e3001464. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Huang, W.E.; Yang, Q. Clinical Perspective of Antimicrobial Resistance in Bacteria. Infect. Drug Resist. 2022, 15, 735–746. [Google Scholar] [CrossRef]
- Halat, D.H.; Moubareck, C.A. The Current Burden of Carbapenemases: Review of Significant Properties and Dissemination among Gram-Negative Bacteria. Antibiotics 2020, 9, 186. [Google Scholar] [CrossRef]
- Santajit, S.; Indrawattana, N. Mechanisms of antimicrobial resistance in ESKAPE pathogens. Biomed Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef]
- Ambler, R.P. The structure of beta-lactamases. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1980, 289, 321–331. [Google Scholar] [CrossRef]
- Liao, C.; Huang, X.; Wang, Q.; Yao, D.; Lu, W. Virulence Factors of Pseudomonas aeruginosa and Antivirulence Strategies to Combat Its Drug Resistance. Front. Cell. Infect. Microbiol. 2022, 12, 926758. [Google Scholar] [CrossRef] [PubMed]
- Baquero, M.R.; Galán, J.C.; Turrientes, M.C.; Cantón, R.; Coque, M.T.; Martínez, J.L.; Baquero, F. Increased Mutation Frequencies in Escherichia coli Isolates Harboring Extended-Spectrum β-Lactamases. Antimicrob. Agents Chemother. 2005, 49, 4754–4756. [Google Scholar] [CrossRef] [PubMed]
- Lazar, V.; Chifiriuc, M.C. Medical significance and new therapeutical strategies for biofilm associated infections. Roum. Arch. Microbiol. Immunol. 2010, 69, 125–138. [Google Scholar] [PubMed]
- Simanek, K.A.; Paczkowski, J.E. Resistance Is Not Futile: The Role of Quorum Sensing Plasticity in Pseudomonas aeruginosa Infections and Its Link to Intrinsic Mechanisms of Antibiotic Resistance. Microorganisms 2022, 10, 1247. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.M.; Bachman, M.A. Colonization, Infection, and the Accessory Genome of Klebsiella pneumoniae. Front. Cell. Infect. Microbiol. 2018, 8, 4. [Google Scholar] [CrossRef]
- Du, F.-L.; Huang, Q.-S.; Wei, D.-D.; Mei, Y.-F.; Long, D.; Liao, W.-J.; Wan, L.-G.; Liu, Y.; Zhang, W. Prevalence of Carbapenem-Resistant Klebsiella pneumoniae Co-Harboring BlaKPC-Carrying Plasmid and PLVPK-Like Virulence Plasmid in Bloodstream Infections. Front. Cell. Infect. Microbiol. 2020, 10, 556654. [Google Scholar] [CrossRef]
- Zhong, S.; He, S. Quorum Sensing Inhibition or Quenching in Acinetobacter baumannii: The Novel Therapeutic Strategies for New Drug Development. Front. Microbiol. 2021, 12, 558003. [Google Scholar] [CrossRef]
- Kyriakidis, I.; Vasileiou, E.; Pana, Z.D.; Tragiannidis, A. Acinetobacter baumannii Antibiotic Resistance Mechanisms. Pathogens 2021, 10, 373. [Google Scholar] [CrossRef]
- Delcaru, C.; Alexandru, I.; Podgoreanu, P.; Grosu, M.; Stavropoulos, E.; Chifiriuc, M.C.; Lazar, V. Microbial Biofilms in Urinary Tract Infections and Prostatitis: Etiology, Pathogenicity, and Combating strategies. Pathogens 2017, 5, 65. [Google Scholar] [CrossRef]
- Lazar, V.; Holban, A.M.; Curutiu, C.; Chifiriuc, M.C. Modulation of Quorum Sensing and Biofilms in Less Investigated Gram-Negative ESKAPE Pathogens. Front. Microbiol. 2021, 12, 676510. [Google Scholar] [CrossRef] [PubMed]
- Trușcă, B.S.; Ianculescu, E.; Maruțescu, L.; Gheorghe-Barbu, I.; Chifiriuc, M.C.; Lazăr, V. Comparative performance of two immunochromatographic tests for the rapid detection of PCR confirmed, carbapenemase producing-Enterobacterales. Biointerface Res. Apll Chem. 2022, 13, 322. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 31st ed.; CLSI supplement M100 ([Electronic]); Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2021; ISBN 978-1-68440-105-5. [Google Scholar]
- Georgescu, M.; Gheorghe, I.; Curutiu, C.; Lazar, V.; Bleotu, C.; Chifiriuc, M.C. Virulence and resistance features of Pseudomonas aeruginosa strains isolated from chronic leg ulcers. BMC Infect. Dis. 2016, 16 (Suppl. 1), 92. [Google Scholar] [CrossRef] [PubMed]
- Available online: http://vassarstats.net/ (accessed on 5 September 2022).
- Available online: http://gnu.org/software/pspp/ (accessed on 2 March 2023).
- Borcan, E.; Ghiţă, C.; Chifiriuc, M.C.; Lazăr, V. Antibiotic resistance profiles of Acinetobacter sp. strains isolated from intensive-care unit patients. Roum. Arch. Microbiol. Immunol. 2012, 71, 75–80. [Google Scholar]
- Gheorghe, I.; Novais, A.; Grosso, F.; Rodrigues, C.; Chifiriuc, M.C.; Lazar, V.; Peixe, L. Snapshop of carbapenemase-producing Pseudomonas aeruginosa and Acinetobacter baumannii in Bucharest hospitals reveals unusual clones and novel genetic surroundings for blaOXA-23. J. Antimicrob. Chemother. 2015, 70, 1016–1020. [Google Scholar] [CrossRef] [PubMed]
- Gheorghe, I.; Cristea, V.C.; Marutescu, L.; Popa, M.; Murariu, C.; Trusca, B.S.; Borcan, E.; Ghita, M.C.; Lazar, V.; Chifiriuc, M.C. Resistance and Virulence Features in Carbapenem-resistant Acinetobacter baumannii Community Acquired and Nosocomial Isolates in Romania. Rev. Chim. 2019, 70, 3502–3507. [Google Scholar] [CrossRef]
- Boyd, A.; Chakrabarty, A.M. Pseudomonas aeruginosa biofilms: Role of the alginate exopolysaccharide. J. Ind. Microbiol. 1995, 15, 162–168. [Google Scholar] [CrossRef]
- Santos, D.A.; Nascimento, M.M.; Vitali, L.H.; Martinez, R. In vitro activity of antimicrobial combinations against multidrug-resistant Pseudomonas aeruginosa. Rev. Soc. Bras. Med. Trop. 2013, 46, 299–303. [Google Scholar] [CrossRef]
- Shahid, M.; Malik, A.; Sheeba. Multidrug-resistant Pseudomonas aeruginosa strains harbouring R-plasmids and AmpC β-lactamases isolated from hospitalised burn patients in a tertiary care hospital of North India. FEMS Microbiol. Lett. 2003, 228, 181–186. [Google Scholar] [CrossRef]
- Rocha, A.J.; Barsottini, M.R.O.; Rocha, R.R.; Laurindo, M.V.; Moraes, F.L.L.; Rocha, S.L. Pseudomonas aeruginosa: Virulence Factors and Antibiotic Resistance Genes. Braz. Arch. Biol. Technol. 2019, 62, e19180503. [Google Scholar] [CrossRef]
- Liao, W.; Huang, N.; Zhang, Y.; Sun, Y.; Chen, T.; Zeng, W.; Chen, L.; Wen, H.; Cao, J.; Zhou, T. Comparison of Carbapenem-Resistant Klebsiella Pneumoniae Strains Causing Intestinal Colonization and Extraintestinal Infections: Clinical, Virulence, and Molecular Epidemiological Characteristics. Front. Public Health 2021, 9, 783124. [Google Scholar] [CrossRef]
- Lazăr, V.; Măruțescu, L.G.; Chifiriuc, M.C. Microbiologie Generala și Aplicată; Editura Universității din București: Bucharest, Romania, 2017. [Google Scholar]
- Toder, D.S.; Ferrell, S.J.; Nezezon, J.L.; Rust, L.; Iglewski, B.H. lasA and lasB genes of Pseudomonas aeruginosa: Analysis of transcription and gene product activity. Infect. Immun. 1994, 62, 1320–1327. [Google Scholar] [CrossRef] [PubMed]
- Galdino, A.; Viganor, L.; de Castro, A.A.; da Cunha, E.; Mello, T.P.; Mattos, L.M.; Pereira, M.D.; Hunt, M.C.; O’Shaughnessy, M.; Howe, O.; et al. Disarming Pseudomonas aeruginosa Virulence by the Inhibitory Action of 1,10-Phenanthroline-5,6-Dione-Based Compounds: Elastase B (LasB) as a Chemotherapeutic Target. Front. Microbiol. 2019, 10, 1701. [Google Scholar] [CrossRef]
- Gupta, V. Metallo beta lactamases in Pseudomonas aeruginosa and Acinetobacter species. Expert Opin. Investig. Drugs 2008, 17, 131–143. [Google Scholar] [CrossRef]
- Walsh, T.R.; Toleman, M.A.; Poirel, L.; Nordmann, P. Metallo-beta-lactamases: The quiet before the storm? Clin. Microbiol. Rev. 2005, 18, 306–325. [Google Scholar] [CrossRef] [PubMed]
- Faraji, F.; Mahzounieh, M.; Ebrahimi, A.; Fallah, F.; Teymournejad, O.; Lajevardi, B. Molecular detection of virulence genes in Pseudomonas aeruginosa isolated from children with Cystic Fibrosis and burn wounds in Iran. Microb. Pathog. 2016, 99, 1–4. [Google Scholar] [CrossRef]
- Qin, S.; Xiao, W.; Zhou, C.; Pu, Q.; Deng, X.; Lan, L.; Liang, H.; Song, X.; Wu, M. Pseudomonas aeruginosa: Pathogenesis, virulence factors, antibiotic resistance, interaction with host, technology advances and emerging therapeutics. Signal Transduct. Target. Ther. 2022, 7, 199. [Google Scholar] [CrossRef] [PubMed]
- Jurado-Martín, I.; Sainz-Mejías, M.; McClean, S. Pseudomonas aeruginosa: An Audacious Pathogen with an Adaptable Arsenal of Virulence Factors. Int. J. Mol. Sci. 2021, 22, 3128. [Google Scholar] [CrossRef]
- Hsieh, P.-F.; Lu, Y.-R.; Lin, T.-L.; Lai, L.-Y.; Wang, J.-T. Klebsiella pneumoniae Type VI Secretion System Contributes to Bacterial Competition, Cell Invasion, Type-1 Fimbriae Expression, and In Vivo Colonization. J. Infect. Dis. 2019, 219, 637–647. [Google Scholar] [CrossRef]
- Tang, M.; Kong, X.; Hao, J.; Liu, J. Epidemiological Characteristics and Formation Mechanisms of Multidrug-Resistant Hypervirulent Klebsiella pneumoniae. Front. Microbiol. 2020, 11, 581543. [Google Scholar] [CrossRef] [PubMed]
- Huynh, D.T.N.; Kim, A.Y.; Kim, Y.R. Identification of Pathogenic Factors in Klebsiella pneumoniae Using Impedimetric Sensor Equipped with Biomimetic Surfaces. Sensors 2017, 17, 1406. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Wang, T.; Chen, L.; Du, H. Virulence Factors in Hypervirulent Klebsiella pneumoniae. Front. Microbiol. 2021, 12, 642484. [Google Scholar] [CrossRef] [PubMed]
- Turton, J.F.; Perry, C.; Elgohari, S.; Hampton, C.V. PCR characterization and typing of Klebsiella pneumoniae using capsular type-specific, variable number tandem repeat and virulence gene targets. J. Med. Microbiol. 2010, 59, 541–547. [Google Scholar] [CrossRef]
- Lee, C.R.; Lee, J.H.; Park, K.S.; Jeon, J.H.; Kim, Y.B.; Cha, C.J.; Jeong, B.C.; Lee, S.H. Antimicrobial Resistance of Hypervirulent Klebsiella pneumoniae: Epidemiology, Hypervirulence-Associated Determinants, and Resistance Mechanisms. Front. Cell. Infect. Microbiol. 2017, 7, 483. [Google Scholar] [CrossRef]
- Dong, N.; Yang, X.; Chan, E.W.; Zhang, R.; Chen, S. Klebsiella species: Taxonomy, hypervirulence and multidrug resistance. EBioMedicine 2022, 79, 103998. [Google Scholar] [CrossRef] [PubMed]
- Ballén, V.; Gabasa, Y.; Ratia, C.; Ortega, R.; Tejero, M.; Soto, S. Antibiotic Resistance and Virulence Profiles of Klebsiella pneumoniae Strains Isolated From Different Clinical Sources. Front. Cell. Infect. Microbiol. 2021, 11, 738223. [Google Scholar] [CrossRef]
- Alcántar-Curiel, M.D.; Blackburn, D.; Saldaña, Z.; Gayosso-Vázquez, C.; Iovine, N.; De la Cruz, M.A.; Girón, J.A. Multi-Functional Analysis of Klebsiella pneumoniae Fimbrial Types in Adherence and Biofilm Formation. Virulence 2013, 4, 129–138. [Google Scholar] [CrossRef]
- Dargahi, Z.; Hamad, A.A.; Sheikh, A.F.; Ahmad Khosravi, N.; Samei Fard, S.; Motahar, M.; Mehr, F.J.; Abbasi, F.; Meghdadi, H.; Bakhtiyariniya, P.; et al. The Biofilm Formation and Antibiotic Resistance of Bacterial Profile from Endotracheal Tube of Patients Admitted to Intensive Care Unit in Southwest of Iran. PLoS ONE 2022, 17, e0277329. [Google Scholar] [CrossRef]
- Ciofu, O.; Tolker-Nielsen, T. Tolerance and Resistance of Pseudomonas aeruginosa Biofilms to Antimicrobial Agents-How P. aeruginosa Can Escape Antibiotics. Front. Microbiol. 2019, 10, 913. [Google Scholar] [CrossRef]
- Hazhirkamal, M.; Zarei, O.; Movahedi, M.; Karami, P.; Shokoohizadeh, L.; Taheri, M. Molecular Typing, Biofilm Production, and Detection of Carbapenemase Genes in Multidrug-Resistant Acinetobacter baumannii Isolated from Different Infection Sites Using ERIC-PCR in Hamadan, West of Iran. BMC Pharm. Toxicol. 2021, 22, 32. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, S.T.; Bassler, B.L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.; Al-Saryi, N.; Al-Kadmy, I.M.S.; Aziz, S.N. Multidrug-resistant Acinetobacter baumannii as an emerging concern in hospitals. Mol. Biol. Rep. 2021, 48, 6987–6998. [Google Scholar] [CrossRef]
- Zeighami, H.; Valadkhani, F.; Shapouri, R.; Samadi, E.; Haghi, F. Virulence characteristics of multidrug resistant biofilm forming Acinetobacter baumannii isolated from intensive care unit patients. BMC Infect. Dis. 2019, 19, 629. [Google Scholar] [CrossRef] [PubMed]
- Lazar, V. Quorum sensing in biofilms--how to destroy the bacterial citadels or their cohesion/power? Anaerobe 2011, 17, 280–285. [Google Scholar] [CrossRef]
- Saipriya, K.; Swathi, C.H.; Ratnakar, K.S.; Sritharan, V. Quorum-sensing system in Acinetobacter baumannii: A potential target for new drug development. J. Appl. Microbiol. 2020, 128, 15–27. [Google Scholar] [CrossRef]
- Holban, A.M.; Chifiriuc, M.C.; Cotar, A.I.; Bleotu, C.; Grumezescu, A.M.; Banu, O.; Lazar, V. Virulence markers in Pseudomonas aeruginosa isolates from hospital acquired infections occurred in patients with underlying cardiovascular disease. Rom. Biotechnol. Lett. 2013, 18, 8843–8854. [Google Scholar]
- Főldes, A.; Bilca, D.; Székely, E. Phenotypic and molecular identification of carbapenemase-producing Enterobacteriaceae-challenges in diagnosis and treatment. Rev. Romana Med. Lab. 2018, 26, 221–230. [Google Scholar] [CrossRef]
- Gavriliu, L.C.; Benea, O.E.; Benea, S. Antimicrobial resistance temporal trend of Klebsiella pneumoniae isolated from blood. J. Med. Life 2016, 9, 419–423. [Google Scholar] [CrossRef]
- Gheorghe, I.; Czobor, I.; Chifiriuc, M.C.; Borcan, E.; Ghita, C.; Banu, O.; Lazar, V.; Mihaescu, G.; Mihailescu, D.; Zhiyong, Z. Molecular screening of carbapenemase-producing Gram-negative strains in Romanian intensive care units during a one year survey. J. Med. Microbiol. 2014, 63, 1303–1310. [Google Scholar] [CrossRef]
- Lixandru, B.E.; Cotar, A.I.; Straut, M.; Usein, C.R.; Cristea, D.; Ciontea, S. Carbapenemase-Producing Klebsiella pneumoniae in Romania: A Six-Month Survey. PLoS ONE 2015, 10, e0143214. [Google Scholar] [CrossRef] [PubMed]
- Georgescu, M.; Gheorghe, I.; Dudu, A.; Czobor, I.; Costache, M.; Cristea, V.C.; Lazar, V.; Chifiriuc, M.C. First report of OXA-72 producing Acinetobacter baumannii in Romania. New Microbes New Infect. 2016, 13, 87–88. [Google Scholar] [CrossRef] [PubMed]
- Perlman, R.L. Life Histories of Pathogen Populations. Int. J. Infect. Dis. 2009, 13, 121–124. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Name | Primer Sequence | Amplicon Size (bp) |
|---|---|---|---|
| Primers used for antibiotic resistance genes detection | |||
| blaIMP | IMP-F IMP-R | 5′-GGAATAGAGTGGCTTAAYTCTC-3′ 5′-GGTTTAAYAAAACAACCACC-3′ | 232 |
| blaVIM | VIM-F VIM-R | 5′-GATGGTGTTTGGTCGCATA-3′ 5′-CGAATGCGCAGCACCAG-3′ | 390 |
| blaOXA-48 | OXA-48-F OXA-48-R | 5′-ATGCGTGTATTAGCCTTATC-3′ 5′-CTAGGGAATAATTTTTTCCT-3′ | 438 |
| blaNDM | NDM-F NDM-R | 5′-GGGCAGTCGCTTCCAACGGT-3′ 5′-GTAGTGCTCAGTGTCGGCAT-3′ | 621 |
| blaKPC | KPC-F KPC-R | 5′-GCTCAGGCGCAACTGTAA-3′ 5′-AGCACAGCGGCAGCAAGAAAG-3′ | 798 |
| blaCTX-M | CTX-M-F CTX-M-R | 5′-CGCTGTTGTTAGGAAGTGTG-3′ 5′-GGCTGGGTGAAGTAAGTGAC-3′ | 754 |
| blaTEM | TEM-F TEM-R | 5′-ATGAGTATTCAACATTTCCGT-3′ 5′-TTACCAATGCTTAATCAGTGA-3′ | 1080 |
| blaOXA-23 | OXA-23-F OXA-23-R | 5′-ATGAGTTATCTATTTTTGTC-3′ 5′-TGTCAAGCTCTTAAATAATA-3′ | 501 |
| blaOXA-24 | OXA24/40-F OXA24/40-R | 5′ GCAGAAAGAAGTAAARCGGGT3′ 5′ CCAACCWGTCAACCAACCTA3′ | 270 |
| blaOXA-51 | OXA-51-F OXA-51-R | 5′-TAATGCTTTGATCGGCCTTG-3′ 5′-TGGATTGCACTTCATCTTGG-3′ | 353 |
| Primers used for virulence markers detection | |||
| epsA | epsA-F epsA-R | 5′-AGCAAGTGGTTATCCAATCG-3′ 5′-T ACCAGACTCACCCATTACA-3′ | 451 |
| ompA | ompA-F ompA-R | 5′-CGCTTCTGCTGGTGCTGAAT-3′ 5′-CGTGCAGTAGCGTTAGGGTA-3′ | 531 |
| plcH | plcH-F plcH-R | 5′GAAGCCATGGGCTACTTCAA-3′ 5′AGAGTGACGAGGGGTAG-3′ | 466 |
| plcN | plcN-F plcN-R | 5′-GTTATCGCAACCAGCCCTAC-3′ 5′-AGGTCGAACACCTGGAACAC-3′ | 307 |
| exoU | exoU-F exoU-R | 5′-CCGTTGTGGTGCCGTTGAAG-3′ 5′-CCAGATGTTCACCGACTCG-3′ | 134 |
| exoT | ExoT-F ExoT-R | 5′-AATCGCCGTCCAACTGCATGCG-3′ 5′-TGTTCGCCGAGGTACTGCTC-3′ | 152 |
| exoS | ExoS-F ExoS-R | 5′-ATCGCTTCAGCAGAGTCCGTC-3′ 5′-CAGGCCAGATCAAGGCCGCGC-3′ | 1352 |
| algD | algD-F algD-R | 5′-ATGCGAATCAGCATCTTTGGT-3′ 5′-CTACCAGCAGATGCCCTCGGC-3′ | 1310 |
| TC | TCF TCR | 5′-TATTTCGCCGACTCCCTGTA-3′ 5′-GAATAGACGCCGCTGAAATC3′ | 752 |
| fimH | fimH-F fimH-R | 5′-TGCAGAACGGATAAGCCGTGG3′ 5′-GCAGTCACCTGCCCTCCGGTA3′ | 506 |
| fimA | fimA-F fimA-R | 5′-CGGACGGTACGCTGTATTTT-3′ 5′-GCTTCGGCGTTGTCTTTATC-3′ | 500 |
| hlyD | hlyD-F hlyD-R | 5′-CTCCGGTACGTGAAAAGGAC3′ 5′-GCCCTGATTACTGAAGCCTG3′ | 904 |
| hlyA | hlyA-F hlyA-R | 5′-AACAAGGATAAGCACTGTTCTGGCT-3′ 5′-ACCATATAAGCGGTCATTCCCGTCA-3′ | 1177 |
| cnf1 | cnf1-F cnf1-R | 5′-GAACTTATTAAGGATAGT-3′ 5′-CATTATTTATAACGCTG-3′ | 544 |
| ecpRAB | ecpRAB-F ecpRAB-R | 5′-CCTATGTAATTAATGGCAGGTTT-3′ 5′-GCTGTTCATAAAGGATGAAATATC-3′ | 1025 |
| mrkA | mrkA-F mrkA-R | 5′-CGGTAAAGTTACCGACGTATCTTGTACTG-3′ 5′-GCTGTTAACCACACCGGTGGTAAC-3′ | 498 |
| mrkD | mrkD-F mrkD-R | 5′-CTGACGCTTTTTATTGGCTTAATGGCGC-3′ 5-′GCAGAATTTCCGGTCTTTTCGTTTAGTAG-3′ | 756 |
| Isolation Source | Number (n) of Isolated Species | ||
|---|---|---|---|
| K. pneumoniae | Acinetobacter spp. | P. aeruginosa | |
| Urine | n = 28 | n = 3 | n = 4 |
| Blood cultures | n = 10 | n = 3 | n = 4 |
| Others * | n = 13 | n = 5 | n = 2 |
| Microorganism | Virulence Factors | Urine N (%) * | Blood Culture N (%) * | Bile Culture N (%) * | Wound N (%) * | Bronchial Aspirate N (%) * | Rectal Swab N (%) * | Drainage Tube N (%) * | Ascitic Fluid N (%) * | Stool Culture N (%) * | p-Value ** |
|---|---|---|---|---|---|---|---|---|---|---|---|
| K. pneumoniae | amylase | 9 (34.6) | 1 (10) | 0 | 0 | 0 | 1 (50) | 1 (50) | 1 (100) | 0 | 0.349 |
| esculinase | 25 (96.2) | 10 (100) | 1 (100) | 1 (100) | 4 (100) | 2 (100) | 2 (100) | 1 (100) | 2 (66.7) | 0.556 | |
| lecithinase | 2 (7.7) | 2 (20) | 0 | 0 | 0 | 0 | 0 | 0 | 2 (66.7) | 0.251 | |
| caseinase | 0 | 2 (20) | 0 | 0 | 1 (25) | 0 | 1 (50) | 0 | 1 (33.3) | 0.047 | |
| lipase | 9 (34.6) | 3 (30) | 0 | 0 | 1 (25) | 0 | 1 (50) | 0 | 2 (66.7) | 0.833 | |
| gelatinase | 1 (3.8) | 3 (30) | 0 | 0 | 0 | 0 | 0 | 1 (100) | 0 | 0.059 | |
| hemolysin | 2 (7.7) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.991 | |
| P. aeruginosa | protease | 2 (50) | 2 (50) | 0 | 1 (50) | 0 | 0 | 0 | 0 | 0 | NA |
| amylase | 0 | 0 | 0 | 1 (50) | 0 | 0 | 0 | 0 | 0 | 0.108 | |
| esculinase | 0 | 1 (25) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.435 | |
| lecithinase | 3 (75) | 4 (100) | 0 | 2 (100) | 0 | 0 | 0 | 0 | 0 | 0.435 | |
| caseinase | 1 (25) | 2 (50) | 0 | 1 (50) | 0 | 0 | 0 | 0 | 0 | 0.732 | |
| lipase | 0 | 2 (50) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.153 | |
| hemolysin | 4 (100) | 4 (100) | 0 | 2 (100) | 0 | 1 (100) | 0 | 0 | 0 | NA | |
| Acinetobacter spp. | amylase | 0 | 0 | 0 | 0 | 0 | 0 | 1 (100) | 0 | 0 | 0.088 |
| lipase | 0 | 1 (33.3) | 0 | 0 | 1 (100) | 1 (100) | 0 | 0 | 0 | 0.266 |
| Microorganism | Virulence Gene Pattern | Urine N (%) * | Blood Culture N (%) * | Other N (%) * | p-Value ** |
|---|---|---|---|---|---|
| K. pneumoniae | mrkA | 0 | 1 (10) | 2 (13.3) | 0.11 |
| fimA | 0 | 1 (10) | 0 | 0.19 | |
| fimA, mrkA | 0 | 2 (20) | 0 | 0.03 | |
| fimA, ecpRAB | 0 | 1 (10) | 0 | 0.19 | |
| ecpRAB, mrkA | 8 (30) | 2 (20) | 8 (53.3) | 0.22 | |
| fimA, ecpRAB, mrkA | 10 (38) | 2 (20) | 4 (26.6) | 0.56 | |
| fimH, ecpRAB, cnf1 | 1 (3.8) | 0 | 0 | 0.99 | |
| fimH, mrkA, cnf1 | 1 (3.8) | 0 | 0 | 0.99 | |
| fimH, fimA, ecpRAB, mrkA | 1 (3.8) | 0 | 0 | 0.99 | |
| fimA, hlyD. hlyA, ecpRAB | 1 (3.8) | 0 | 0 | 0.99 | |
| hlyD, hlyA, ecpRAB, mrkA | 0 | 1 (10) | 1 (6.66) | 0.23 | |
| fimH, hlyD, hlyA, ecpRAB, cnf1 | 1 (3.8) | 0 | 0 | 0.99 | |
| fimH, fimA, ecpRAB, mrkA, cnf1 | 1 (3.8) | 0 | 0 | 0.99 | |
| P. aeruginosa | plcH, plcN, exoU, algD | 0 | 1 (33.3) | 0 | 0.9 |
| plcH, plcN, exoU, exoS, algD | 2 (50) | 1 (33.3) | 0 | 0.9 | |
| plcH, plcN, exoT, exoS, algD | 1 (25) | 1 (33.3) | 1 (50) | 1 | |
| plcH, plcN, exoU, exoT, exoS, algD | 0 | 1 (33.3) | 0 | 0.9 | |
| Acinetobacter spp. | ompA | 3 (100) | 3 (100) | 4 (80) | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truşcă, B.S.; Gheorghe-Barbu, I.; Manea, M.; Ianculescu, E.; Barbu, I.C.; Măruțescu, L.G.; Dițu, L.-M.; Chifiriuc, M.-C.; Lazăr, V. Snapshot of Phenotypic and Molecular Virulence and Resistance Profiles in Multidrug-Resistant Strains Isolated in a Tertiary Hospital in Romania. Pathogens 2023, 12, 609. https://doi.org/10.3390/pathogens12040609
Truşcă BS, Gheorghe-Barbu I, Manea M, Ianculescu E, Barbu IC, Măruțescu LG, Dițu L-M, Chifiriuc M-C, Lazăr V. Snapshot of Phenotypic and Molecular Virulence and Resistance Profiles in Multidrug-Resistant Strains Isolated in a Tertiary Hospital in Romania. Pathogens. 2023; 12(4):609. https://doi.org/10.3390/pathogens12040609
Chicago/Turabian StyleTruşcă, Bianca Simona, Irina Gheorghe-Barbu, Marina Manea, Elvira Ianculescu, Ilda Czobor Barbu, Luminița Gabriela Măruțescu, Lia-Mara Dițu, Mariana-Carmen Chifiriuc, and Veronica Lazăr. 2023. "Snapshot of Phenotypic and Molecular Virulence and Resistance Profiles in Multidrug-Resistant Strains Isolated in a Tertiary Hospital in Romania" Pathogens 12, no. 4: 609. https://doi.org/10.3390/pathogens12040609
APA StyleTruşcă, B. S., Gheorghe-Barbu, I., Manea, M., Ianculescu, E., Barbu, I. C., Măruțescu, L. G., Dițu, L. -M., Chifiriuc, M. -C., & Lazăr, V. (2023). Snapshot of Phenotypic and Molecular Virulence and Resistance Profiles in Multidrug-Resistant Strains Isolated in a Tertiary Hospital in Romania. Pathogens, 12(4), 609. https://doi.org/10.3390/pathogens12040609